
Nutritional Value of Silkworm Pupae (Bombyx mori) with Emphases on Fatty Acids Profile and Their Potential Applications for Humans and Animals

Abstract
:Simple Summary
Abstract
1. Introduction
2. Data Sources
3. Nutritional Value of Silkworm Pupae
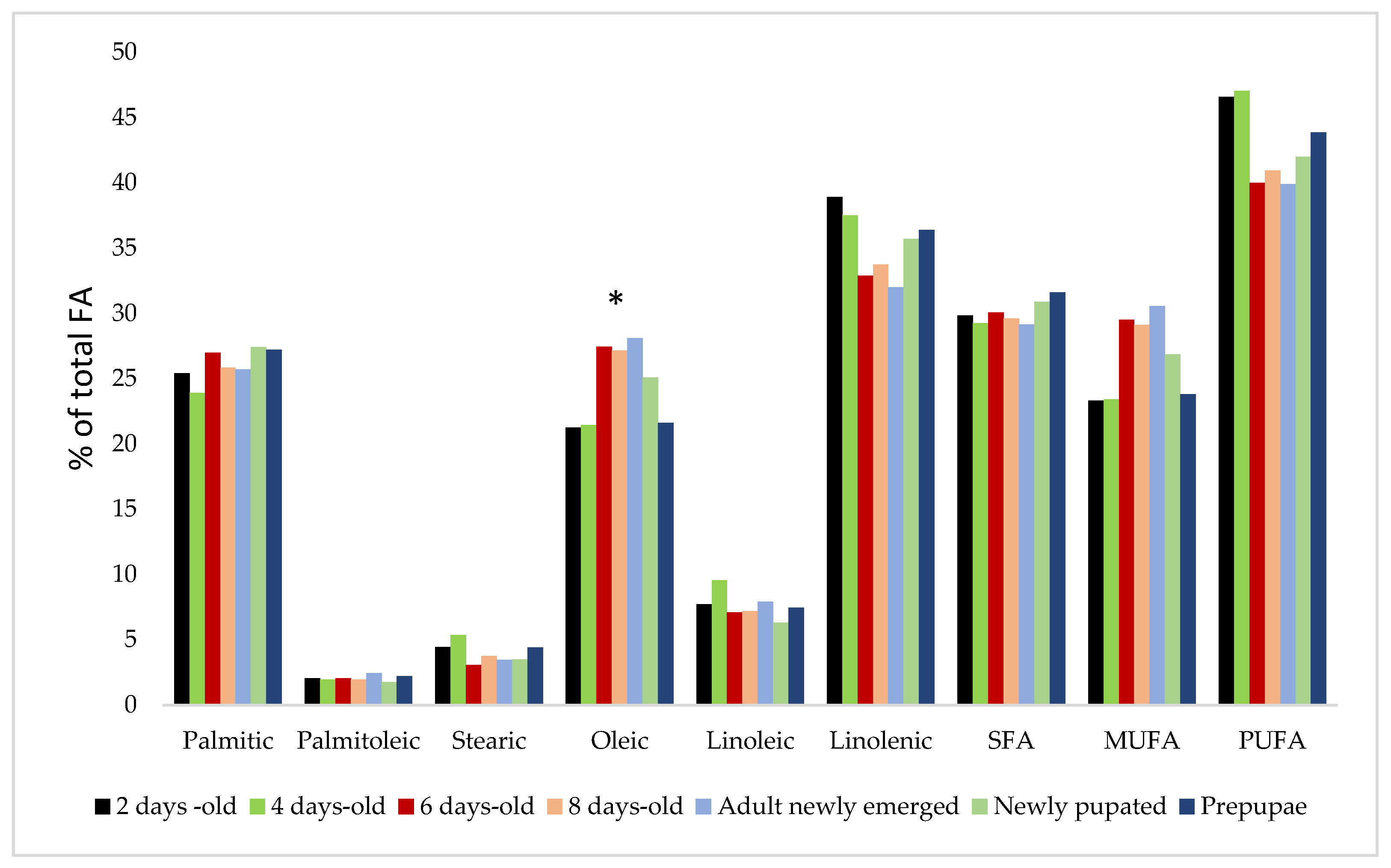
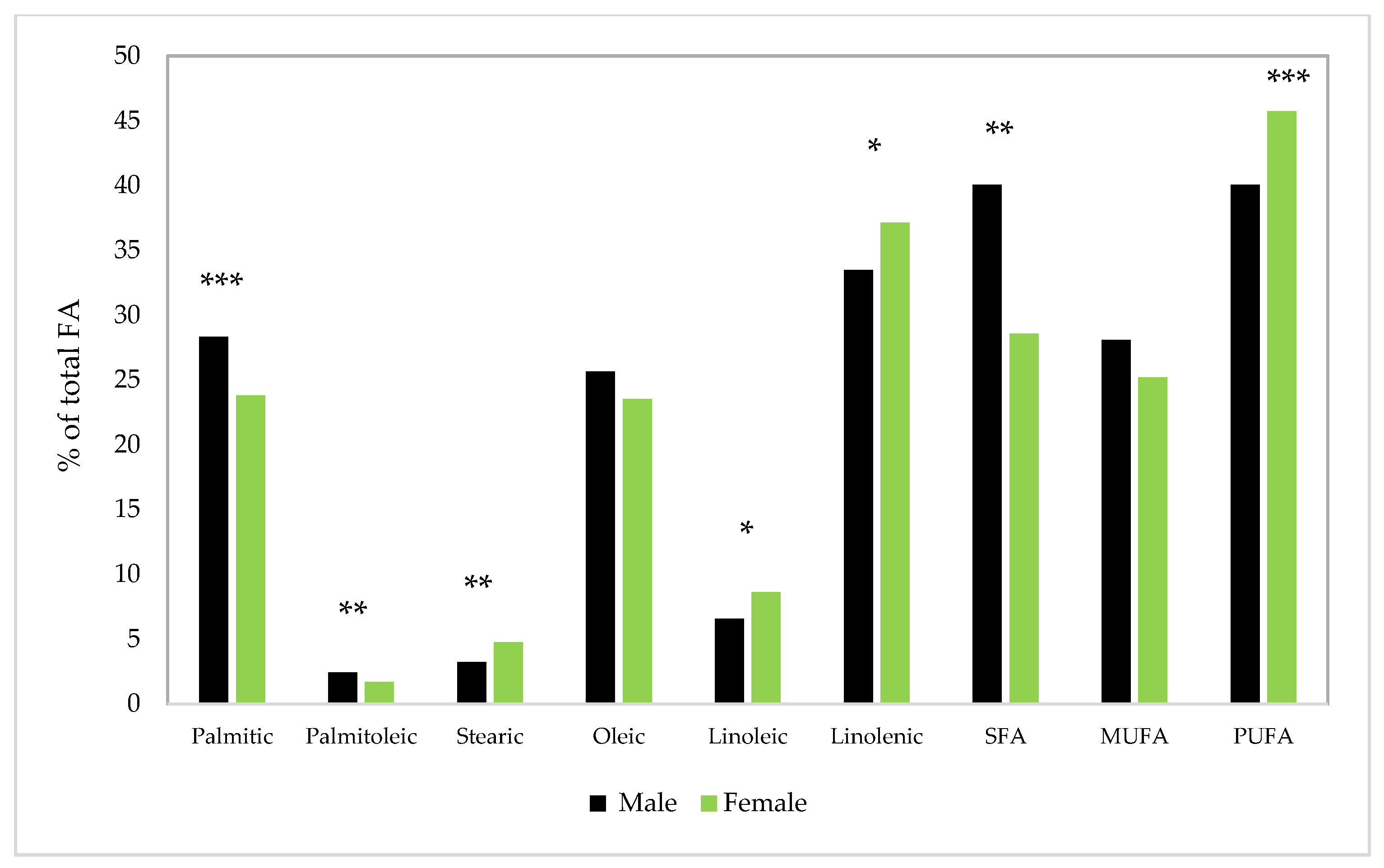
4. Silkworm Pupae Fatty Acids Composition
5. Role of Fatty Acids
6. Potential Applications of Silkworm Pupae
6.1. Application for Humans
6.2. Application in Animal Feeding
6.3. Environmental Effects
7. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Scrimgeour, C.; Gao, Y.; Oh, W.Y.; Shahidi, F. Edible Oil and Fat Products: Chemistry, Properties, and Safety Aspects. Chemistry of fatty acids. Chemistry of Fatty Acids. In Bailey’s Industrial Oil and Fat Products, 7th ed.; Shahidi, F., Ed.; Wiley Online Library: Hoboken, NJ, USA, 2005; pp. 1–40. [Google Scholar] [CrossRef]
- Calder, P.C. Mechanisms of Action of (n-3) Fatty Acids. J. Nutr. 2012, 142, 592S–599S. [Google Scholar] [CrossRef] [Green Version]
- Ohwada, T.; Yokokawa, T.; Kanno, Y.; Hotsuki, Y.; Sakamoto, T.; Watanabe, K.; Nakazato, K.; Takeishi, Y. Vascular composition data supporting the role of N-3 polyunsaturated fatty acids in the prevention of cardiovascular disease events. Data Brief 2016, 7, 1237–1247. [Google Scholar] [CrossRef] [Green Version]
- Bartkiene, E.; Bartkevics, V.; Berzina, Z.; Klementaviciute, J.; Sidlauskiene, S.; Isariene, A.; Zeimiene, V.; Lele, V.; Mozuriene, E. Fatty acid profile and safety aspects of the edible oil prepared by artisans at small-scale agricultural companies. Food Sci. Nutr. 2021, 9, 5402–5414. [Google Scholar] [CrossRef]
- Kumar, R.V.; Srivastava, D.; Kumar, U.; Kumar, M.; Singh, P. Bioprospecting of omega-3 fatty acid from silkworm pupal oil: From molecular mechanism to biological activities. J. Biol. Act. Prod. Nat. 2021, 10, 495–506. [Google Scholar] [CrossRef]
- Orsavova, J.; Misurcova, L.; Ambrozova, J.V.; Vicha, R.; Mlcek, J. Fatty Acids Composition of Vegetable Oils and Its Contribution to Dietary Energy Intake and Dependence of Cardiovascular Mortality on Dietary Intake of Fatty Acids. Int. J. Mol. Sci. 2015, 16, 12871–12890. [Google Scholar] [CrossRef]
- Vasseghian, Y.; Moradi, M.; Dragoi, E.-N.; Khaneghah, A.M. A review on mycotoxins detection techniques in edible oils. Int. J. Environ. Anal. Chem. 2020, 102, 2125–2139. [Google Scholar] [CrossRef]
- Hăbeanu, M.; Hebean, V.; Taranu, I.; Ropota, M.; Lefter, N.A.; Marin, D. Dietary Ecologic Camelina Oil—A Beneficial Source of N-3 PUFA In Muscle Tissue and Health Status in Finishing Pig. Rom. Biotechnol. Lett. 2011, 16, 6564–6571. [Google Scholar]
- Hăbeanu, M.; Lefter, N.A.; Gheorghe, A.; Nagy, A.; Marin, D.; Ropotă, M. Effects of dietary flaxseed oil on the muscle fatty acid composition in Mangalitsa pigs in an extensive rearing system. S. Afr. J. Sci. 2014, 44, 240–244. [Google Scholar] [CrossRef] [Green Version]
- Hăbeanu, M.; Lefter, N.A.; Gheorghe, A.; Untea, A.; Ropotă, M.; Grigore, D.-M.; Varzaru, I.; Toma, S.M. Evaluation of Performance, Nitrogen Metabolism and Tissue Composition in Barrows Fed an n-3 PUFA-Rich Diet. Animals 2019, 9, 234. [Google Scholar] [CrossRef] [Green Version]
- Hăbeanu, M.; Lefter, N.A.; Gheorghe, A.; Ropota, M.; Toma, S.M.; Pistol, G.C.; Surdu, I.; Dumitru, M. Alterations in essential fatty acids, immunoglobulins (IgA, IgG and IgM), and enteric methane emission in primiparous sows fed hemp seed oil and their offspring response. Vet. Sci. 2022, 9, 352. [Google Scholar] [CrossRef]
- Frančáková, H.; Ivanišová, E.; Dráb, Š.; Krajčovič, T.; Tokár, M.; Mareček, J.; Musilová, J. Composition of Fatty Acids in Selected Vegetable Oils. Potravin. Slovak J. Food Sci. 2015, 9, 538–542. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Zhao, W.; Lai, Y.; Zhang, B.; Zhang, D. Edible Plant Oil: Global Status, Health Issues, and Perspectives. Front. Plant Sci. 2020, 11, 1315. [Google Scholar] [CrossRef]
- Longvah, T.; Mangthya, K.; Ramulu, P. Nutrient composition and protein quality evaluation of eri silkworm (Samia ricinii) prepupae and pupae. Food Chem. 2011, 128, 400–403. [Google Scholar] [CrossRef] [PubMed]
- Mokaya, H.O.; Ndunda, R.M.; Kegode, T.M.; Koech, S.J.; Tanga, C.M.; Subramanian, S.; Ngok, B. Silkmoth pupae: Potential and less exploited alternative source of nutrients and natural antioxidants. J. Insects Food Feed 2022, in press. [Google Scholar] [CrossRef]
- Ordoñez-Araque, R.; Quishpillo-Miranda, N.; Ramos-Guerrero, L. Edible Insects for Humans and Animals: Nutritional Composition and an Option for Mitigating Environmental Damage. Insects 2022, 13, 944. [Google Scholar] [CrossRef]
- Zhou, Y.; Zhou, S.; Duan, H.; Wang, J.; Yan, W. Silkworm Pupae: A Functional Food with Health Benefits for Humans. Foods 2022, 11, 1594. [Google Scholar] [CrossRef]
- Yoon, S.; Wong, A.K.N.; Chae, M.; Auh, J.-H. Comparative Characterization of Protein Hydrolysates from Three Edible Insects: Mealworm Larvae, Adult Crickets, and Silkworm Pupae. Foods 2019, 8, 563. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; He, K.; Cirkovic Velickovic, T.; Liu, Z. Nutritional, functional, and allergenic properties of silkworm pupae. Food Sci. Nutr. 2021, 9, 4655–4665. [Google Scholar] [CrossRef]
- Dong, H.L.; Zhang, S.X.; Tao, H.; Chen, Z.H.; Li, X.; Qiu, J.F.; Cui, W.Z.; Sima, Y.H.; Cui, W.Z.; Xu, S.Q. Metabolomics differences between silkworms (Bombyx mori) reared on fresh mulberry (Morus) leaves or artificial diets. Sci. Rep. 2017, 7, 1–16. [Google Scholar] [CrossRef] [Green Version]
- Lamberti, C.; Gai, F.; Cirrincione, S.; Giribaldi, M.; Purrotti, M.; Manfredi, M.; Marengo, E.; Sicuro, B.; Saviane, A.; Cappellozza, S.; et al. Investigation of the protein profile of silkworm (Bombyx mori) pupae reared on a well-calibrated artificial diet compared to mulberry leaf diet. PeerJ 2019, 7, e6723. [Google Scholar] [CrossRef] [Green Version]
- Herman, R.A.; Yan, C.-H.; Wang, J.-Z.; Xun, X.-M.; Wu, C.-K.; Li, Z.-N.; Ayepa, E.; You, S.; Gong, L.-C.; Wang, J. Insight into the silkworm pupae: Modification technologies and functionality of the protein and lipids. Trends Food Sci. Tech. 2022, 129, 408–420. [Google Scholar] [CrossRef]
- Kumar, D.; Dev, P.; Kumar, R.V. Biomedical Applications of Silkworm Pupae Proteins. Biochemistry and Molecular Biology. In Biomedical Applications of Natural Proteins; Springer: Berlin/Heidelberg, Germany, 2015; Chapter 3; pp. 41–49. [Google Scholar] [CrossRef]
- Tomotake, H.; Katagiri, M.; Yamato, M. Silkworm Pupae (Bombyx mori) Are New Sources of High-Quality Protein and Lipid. J. Nutr. Sci. Vitaminol. 2010, 56, 446–448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kouřimská, L.; Adámkov, A. Nutritional and sensory quality of edible insects. NFS J. 2016, 4, 22–26. [Google Scholar] [CrossRef] [Green Version]
- Roychoudhury, N.; Mishra, R.K. Silkworm as human food. Van Sangyan 2020, 7, 35–39. [Google Scholar]
- Meyer-Rochow, V.B.; Gahukar, R.T.; Ghosh, S.; Jung, C. Chemical composition, nutrient quality and acceptability of edible insects are affected by species, developmental stage, gender, diet, and processing method. Foods 2021, 10, 1036. [Google Scholar] [CrossRef]
- Shukurova, Z.Y.; Khalilov, Z.M.; Shukurlu, I.H. Study of the organic and mineral composition of living pupae of the wild silkworm Saturnia pyri for use as food additives. Int. J. Indust. Entomol. 2021, 43, 52–58. [Google Scholar] [CrossRef]
- Zotte, A.D.; Singh, Y.; Squartini, A.; Stevanato, P.; Cappellozza, S.; Kovitvadhi, A.; Subaneg, S.; Bertelli, D.; Cullere, M. Effect of a dietary inclusion of full-fat or defatted silkworm pupa meal on the nutrient digestibility and faecal microbiome of fattening quails. Animal 2021, 15, 100112. [Google Scholar] [CrossRef] [PubMed]
- Nowak, V.; Persijn, D.; Rittenschober, D.; Charrondiere, U.R. Review of food composition data for edible insects. Food Chem. 2014, 193, 39–46. [Google Scholar] [CrossRef]
- Pereira, N.R.; Ferrarese-Filho, O.; Matsushita, M.; de Souza, N.E. Proximate composition and fatty acid profile of Bombyx mori L. chrysalis toast. J. Food Compos. Anal. 2003, 16, 451–457. [Google Scholar] [CrossRef]
- Singh, K.P.; Jayasomu, R.S. Bombyx mori—A Review of its Potential as a Medicinal Insect. Pharm. Biol. 2002, 40, 28–32. [Google Scholar] [CrossRef]
- Feedpedia. Animal Feed Resources Information System. Available online: https://www.feedipedia.org/node/199 (accessed on 10 January 2023).
- Rumpold, B.A.; Schlüter, O.K. Nutritional composition and safety aspects of edible insects. Mol. Nutr. Food Res. 2013, 57, 802–823. [Google Scholar] [CrossRef]
- Raja, P.K.; Aanand, S.; Sampathkumar, S.J.; Padmavathy, P. Silkworm pupae meal as alternative source of protein in fish feed. J. Entomol. Zool. Stud. 2019, 7, 78–85. [Google Scholar]
- Jeyaprakashsabari, S.; Aanand, S. Silkworm Pupae Meal—A Promising Fish Meal Substitute in Aqua Feed. AgriCos e-Newsl. 2021, 2, 17. [Google Scholar]
- Altomare, A.A.; Baron, G.; Aldini, G.; Carini, M.; D’Amato, A. Silkorm pupae as source of high-value edible proteins and bioactive peptides. Food Sci. Nutr. 2020, 8, 2652–2661. [Google Scholar] [CrossRef] [PubMed]
- Zhou, J.; Han, D. Proximate, amino acid and mineral composition of pupae of the silkworm Antheraea pernyi in China. J. Food Compos. Anal. 2006, 19, 850–853. [Google Scholar] [CrossRef]
- Kotake-Nara, E.; Yamamoto, K.; Nozawa, M.; Miyashita, K.; Murakami, T. Lipid Profiles and Oxidative Stability of Silkworm Pupal Oil. J. Oleo Sci. 2002, 51, 681–690. [Google Scholar] [CrossRef] [Green Version]
- Longvah, T.; Manghtya, K.; Qadri, S.S.Y.H. Eri silkworm: A source of edible oil with a high content of α-linolenic acid and of significant nutritional value. J. Sci. Food Agric. 2012, 92, 1988–1993. [Google Scholar] [CrossRef]
- Ray, M.; Gangopadhyay, D. Effect of maturation stage and sex on proximate, fatty acid and mineral composition of eri silkworm (Samia ricini) from India. J. Food Compos. Anal. 2021, 100, 103898. [Google Scholar] [CrossRef]
- Mahesh, D.S.; Vidhathri, B.S.; Narayanaswamy, T.K.; Subbarayappa, C.T.; Muthuraju, R.; Shruthi, P. A Review—Bionutritional Science of Silkworm Pupal residue to Mine New ways for utilization. Int. J. Adv. Res. Biol. Sci. 2015, 2, 135–140. [Google Scholar]
- Nakasone, S.; Ito, T. Fatty acid composition of the silkworm, Bombyx mori L. J. Insect Physiol. 1967, 13, 1237–1246. [Google Scholar] [CrossRef]
- Tangsanthatkun, J.; Peanparkdee, M.; Katekhong, W.; Harnsilawat, T.; Tan, C.P.; Klinkesorn, U. Application of Aqueous Saline Process to Extract Silkworm Pupae Oil (Bombyx mori): Process Optimization and Composition Analysis. Foods 2022, 11, 291. [Google Scholar] [CrossRef] [PubMed]
- Yu, X.-B.; Shen, Y.-Y.; Cui, Q.-M.; Chen, Y.; Sun, W.; Huang, X.-Z.; Zhu, Y. Silkworm (Bombyx mori) has the capability to accumulate C20 and C22 polyunsaturated fatty acids. Eur. J. Lipid Sci. Technol. 2018, 120, 1700268. [Google Scholar] [CrossRef]
- Glick, N.R.; Fischer, M.H. The Role of Essential Fatty Acids in Human Health. J. Evid-Based Complement. Altern. Med. 2013, 18, 268–289. [Google Scholar] [CrossRef]
- Park, S.; Lee, J.-J.; Lee, J.; Lee, J.K.; Byun, J.; Kim, I.; Ha, J.-H. Lowering n-6/n-3 Ratio as an Important Dietary Intervention to Prevent LPS-Inducible Dyslipidemia and Hepatic Abnormalities in ob/ob Mice. Int. J. Mol. Sci. 2022, 23, 6384. [Google Scholar] [CrossRef] [PubMed]
- Burns-Whitmore, B.; Froyen, E.; Heskey, C.; Parker, T.; San Pablo, G. Alpha-Linolenic and Linoleic Fatty Acids in the Vegan Diet: Do They Require Dietary Reference Intake/Adequate Intake Special Consideration? Nutrients 2019, 11, 2365. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Zhang, J.-L.; Wu, F.-A. Enrichment process for a-linolenic acid from silkworm pupae oil. Eur. J. Lipid Sci. Technol. 2013, 115, 791–799. [Google Scholar] [CrossRef]
- Suzuki, N.; Sawada, K.; Takahashi, I.; Matsuda, M.; Fukui, S.; Tokuyasu, H.; Shimizu, H.; Yokoyama, J.; Akaike, A.; Nakaji, S. Association between polyunsaturated fatty acid and reactive oxygen species production of neutrophils in the general population. Nutrients 2020, 12, 3222. [Google Scholar] [CrossRef] [PubMed]
- Petermann, A.B.; Reyna-Jeldes, M.; Ortega, L.; Coddou, C.; Yévenes, G.E. Roles of the Unsaturated Fatty Acid Docosahexaenoic Acid in the Central Nervous System: Molecular and Cellular Insights. Int. J. Mol. Sci. 2022, 23, 5390. [Google Scholar] [CrossRef]
- Bourre, J.M.; Bonneil, M.; Clément, M.; Dumont, O.; Durand, G.; Lafont, H.; Nalbone, G.; Piciotti, M. Function of dietary polyunsaturated fatty acids in the nervous system. Prostaglandins Leukot. Essent. Fat. Acids 1993, 48, 5–15. [Google Scholar] [CrossRef]
- Williams, C.M. Dietary fatty acids and human health. Anim. Res. 2000, 49, 165–180. [Google Scholar] [CrossRef] [Green Version]
- Alessandri, J.M.; Guesneta, P.; Vancassel, S.; Astorg, P.; Denisa, I.; Langelier, B.; Aïd, S.; Poumès-Ballihaut, C.; Champeil-Potokar, G.; Laviall, M. Polyunsaturated fatty acids in the central nervous system: Evolution of concepts and nutritional implications throughout life. Reprod. Nutr. Dev. 2004, 44, 509–538. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rangacharyulu, P.V.; Giri, S.S.; Paul, B.N.; Yashoda, K.P.; Rao, R.J.; Mahendrakar, N.S.; Mohanty, S.N.; Mukhopadhyay, P.K. Utilization of fermented silkworm pupae silage in feed for carps. Bioresour. Technol. 2003, 86, 29–32. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.J.; Lee, K.P.; Lee, D.Y.; Kim, Y.T.; Baek, S.; Yoon, M.S. Inhibitory effect of modified silkworm pupae oil in PDGF-BB-induced proliferation and migration of vascular smooth muscle cells. Food Sci. Biotechnol. 2020, 29, 1091–1099. [Google Scholar] [CrossRef] [PubMed]
- Saviane, A.; Tassoni, L.; Naviglio, D.; Lupi, D.; Savoldelli, S.; Bianchi, G.; Cortellino, G.; Bondioli, P.; Folegatti, L.; Casartelli, M.; et al. Mechanical Processing of Hermetia illucens Larvae and Bombyx mori Pupae Produces Oils with Antimicrobial Activity. Animals 2021, 11, 783. [Google Scholar] [CrossRef] [PubMed]
- Ryu, S.P. Silkworm pupae powder ingestion increases fat metabolism in swim-trained rats. J. Exerc. Nutr. Biochem. 2014, 18, 141–149. [Google Scholar] [CrossRef] [Green Version]
- Long, X.; Song, J.; Zhao, X.; Zhang, Y.; Wang, H.; Liu, X.; Suo, H. Silkworm pupa oil attenuates acetaminophen-induced acute liver injury by inhibiting oxidative stress-mediated NF-κB signaling. Food Sci. Nutr. 2020, 8, 237–245. [Google Scholar] [CrossRef] [PubMed]
- Long, X.; Zhao, X.; Wang, W.; Zhang, Y.; Wang, H.; Liu, X.; Suo, H. Protective effect of silkworm pupa oil on hydrochloric acid/ethanol-induced gastric ulcers. J. Sci. Food Agric. 2019, 99, 2974–2986. [Google Scholar] [CrossRef]
- Wattanathorn, J.; Muchimapura, S.; Boosel, A.; Kongpa, S.; Kaewrueng, W.; Tong-Un, T.; Wannanon, P.; Thukhammee, W. Silkworm pupae protect against Alzheimer’s disease. Am. J. Agric. Biol. Sci. 2012, 7, 330–336. [Google Scholar] [CrossRef] [Green Version]
- Baek, S.Y.; Li, F.Y.; Kim, J.H.; Ahn, C.; Kim, H.J.; Kim, M.R. Protein hydrolysate of silkworm pupa prevents memory impairment induced by oxidative stress in scopolamine-induced mice via modulating the cholinergic nervous system and antioxidant defense system. Prev. Nutr. Food Sci. 2020, 25, 389–399. [Google Scholar] [CrossRef]
- Manosroi, A.; Boonpisuttinant, K.; Winitchai, S.; Manosroi, W.; Manosroi, J. Free radical scavenging and tyrosinase inhibition activity of oils and sericin extracted from Thai native silkworms (Bombyx mori). Pharm. Biol. 2010, 48, 855–860. [Google Scholar] [CrossRef] [Green Version]
- Majumder, U.K.; Sengupta, A. Triglyceride composition of chrysalis oil, an insect lipid. J. Am. Oil Chem. Soc. 1997, 56, 620–623. [Google Scholar] [CrossRef]
- Deori, M.; Boruah, D.C.; Devi, D.; Devi, R. Antioxidant and antigenotoxic effects of pupae of the muga silkworm Antheraea assamensis. Food Biosci. 2014, 5, 108–114. [Google Scholar] [CrossRef]
- Łochynska, M.; Frankowski, J. The biogas production potential from silkworm waste. Waste Manag. 2018, 79, 564–570. [Google Scholar] [CrossRef]
- World Health Organization (WHO). Healthy Diet: Fact Sheet. 2015. Available online: http://www.who.int/nutrition/publications/nutrientrequirements/healthydiet_factsheet394.pdf?ua¼1 (accessed on 15 January 2023).
- Ji, K.M.; Zhan, Z.K.; Chen, J.J.; Liu, Z.G. Anaphylactic shock caused by silkworm pupa consumption in China. Allergy 2008, 63, 1407–1408. [Google Scholar] [CrossRef]
- Choi, G.S.; Shin, Y.S.; Kim, J.E.; Ye, Y.M.; Park, H.S. Five cases of food allergy to vegetable worm (Cordyceps sinensis) showing cross-reactivity with silkworm pupae. Allergy 2010, 65, 1196–1197. [Google Scholar] [CrossRef]
- Feng, Y.; Chen, X.M.; Zhao, M.; He, Z.; Sun, L.; Wang, C.Y.; Ding, D.F. Edible insects in China: Utilization and prospects. Insect Sci. 2018, 25, 184–198. [Google Scholar] [CrossRef]
- Sheikh, I.U.; Banday, M.T.; Baba, I.A.; Adil, S.; Shaista, S.N.; Bushra, Z.; BulbuI, K.H. Utilization of silkworm pupae meal as an alternative source of protein in the diet of livestock and poultry: A review. J. Entomol. Zool. Stud. 2018, 6, 1010–1016. [Google Scholar]
- Asimi, O.A.; Bhat, T.H.; Nasir, H.; Irfan, K. Alternative Source of Protein “Silkworm Pupae” (Bombyx mori) In Coldwater Aquaculture. Int. J. Poult. Sci. 2017, 1, 1–4. [Google Scholar] [CrossRef] [Green Version]
- Matthäus, B.; Özcan, M.M. Fatty Acid Composition, Tocopherol and Sterol Contents in Linseed (Linum usitatissimum L.) Varieties. Iran. J. Chem. Chem. Eng. 2017, 36, 147–152. [Google Scholar] [CrossRef]
- Ostrikov, A.N.; Kleimenova, N.L.; Kopylov, M.V.; Bolgova, I.N. The study of the fatty acid composition of camelina oil obtained by cold pressing. IOP Publ. Conf. Ser. Earth Environ. Sci. 2021, 640, 042009. [Google Scholar] [CrossRef]
- Valerie, H.; Tran, G.; Giger-Reverdin, S.; Lebas, F.; Silkworm Pupae Meal. Feedipedia, a Programme by INRA, CIRAD, AFZ and FAO. 2015. Available online: http://www.feedipedia.org/node/199 (accessed on 12 December 2022).
- Fagoonee, I. Possible growth factors for chickens in silkworm pupae meal. Brit. Poult. Sci. 1983, 24, 295–300. [Google Scholar] [CrossRef] [PubMed]
- Priyadharshini, P.; Joncy, M.A.; Saratha, M. Industrial utilization of silkworm pupae—A review. J. Int. Acad. Res. Multidiscip. 2017, 5, 62–70. [Google Scholar]
- Miah, M.; Singh, Y.; Cullere, M.; Tenti, S.; Zotte, A.D. Effect of dietary supplementation with full-fat silkworm (Bombyx mori L.) chrysalis meal on growth performance and meat quality of Rhode Island Red Fayoumi crossbred chickens. Ital. J. Anim. Sci. 2020, 19, 447–456. [Google Scholar] [CrossRef]
- Shakoori, M.; Gholipour, M.; Naseri, S. Effect of replacing dietary fish meal with silkworm (Bombyx mori) pupae on hematological parameters of rainbow trout Oncorhynchus mykiss. Comp. Clin. Path. 2014, 24, 139–143. [Google Scholar] [CrossRef]
- Giacomin, A.M.; Garcia, J.B., Jr.; Zonatti, W.F.; Silva-Santos, M.C.; Laktim, M.C.; Baruque-Ramos, J. Silk industy and carbon footprint mitigation. IOP Publ. Conf. Ser. Mater. Sci. Eng. 2017, 254, 192008. [Google Scholar] [CrossRef] [Green Version]
- Thirumalaisamy, G.; Malik, P.K.; Trivedi, S.; Kolte, A.P.; Dhali, A.; Bhatta, R. Effect of silkworm (Bombyx mori) pupae oil supplementation on enteric methane emission and methanogens diversity in sheep. Anim. Biotechnol. 2022, 33, 128–140. [Google Scholar] [CrossRef]
- Belanche, A.; de la Fuente, G.; Charles, J.N. Study of methanogen communities associated with different rumen protozoal populations. FEMS Microbiol. Ecol. 2014, 90, 663–677. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | % (DM Bases) | Reference |
|---|---|---|
| Protein | 48–60 | Herman et al. [22] |
| 49–54 | Wu et al. [19] | |
| 59.8–75.1 | Shukurova et al. [28] | |
| 52–80 | Zotte et al. [29] | |
| 59.52–94.98 | Lamberti et al. [21] * | |
| 55.60 | Kumar et al. [23] | |
| 49–54 | Nowak et al. [30] | |
| 55.6 | Tomotake et al. [24] | |
| 49.1–53.5 | Pereira et al. [31] | |
| 48.7–51.6 | Singh and Jayasomu [32] | |
| 58.8 | Feedpedia [33] | |
| 48.7 | Rumpold et. al. [34] | |
| Fat | 30 | Herman et al. [22] |
| 25–30 | Wu et al. [19] | |
| 12.1–27.4 | Shukurova et al. [28] | |
| 29 | Kouřimská and Adámkov [25] | |
| 32.2 | Kumar et al. [23] | |
| 32.2 | Tomotake et al. [24] | |
| 33.3–35.7 | Pereira et al. [31] | |
| 30 | Singh and Jayasomu [32] | |
| 28.5 | Feedpedia [33] | |
| 30.10 | Rumpold et. al. [34] | |
| Fiber | 3.5–4.7 | Shukurova et al. [28] |
| 14 | Kouřimská and Adámkov [25] | |
| 5.8 | Feedpedia [33] | |
| Energy | 5.09–6.82 MJ/kg | Lamberti et al. [21] |
| 26.5 MJ/kg DM | Feedpedia [33] |
| Fatty Acids (%) | Male | Female | Total | Reference |
|---|---|---|---|---|
| C16:0 (palmitic) | 28.60 | 22.80 | Nakasone and Ito [43] | |
| 24.90 | 19.50 | Kotake-Nara et al. [39] | ||
| 21.3–28.30 | Pereira et al. [31] | |||
| 24.20 | Tomotake et al. [24] | |||
| 23.18 | Kumar et al. [5] | |||
| 23.18 | Zhou et al. [17] | |||
| C16:1n-7 (palmitoleic) | 3.10 | 1.80 | Nakasone and Ito [43] | |
| 0.80 | 0.60 | Kotake-Nara et al. [39] | ||
| 0.60–0.70 | Pereira et al. [31] | |||
| 1.70 | Tomotake et al. [24] | |||
| 1.07 | Kumar et al. [5] | |||
| 1.07 | Zhou et al. [17] | |||
| C18:0 (stearic) | 2.60 | 4.30 | Nakasone and Ito [43] | |
| 5.40 | 6.30 | Kotake-Nara et al. [39] | ||
| 5.5–9.20 | Pereira et al. [31] | |||
| 4.50 | Tomotake et al. [24] | |||
| 4.69 | Kumar et al. [5] | |||
| 4.69 | Zhou et al. [17] | |||
| C18:1n-9 (oleic) | 29.0 | 27.20 | Nakasone and Ito [43] | |
| 24.30 | 22.60 | Kotake-Nara et al. [39] | ||
| 30.6–38.0 | Pereira et al. [31] | |||
| 26.00 | Tomotake et al. [24] | |||
| 28.32 | Kumar et al. [5] | |||
| 28.32 | Zhou et al. [17] | |||
| C18:2n-6 (linoleic) | 7.30 | 8.50 | Nakasone and Ito [43] | |
| 6.30 | 7.70 | Kotake-Nara et al. [39] | ||
| 5.81–8.57 | Pereira et al. [31] | |||
| 7.30 | Tomotake et al. [24] | |||
| 3.88 | Kumar et al. [5] | |||
| 3.88 | Zhou et al. [17] | |||
| C18:3n-3 (alpha-linolenic) | 29.20 | 34.90 | Nakasone and Ito [43] | |
| 17–33.40 | Pereira et al. [39] | |||
| 36.00 | 40.70 | Kotake-Nara et al. [31] | ||
| 36.30 | Tomotake et al. [24] | |||
| 38.25 | Kumar et al. [5] | |||
| 38.25 | Zhou et al. [17] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hăbeanu, M.; Gheorghe, A.; Mihalcea, T. Nutritional Value of Silkworm Pupae (Bombyx mori) with Emphases on Fatty Acids Profile and Their Potential Applications for Humans and Animals. Insects 2023, 14, 254. https://doi.org/10.3390/insects14030254
Hăbeanu M, Gheorghe A, Mihalcea T. Nutritional Value of Silkworm Pupae (Bombyx mori) with Emphases on Fatty Acids Profile and Their Potential Applications for Humans and Animals. Insects. 2023; 14(3):254. https://doi.org/10.3390/insects14030254
Chicago/Turabian StyleHăbeanu, Mihaela, Anca Gheorghe, and Teodor Mihalcea. 2023. "Nutritional Value of Silkworm Pupae (Bombyx mori) with Emphases on Fatty Acids Profile and Their Potential Applications for Humans and Animals" Insects 14, no. 3: 254. https://doi.org/10.3390/insects14030254