
The Influencing Factors of Aggregation Behavior of Tree-of-Heaven Trunk Weevil, Eucryptorrhynchus brandti (Harold) (Coleoptera: Curculionidae)



Abstract
:Simple Summary
Abstract
1. Introduction
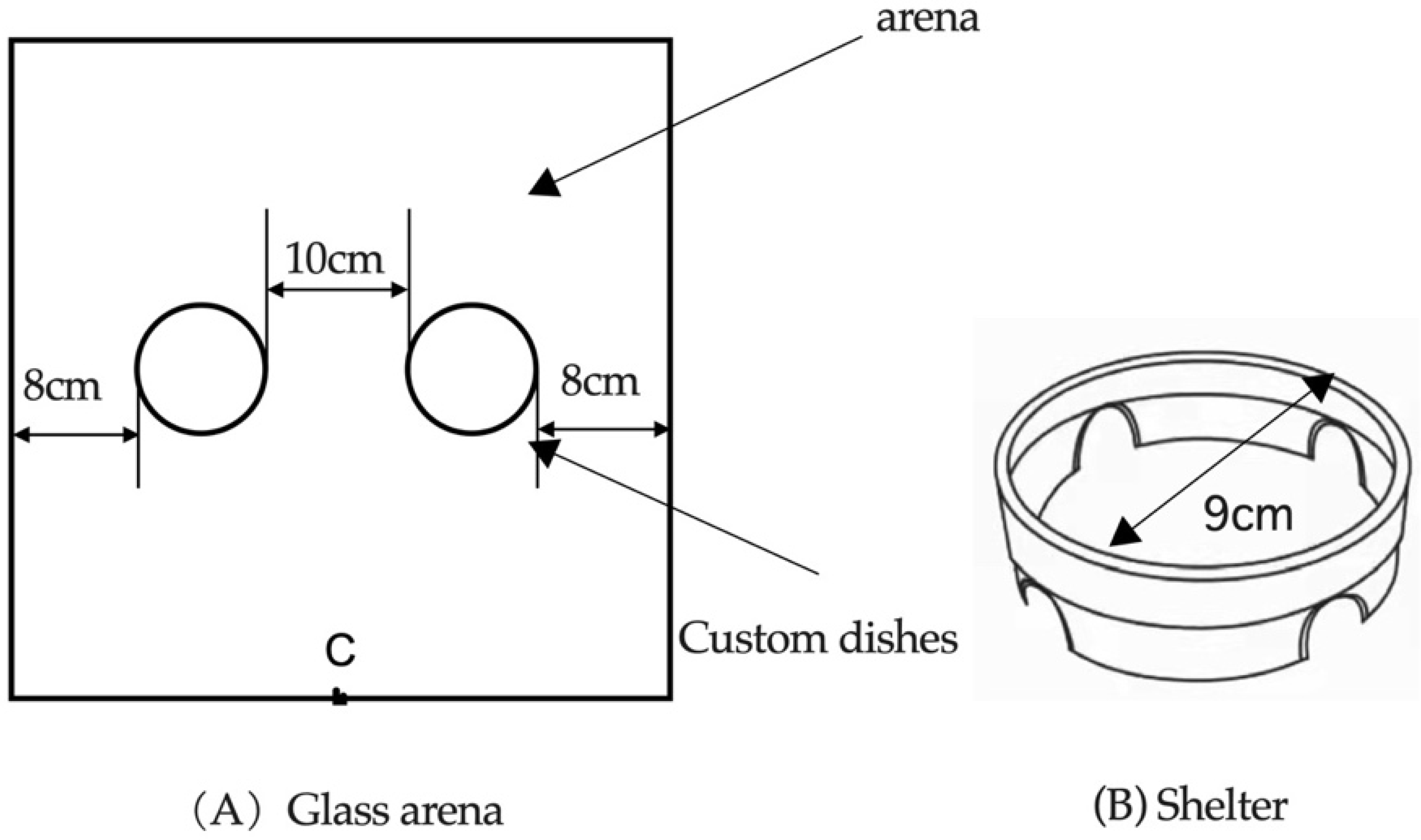
2. Materials and Methods
2.1. Test Insects
2.2. Binomial Choice Experiment of Host Plant
2.3. Temperature Experiment
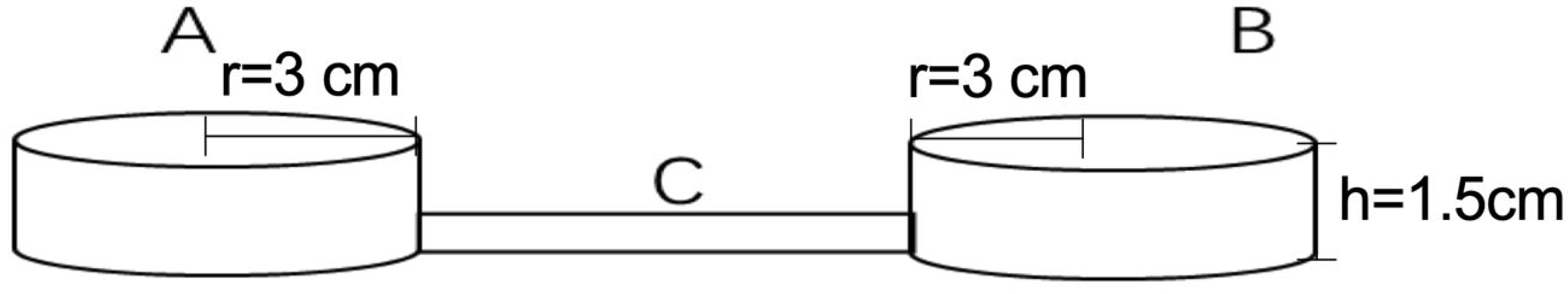
2.4. Conspecific Individual Interaction
2.4.1. Gender Factor
2.4.2. Reaction to Crude Extracts
2.5. Light–Dark Experiments
2.6. Data Analysis
3. Results
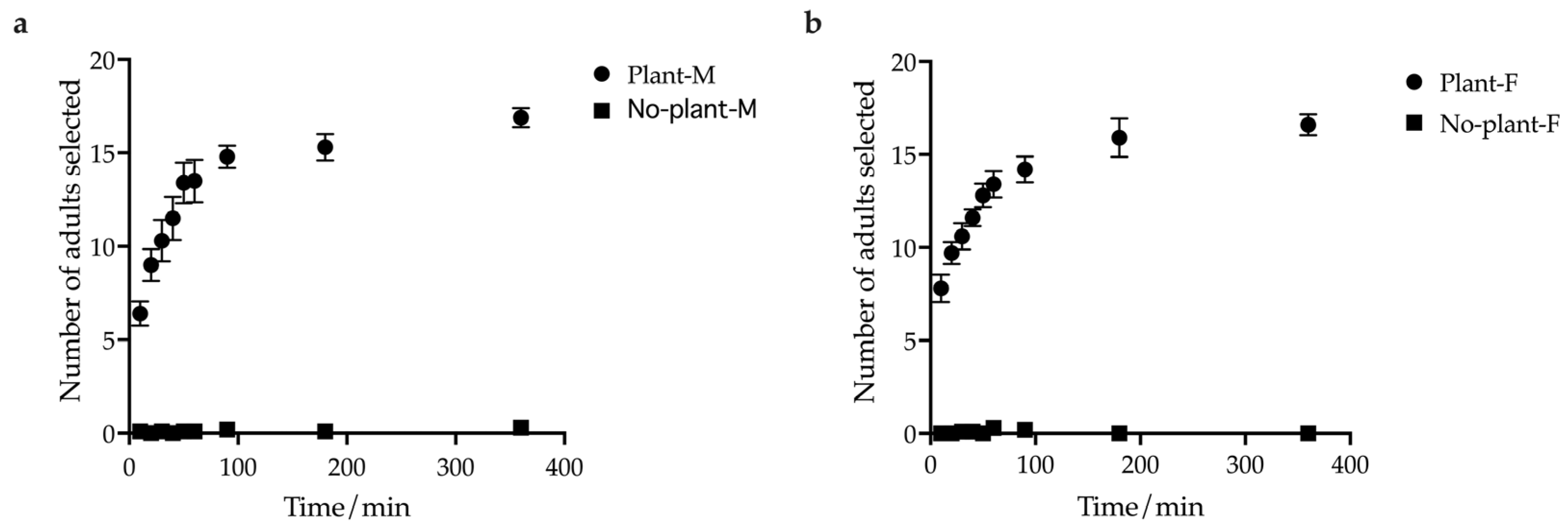
3.1. The Influence of Host Plants on the Aggregation Tendency of E. brandti Adults
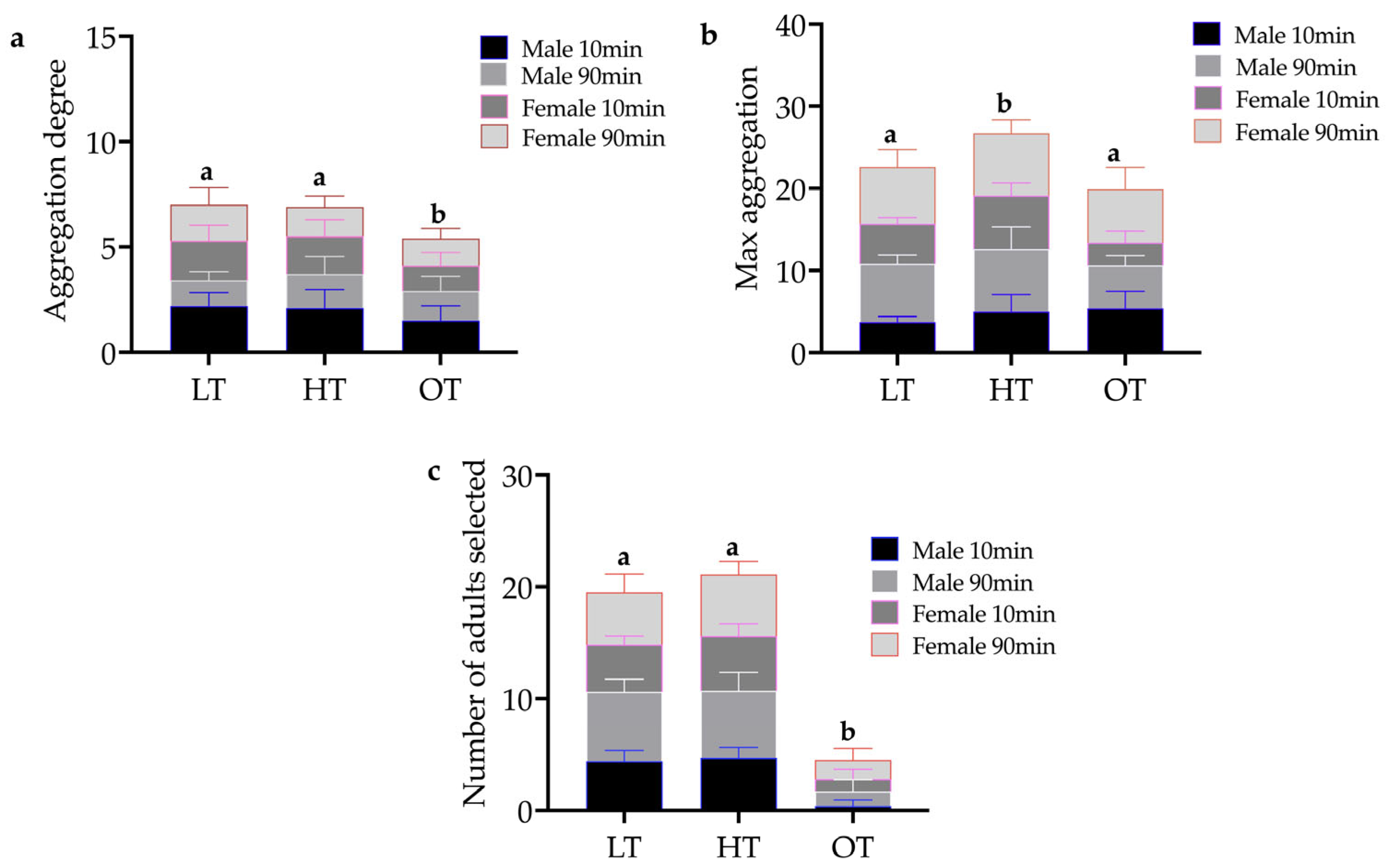
3.2. Effect of Temperature on Aggregation Behavior and the Tendency of E. brandti Adults
3.3. The Influence of Individual Interaction on the Aggregation Tendency of E. brandti Adults
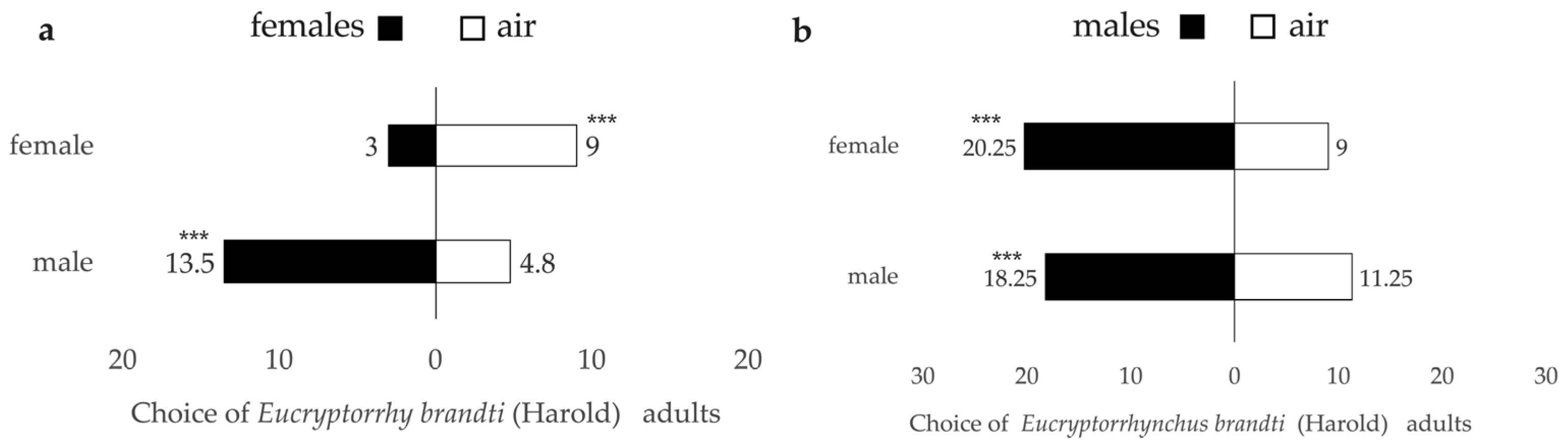
3.3.1. Genders’ Choice
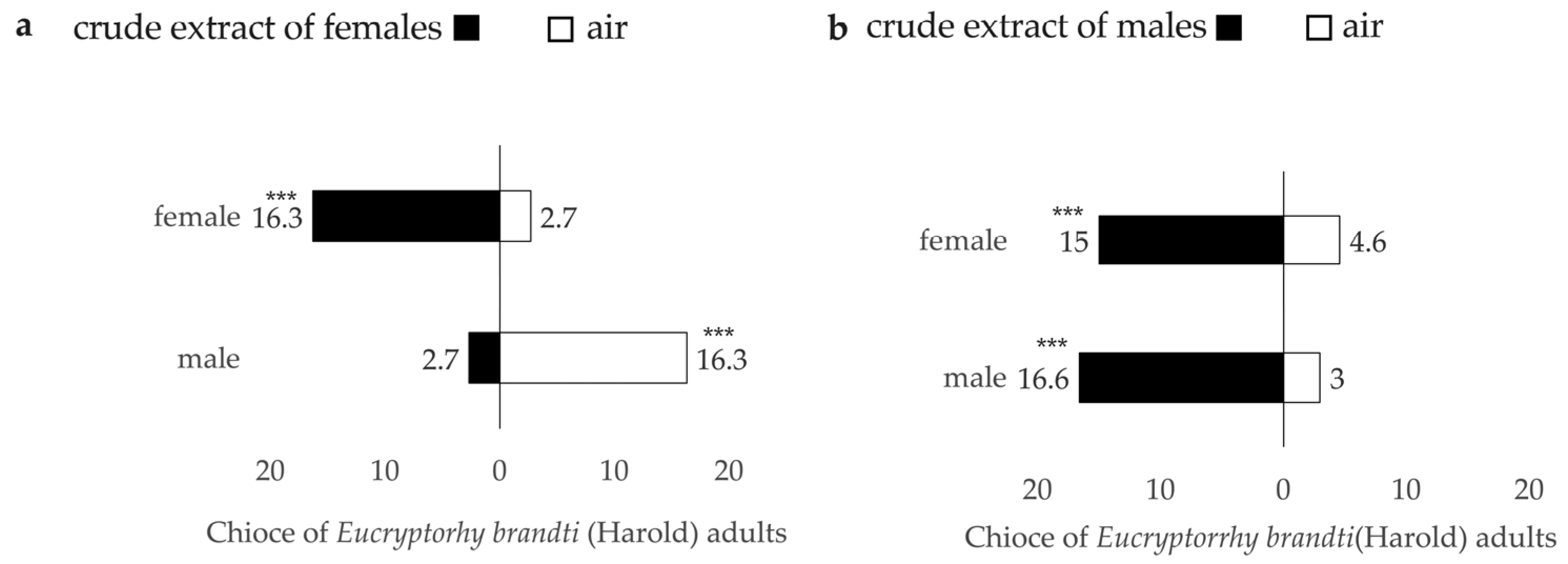
3.3.2. Responses to Intestinal Crude Extracts
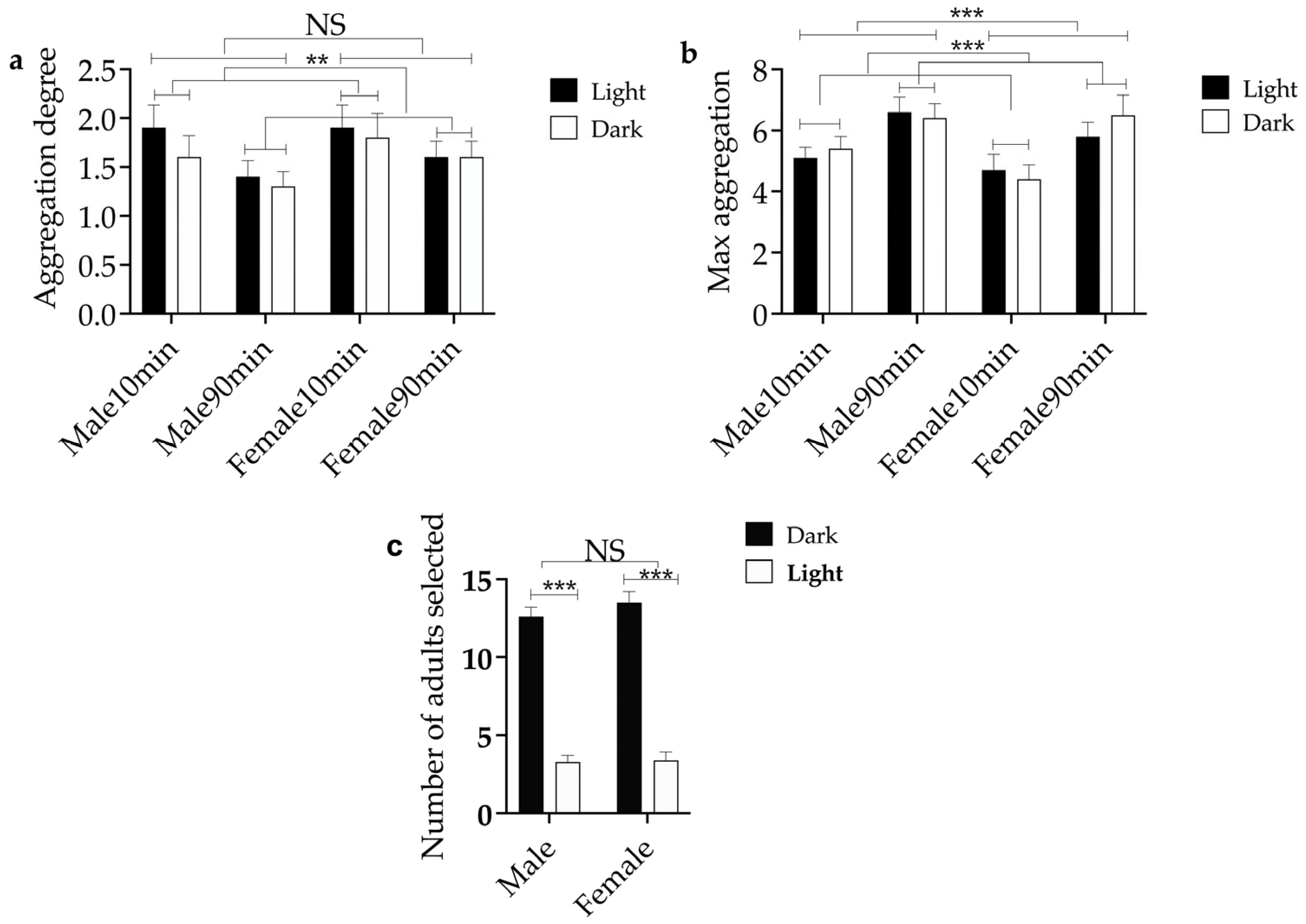
3.4. The Influence of Light Intensity on Aggregation Behavior and Aggregation Tendency of E. brandti Adults
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Qin, X.B.; Lai, B.M.; Li, D.J.; Liu, Y.Q.; Tan, R.S.; Sun, K.W. Bionomics and control of Eucryptorrhynchus brandti. Chin. For. Pest Dis. 1999, 5, 19–21. [Google Scholar]
- Herrick, N.J.; Mcavoy, T.; Snyder, A.L.; Salom, S. Host-Range Testing of Eucryptorrhynchus brandti (Coleoptera: Curculionidae), a Candidate for Biological Control of Tree-of-Heaven, Ailanthus altissima. Environ. Entomol. 2012, 41, 118–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, K.L.; Wen, X.J.; Ren, Y.; Wen, J.B. Novel trunk trap net designs for the control of Eucryptorrhynchus scrobiculatus (Coleoptera: Curculionidae). Pest Manag. Sci. 2019, 75, 2618–2626. [Google Scholar] [CrossRef] [PubMed]
- Ma, T.C.; Guo, W.J.; Wen, J.B. Effects of feeding on different parts of Ailanthus altissima on the intestinal microbiota of Eucryptorrhynchus scrobiculatus and Eucryptorrhynchus brandti (Coleoptera: Curculionidae). Front. Microbiol. 2022, 13, 2925. [Google Scholar] [CrossRef]
- Wang, X.; Ji, Y.C.; Wen, C.; Zhang, G.Y.; Wen, J.B. Effects of Trap Color and Shape on the Capture of Eucryptorrhynchus scrobiculatus (Coleoptera: Curculionidae). J. Econ. Entomol. 2019, 112, 2744–2750. [Google Scholar] [CrossRef]
- Wen, C.; Ji, Y.C.; Zhang, G.Y.; Tan, S.B.; Wen, J. Phototactic behaviour of Eucryptorrhynchus scrobiculatus and E. brandti (Coleoptera: Curculionidae) adults. Biocontrol Sci. Technol. 2018, 28, 544–561. [Google Scholar] [CrossRef]
- Qin, Y.N.; Zheng, H.Z.; Guo, W.J.; Yang, K.L.; Wen, J.B. Key factors for the selection of Eucryptorrhynchus brandti adults in overwintering site. J. Environ. Entomol. 2020, 42, 925–937. [Google Scholar] [CrossRef]
- Charabidze, D.; Bourel, B.; Gosset, D. Larval-mass effect: Characterisation of heat emission by necrophageous blowflies (Diptera: Calliphoridae) larval aggregates. Forensic Sci. Int. 2011, 211, 61–66. [Google Scholar] [CrossRef]
- Costa, J.T. The Other Insect Societies. J. Insect Conserv. 2008, 12, 579–580. [Google Scholar] [CrossRef]
- Wertheim, B.; van Baalen, E.-J.A.; Dicke, M.; Vet, L.E.M. Pheromone-mediated aggregation in nonsocial arthropods: An Evolutionary Ecological Perspective. Annu. Rev. Entomol. 2005, 50, 321–346. [Google Scholar] [CrossRef] [PubMed]
- Hopkins, G.; Lahanas, P. Aggregation behaviour in a neotropical dendrobatid frog (Allobates talamancae) in western Panama. Behaviour 2011, 148, 359–372. [Google Scholar] [CrossRef] [Green Version]
- Grieco, F. Aggregation of Eurasian Scops Owls Otus scops Breeding in Magpie Pica pica Nests. Ardea 2018, 106, 177–191. [Google Scholar] [CrossRef]
- Allee, W.C. Studies in animal aggregations: Causes and effects of bunching in land isopods. J. Exp. Zool. 1926, 45, 255–277. [Google Scholar] [CrossRef]
- Deneubourg, S.D.D.F.J.-L. A Basis for Spatial and Social Patterns in Ant Species: Dynamics and Mechanisms of Aggregation. J. Insect Behav. 2004, 17, 81–97. [Google Scholar]
- Maeno, K.O.; Ould Babah Ebbe, M.A. Aggregation Site Choice by Gregarious Nymphs of the Desert Locust, Schistocerca gregaria, in the Sahara Desert of Mauritania. Insects 2018, 9, 99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bonsignore, C.P.; Jones, T.M. Aggregation and mating success of Capnodis tenebrionis (Coleoptera: Buprestidae). Insect Sci. 2014, 21, 203–212. [Google Scholar] [CrossRef]
- Broly, P.; Mullier, R.; Deneubourg, J.L.; Devigne, C. Aggregation in woodlice: Social interaction and density effects. Zookeys 2012, 176, 133–144. [Google Scholar] [CrossRef] [Green Version]
- Jeanson, R.; Rivault, C.; Deneubourg, J.-L.; Blanco, S.; Fournier, R.; Jost, C.; Theraulaz, G. Self-organized aggregation in cockroaches. Anim. Behav. 2005, 69, 169–180. [Google Scholar] [CrossRef]
- Copp, N.H. Temperature-dependent behaviours and cluster formation by aggregating ladybird beetles. Anim. Behav. 1983, 31, 424–430. [Google Scholar] [CrossRef]
- Depickère, S.; Fresneau, D.; Deneubourg, J.-L. Effect of social and environmental factors on ant aggregation: A general response? J. Insect Physiol. 2008, 54, 1349–1355. [Google Scholar] [CrossRef] [PubMed]
- Sugita, N.; Ueda, K. The role of temperature on clustering behavior and mating opportunity in Bonin flying foxes. Mamm. Biol. 2013, 78, 455–460. [Google Scholar] [CrossRef]
- Tremblay, M.N.; Gries, G. Abiotic and Biotic Factors Affect Microhabitat Selection by the Firebrat, Thermobia domestica (Packard) (Thysanura: Lepismatidae). J. Insect Behav. 2006, 19, 321–335. [Google Scholar] [CrossRef]
- Iino, Y.; Yoshida, K. Parallel Use of Two Behavioral Mechanisms for Chemotaxis in Caenorhabditis elegans. J. Neurosci. 2009, 29, 5370–5380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Costa, J.T. The other insect societies: Overview and new directions. Curr. Opin. Insect Sci. 2018, 28, 40–49. [Google Scholar] [CrossRef] [PubMed]
- Durieux, D.; Fassotte, B.; Deneubourg, J.-L.; Brostaux, Y.; Vandereycken, A.; Joie, E.; Haubruge, E.; Verheggen, F.J. Aggregation behavior of Harmonia axyridis under non-wintering conditions. Insect Sci. 2015, 22, 670–678. [Google Scholar] [CrossRef] [PubMed]
- Cardé, R.T.; Baker, T.C. Sexual Communication with Pheromones. In Chemical Ecology of Insects; Springer: Boston, MA, USA, 1984; pp. 355–383. [Google Scholar]
- Sumpter, D.J.T. The principles of collective animal behaviour. Philos. Trans. R. Soc. B Biol. Sci. 2006, 361, 5–22. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramaswamy, S.B.; Jurenka, R.A.; Linn, C.E.; Roelofs, W.L. Evidence for the presence of a pheromonotropic factor in hemolymph and regulation of sex pheromone production in Helicoverpa zea. J. Insect Physiol. 1995, 41, 501–508. [Google Scholar] [CrossRef]
- Symonds, M.R.E.; Wertheim, B. The mode of evolution of aggregation pheromones in Drosophila species. J. Evol. Biol. 2005, 18, 1253–1263. [Google Scholar] [CrossRef]
- Addesso, K.M.; Mcauslane, H.J.; Cherry, R. Aggregation Behavior of the Southern Chinch Bug (Hemiptera: Blissidae). Environ. Entomol. 2012, 41, 887–895. [Google Scholar] [CrossRef] [Green Version]
- Bell, J.W. Searching Behavior Patterns in Insects. Annu. Rev. Entomol. 1990, 35, 447–467. [Google Scholar] [CrossRef]
- Reis, M.D.; Miller, D.M. Host Searching and Aggregation Activity of Recently Fed and Unfed Bed Bugs (Cimex lectularius L.). Insects 2011, 2, 186–194. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koenraadt, C.J.M.; Dicke, M. The role of volatiles in aggregation and host-seeking of the haematophagous poultry red mite Dermanyssus gallinae (Acari: Dermanyssidae). Exp. Appl. Acarol. 2010, 50, 191–199. [Google Scholar] [CrossRef]
- Wen, X.J.; Yang, K.L.; Piñero, J.C.; Wen, J.B. Contrasting Behavioral and Electrophysiological Responses of Eucryptorrhynchus scrobiculatus and E. brandti (Coleoptera: Curculionidae) to Volatiles Emitted from the Tree of Heaven, Ailanthus altissima. Insects 2021, 12, 68. [Google Scholar] [CrossRef] [PubMed]
- Bonduriansky, R. The evolution of male mate choice in insects: A synthesis of ideas and evidence. Biol. Rev. Camb. Philos. Soc. 2001, 76, S1464793101005693. [Google Scholar] [CrossRef] [Green Version]
- Thornhill, R.; Alcock, J. The Evolution of Insect Mating Systems; Harvard University Press: Cambridge, MA, USA, 1983. [Google Scholar]
- Burk, T. Signaling and Sex in Acalyptrate Flies. Fla. Entomol. 1981, 64, 30. [Google Scholar] [CrossRef]
- Leadbeater, E.; Chittka, L. Social Learning in Insects—From Miniature Brains to Consensus Building. Curr. Biol. 2007, 17, R703–R713. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marler, P. Animal Communication Signals. Science 1967, 157, 769–774. [Google Scholar] [CrossRef]
- Lihoreau, M.; Rivault, C. Tactile stimuli trigger group effects in cockroach aggregations. Anim. Behav. 2008, 75, 1965–1972. [Google Scholar] [CrossRef]
- Polajnar, J.; Maistrello, L.; Bertarella, A.; Mazzoni, V. Vibrational communication of the brown marmorated stink bug (Halyomorpha halys). Physiol. Entomol. 2016, 41, 249–259. [Google Scholar] [CrossRef]
- Varadínová, Z.; Stejskal, V.; Frynta, D. Patterns of aggregation behaviour in six species of cockroach: Comparing two experimental approaches. Entomol. Exp. Appl. 2010, 136, 184–190. [Google Scholar] [CrossRef]
- Turlings, T.C.J.; Davison, A.C.; Tamo, C. A six-arm olfactometer permitting simultaneous observation of insect attraction and odour trapping. Physiol. Entomol. 2004, 29, 45–55. [Google Scholar] [CrossRef]
- Suzuki, T. 4,8-Dimethyldecanal: The Aggregation Pheromone of the Flour Beetles, Tribolium castaneum and T. confusum (Coleoptera: Tenebrionidae). Agric. Biol. Chem. 1980, 44, 2519–2520. [Google Scholar] [CrossRef]
- Reis, A.C.; Neta, P.L.S.; Jordão, J.P.; Moura, J.I.L.; Vidal, D.M.; Zarbin, P.H.G.; Fávaro, C.F. Aggregation Pheromone of the Bearded Weevil, Rhinostomus barbirostris (Coleoptera: Curculionidae): Identification, Synthesis, Absolute Configuration and Bioactivity. J. Chem. Ecol. 2018, 44, 463–470. [Google Scholar] [CrossRef] [PubMed]
- Malo, E.A.; Gutiérrez-Escobar, V.; Castrejón-Ayala, F.; Rojas, J.C. The Aggregation Pheromone of Metamasius spinolae (Coleoptera: Dryophthoridae) Revisited: Less is More. Environ. Entomol. 2020, 49, 803–809. [Google Scholar] [CrossRef] [PubMed]
- Bandeira, P.T.; Fávaro, C.F.; Francke, W.; Bergmann, J.; Zarbin, P.H.G. Aggregation Pheromones of Weevils (Coleoptera: Curculionidae): Advances in the Identification and Potential Uses in Semiochemical-Based Pest Management Strategies. J. Chem. Ecol. 2021, 47, 968–986. [Google Scholar] [CrossRef]
- Larsson, M.C.; Svensson, G.P. Monitoring spatiotemporal variation in abundance and dispersal by a pheromone-kairomone system in the threatened saproxylic beetles Osmoderma eremita and Elater ferrugineus. J. Insect Conserv. 2011, 15, 891–902. [Google Scholar] [CrossRef]
- Conradt, L.; Roper, T.J. Consensus decision making in animals. Trends Ecol. Evol. 2005, 20, 449–456. [Google Scholar] [CrossRef]
- Goss, S.; Aron, S.; Deneubourg, J.L.; Pasteels, J.M. Self-organized shortcuts in the Argentine ant. Naturwissenschaften 1989, 76, 579–581. [Google Scholar] [CrossRef]
- Sumpter, D.J.T. Collective Animal Behavior; Princeton University Press: Princeton, NJ, USA, 2010. [Google Scholar]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Choice of E. brandti Adults | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Odor Source Types | Test Insects | Odor Source Tube | Air Tube | Unresponsive Insects | X2 | p | 1 Rr (%) | 1 SRr (%) | 1 SC |
| Female | female | 18 | 54 | 48 | 41.496 | <0.001 *** | 60.00 | 24.57 | −0.5086 |
| male | 81 | 29 | 8 | 91.67 | 73.74 | 0.4748 | |||
| Male | female | 79 | 36 | 5 | 0.998 | 0.339 | 95.83 | 68.79 | 0.3757 |
| male | 75 | 45 | 1 | 99.17 | 62.24 | 0.2447 | |||
| Choice of E. brandti Adults | |||||||||
|---|---|---|---|---|---|---|---|---|---|
| Odor Source Types | Test Insects | Odor Source Tube | Air Tube | Unresponsive Insects | X2 | p | 1 Rr (%) | 1 SRr (%) | 1 SC |
| 2 C. Female | female | 8 | 49 | 3 | 58.982 | <0.001 *** | 95.00 | 13.97 | 0.7207 |
| male | 49 | 8 | 3 | 95.00 | 86.12 | 0.7224 | |||
| 2 C. Male | female | 45 | 14 | 1 | 1.352 | 0.353 | 98.33 | 84.82 | 0.6950 |
| male | 50 | 9 | 1 | 98.33 | 76.32 | 0.5263 | |||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sun, X.; Song, W.; Guo, W.; Wang, S.; Wen, J. The Influencing Factors of Aggregation Behavior of Tree-of-Heaven Trunk Weevil, Eucryptorrhynchus brandti (Harold) (Coleoptera: Curculionidae). Insects 2023, 14, 253. https://doi.org/10.3390/insects14030253
Sun X, Song W, Guo W, Wang S, Wen J. The Influencing Factors of Aggregation Behavior of Tree-of-Heaven Trunk Weevil, Eucryptorrhynchus brandti (Harold) (Coleoptera: Curculionidae). Insects. 2023; 14(3):253. https://doi.org/10.3390/insects14030253
Chicago/Turabian StyleSun, Xuewen, Wei Song, Wenjuan Guo, Shujie Wang, and Junbao Wen. 2023. "The Influencing Factors of Aggregation Behavior of Tree-of-Heaven Trunk Weevil, Eucryptorrhynchus brandti (Harold) (Coleoptera: Curculionidae)" Insects 14, no. 3: 253. https://doi.org/10.3390/insects14030253