
Temperature-Dependent Life Table Parameters of Brown Marmorated Stink Bug, Halyomorpha halys (Stål) (Hemiptera: Pentatomidae) in the United States

 , , ,
, , , 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Collection and Rearing
2.1.1. Halyomorpha halys Oregon Population Rearing
2.1.2. Halyomorpha halys New Jersey Population Rearing
2.2. Survival at Temperatures below Developmental Thresholds
2.3. Statistical Analysis
2.3.1. Temperature-Related Daily Survival and Reproduction
2.3.2. Physiological Age-Specific Survival and Maternity
3. Results
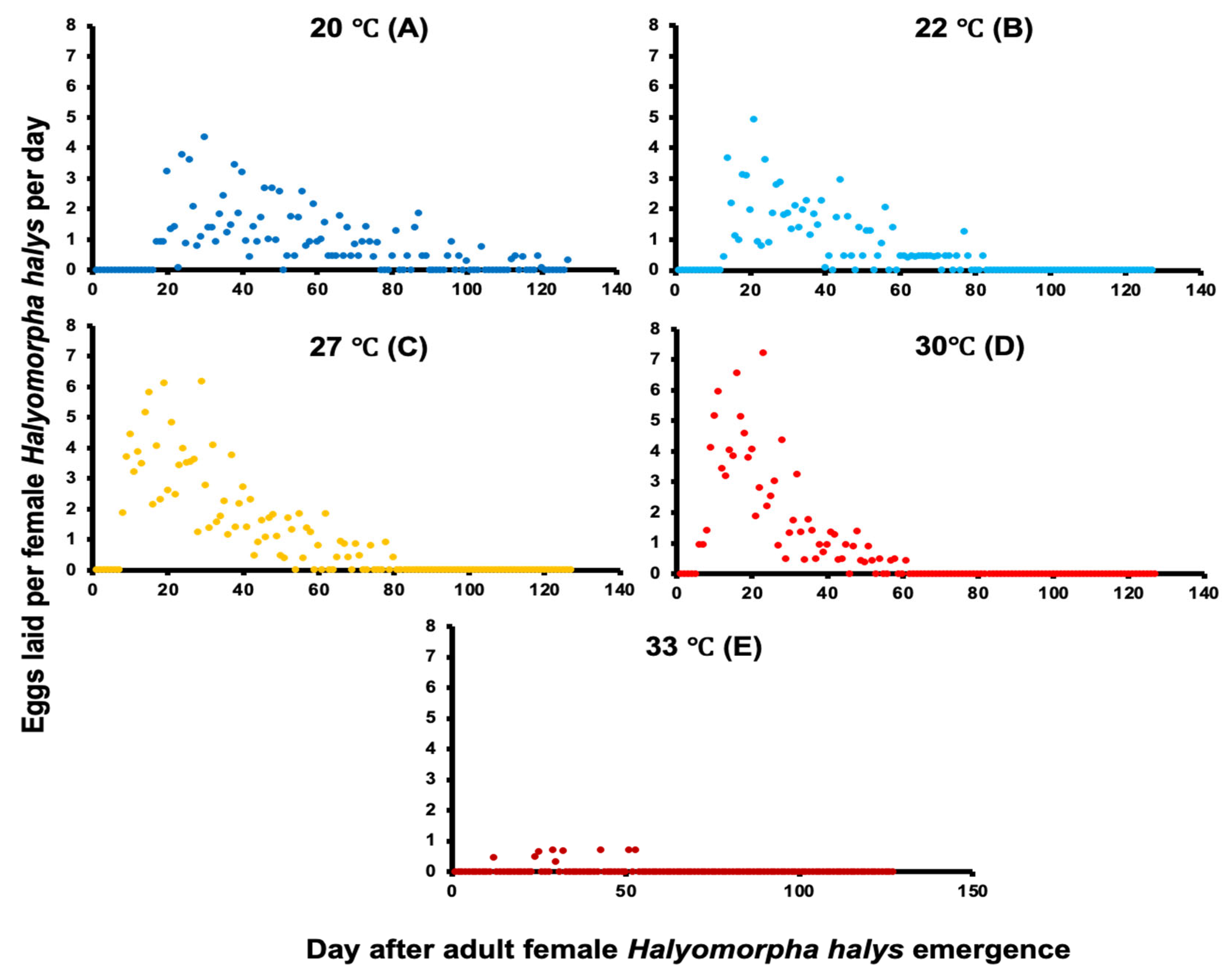
3.1. Life Span, Daily Mortality, Survival, and Oviposition
3.1.1. Halyomorpha halys Oregon Population
3.1.2. Halyomorpha halys New Jersey Population
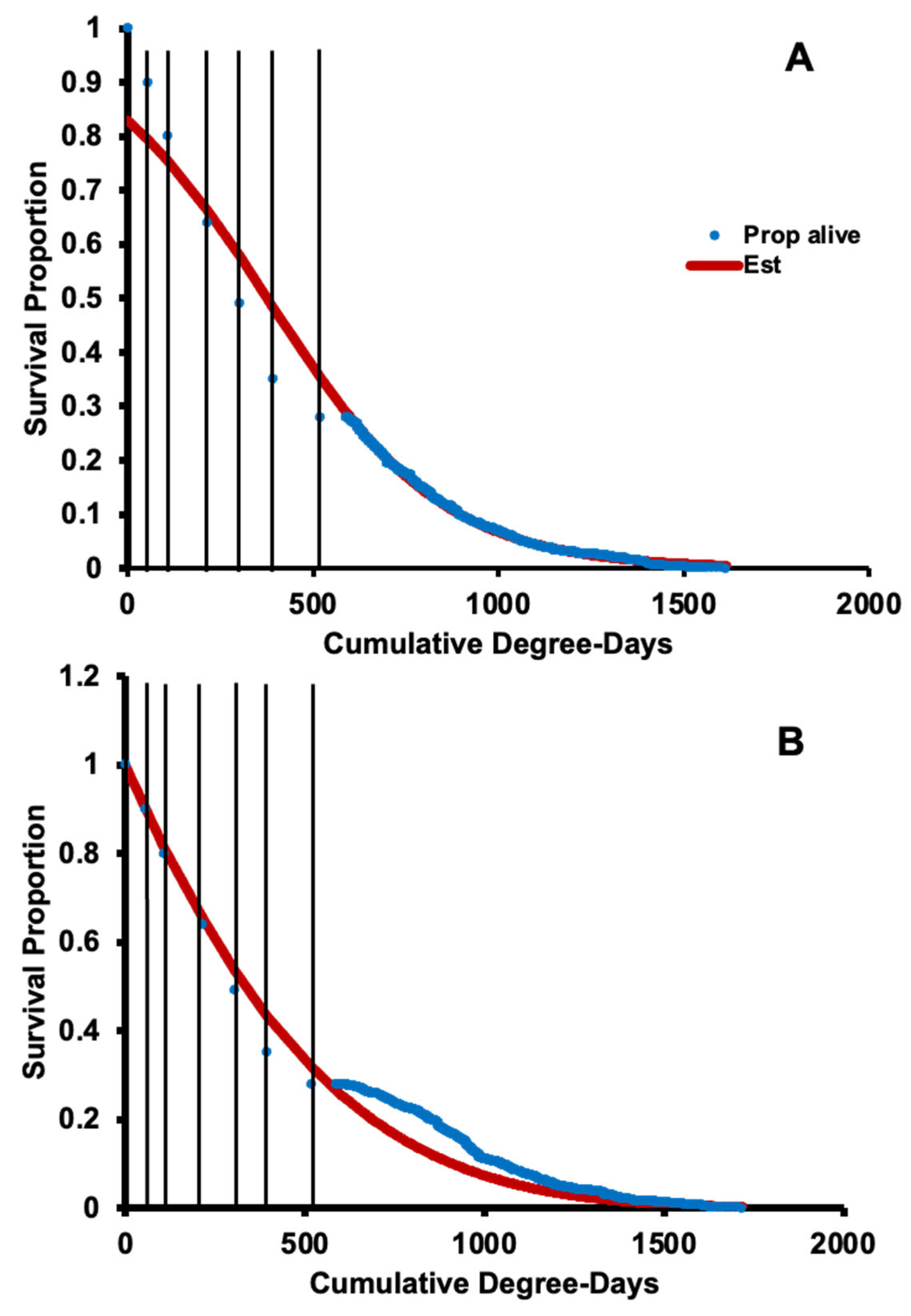
3.2. Survival, Oviposition, and Fecundity Fitting
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Wermelinger, B.; Wynige, D.; Forster, B. First records of an invasive bug in Europe: Halyomorpha halys Stål (Heteroptera: Pentatomidae), a new pest on woody ornamentals and fruit trees? Mitt. Schweiz. Entomol. Ges. 2008, 81, 1–8. [Google Scholar]
- Lee, D.-H.; Short, B.D.; Joseph, S.V.; Bergh, J.C.; Leskey, T.C. Review of the biology, ecology, and management of Halyomorpha halys (Hemiptera: Pentatomidae) in China, Japan, and the Republic of Korea. Environ. Entomol. 2013, 42, 627–641. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.R.; Hesler, S.P.; Loeb, G.M. Potential impact of Halyomorpha halys (Hemiptera: Pentatomidae) on grape production in the Finger Lakes region of New York. J. Entomol. Sci. 2014, 49, 290–303. [Google Scholar] [CrossRef]
- Nielsen, A.L.; Hamilton, G.C.; Matadha, D. Developmental rate estimation and life table analysis for Halyomorpha halys (Hemiptera: Pentatomidae). Environ. Entomol. 2008, 37, 348–355. [Google Scholar] [CrossRef] [PubMed]
- Bergmann, E.J.; Venugopal, P.D.; Martinson, H.M.; Raupp, M.J.; Shrewsbury, P.M. Host plant use by the invasive Halyomorpha halys (Stål) on Woody Ornamental Trees and Shrubs. PLoS ONE 2016, 11, e0149975. [Google Scholar] [CrossRef]
- Peiffer, M.; Felton, G.W. Insights into the saliva of the brown marmorated stink bug Halyomorpha halys (Hemiptera: Pentatomidae). PLoS ONE 2014, 9, e88483. [Google Scholar] [CrossRef]
- Niva, C.C.; Takeda, M. Effects of photoperiod, temperature and melatonin on nymphal development, polyphenism and reproduction in Halyomorpha halys (Heteroptera: Pentatomidae). Zool. Sci. 2003, 20, 963–970. [Google Scholar] [CrossRef]
- Rice, K.B.; Bergh, C.J.; Bergmann, E.J.; Biddinger, D.J.; Dieckhoff, C.; Dively, G.; Fraser, H.; Gariepy, T.; Hamilton, G.; Haye, T.; et al. Biology, ecology, and management of brown marmorated stink bug (Hemiptera: Pentatomidae). J. Integr. Pest Manag. 2014, 5, A1–A13. [Google Scholar] [CrossRef]
- Nielsen, A.L.; Chen, S.; Fleischer, S.J. Coupling developmental physiology, photoperiod, and temperature to model phenology and dynamics of an invasive heteropteran, Halyomorpha halys. Front. Physiol. 2016, 7, 165. [Google Scholar] [CrossRef]
- Haye, T.; Abdallah, S.; Gariepy, T.; Wyniger, D. Phenology, life table analysis and temperature requirements of the invasive brown marmorated stink bug, Halyomorpha halys, in Europe. J. Pest Sci. 2014, 87, 407–418. [Google Scholar] [CrossRef]
- Baek, S.; Hwang, A.; Kim, H.; Lee, H.; Lee, J.-H. Temperature-dependent development and oviposition models of Halyomorpha halys (Hemiptera: Pentatomidae). J. Asia-Pac. Entomol. 2017, 20, 367–375. [Google Scholar] [CrossRef]
- Govindan, B.N.; Hutchison, W.D. Influence of temperature on age-stage, two-sex life tables for a Minnesota-acclimated population of the brown marmorated stink bug (Halyomorpha halys). Insects 2020, 11, 108. [Google Scholar] [CrossRef]
- Kamiyama, M.T.; Matsuura, K.; Yoshimura, T.; Yang, C.-C.S. Improving invasive species management using predictive phenology models: An example from brown marmorated stink bug (Halyomorpha halys) in Japan. Pest Manag. Sci. 2021, 77, 5489–5497. [Google Scholar] [CrossRef]
- Ogburn, E.C.; Ohmen, T.M.; Huseth, A.S.; Reisig, D.D.; Kennedy, G.G.; Walgenbach, J.F. Temperature-driven differences in phenology and habitat suitability for brown marmorated stink bug, Halyomorpha halys, in two ecoregions of North Carolina. J. Pest Sci. 2022, 96, 373–387. [Google Scholar] [CrossRef]
- Kawada, H.; Kitamura, C. The Reproductive behavior of the brown marmorated stink bug, Halyomorpha mista (Heteroptera: Pentatomidae) observation of mating behavior and multiple copulation. Appl. Entomol. Zool. 1983, 18, 234–242. [Google Scholar] [CrossRef]
- Medal, J.; Smith, T.; Cruz, A.S. Biology of the brown marmorated stink bug Halyomorpha halys (Heteroptera: Pentatomidae) in the laboratory. Fla. Entomol. 2013, 96, 1209–1212. [Google Scholar] [CrossRef]
- Fand, B.B.; Sul, N.T.; Bal, S.K.; Minhas, P.S. Temperature impacts the development and survival of common cutworm (Spodoptera litura): Simulation and visualization of potential population growth in India under warmer temperatures through life cycle modelling and spatial mapping. PLoS ONE 2015, 10, e0124682. [Google Scholar] [CrossRef]
- Kiritani, K.; Hôkyo, N.; Kimura, K. Survival rate and reproductivity of the adult southern green stink bug, Nezara viridula. Jpn. J. Appl. Entomol. Zool. 1963, 7, 113–124. [Google Scholar] [CrossRef]
- Carey, J.R. Applied Demography for Biologists with Special Emphasis on Insects; Oxford University Press: New York, NY, USA, 1993. [Google Scholar]
- Price, P.W. Insect Ecology, 3rd ed.; John Wiley & Sons, Inc.: New York, NY, USA, 1997. [Google Scholar]
- Briere, J.-F.; Pracros, P.; Le Roux, A.-Y.; Pierre, J.-S. A novel rate model of temperature-dependent development for arthropods. Environ. Entomol. 1999, 28, 22–29. [Google Scholar] [CrossRef]
- Zalom, F.G.; Goodell, P.B.; Wilson, L.T.; Barnett, W.W.; Bentley, W.J. Degree-Days: The calculation and use of heat units in pest management. Univ. Calif. Div. Agric. Nat. Resour. Leafl. 1983, 21373. Available online: https://www.researchgate.net/publication/262337197_Degree-Days_the_Calculation_and_Use_of_Heat_Units_in_Pest_Management_University_of_California_Division_of_Agriculture_and_Natural_Resources_Leaflet_21373 (accessed on 30 December 2022).
- Jackson, C. Flexsurv: A platform for parametric survival modeling in R. J. Stat. Softw. 2016, 70, i08. [Google Scholar] [CrossRef] [PubMed]
- Rosen, S.J.; Cambridge, J.E.; Hamilton, G.C. Rearing protocol and density trials of the brown marmorated stink bug (Hemiptera: Pentatomidae) in the laboratory. Fla. Entomol. 2016, 99, 808–810. [Google Scholar] [CrossRef]
- Gariepy, T.D.; Haye, T.; Fraser, H.; Zhang, J. Occurrence, genetic diversity, and potential pathways of entry of Halyomorpha halys in newly invaded areas of Canada and Switzerland. J. Pest Sci. 2014, 87, 17–28. [Google Scholar] [CrossRef]
- Cesari, M.; Maistrello, L.; Piemontese, L.; Bonini, R.; Dioli, P.; Lee, W.; Park, C.-G.; Partsinevelos, G.K.; Rebecchi, L.; Guidetti, R. Genetic diversity of the brown marmorated stink bug Halyomorpha halys in the invaded territories of Europe and its patterns of diffusion in Italy. Biol. Invasions 2018, 20, 1073–1092. [Google Scholar] [CrossRef]
- Illán, J.G.; Zhu, G.; Walgenbach, J.F.; Acebes-Doria, A.; Agnello, A.M.; Alston, D.G.; Andrews, H.; Beers, E.H.; Bergh, J.C.; Bessin, R.T.; et al. Evaluating invasion risk and population dynamics of the brown marmorated stink bug across the contiguous United States. Pest Manag. Sci. 2022, 78, 4929–4938. [Google Scholar] [CrossRef]
- Lowenstein, D.M.; Walton, V.M. Halyomorpha halys (Hemiptera: Pentatomidae) winter survival, feeding activity, and reproduction rates based on episodic cold shock and winter temperature regimes. J. Econ. Entomol. 2018, 111, 1210–1218. [Google Scholar] [CrossRef]
- Bergh, J.C.; Morrison, W.R., III; Joseph, S.V.; Leskey, T.C. Characterizing spring emergence of adult Halyomorpha halys using experimental overwintering shelters and commercial pheromone traps. Entomol. Exp. Appl. 2017, 162, 336–345. [Google Scholar] [CrossRef]
- Kereselidze, M.; Pilarska, D.; Linde, A.; Sanscrainte, N.D.; Hajek, A.E. Nosema maddoxi infecting the brown marmorated stink bug, Halyomorpha halys(Stål) (Hemiptera: Pentatomidae), in the Republic of Georgia. Biocontrol. Sci. Technol. 2020, 30, 1083–1089. [Google Scholar] [CrossRef]
- Valentin, R.E.; Nielsen, A.L.; Wiman, N.G.; Lee, D.-H.; Fonseca, D.M. Global invasion network of the brown marmorated stink bug, Halyomorpha halys. Sci. Rep. 2017, 7, 9866. [Google Scholar] [CrossRef]
- McCalla, K.A.; Keçeci, M.; Milosavljević, I.; Ratkowsky, D.A.; Hoddle, M.S. The influence of temperature variation on life history parameters and thermal performance curves of Tamarixia radiata (Hymenoptera: Eulophidae), a parasitoid of the asian citrus psyllid (Hemiptera: Liviidae). J. Econ. Entomol. 2019, 112, 1560–1574. [Google Scholar] [CrossRef]
- Milosavljević, I.; McCalla, K.A.; Ratkowsky, D.A.; Hoddle, M.S. Effects of constant and fluctuating temperatures on development rates and longevity of Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae). J. Econ. Entomol. 2019, 112, 1062–1072. [Google Scholar] [CrossRef]
- Pfab, F.; Stacconi, M.V.R.; Anfora, G.; Grassi, A.; Walton, V.; Pugliese, A. optimized timing of parasitoid release: A mathematical model for biological control of Drosophila suzukii. Theor. Ecol. 2018, 11, 489–501. [Google Scholar] [CrossRef]
- Al Shidi, R.H.; Kumar, L.; Al-Khatri, S.A.H.; Alaufi, M.S.; Albahri, M.M. Does solar radiation affect the distribution of dubas bug (Ommatissus lybicus de Bergevin) infestation. Agriculture 2018, 8, 107. [Google Scholar] [CrossRef]
- Amore, V.; Hernández, M.I.M.; Carrascal, L.M.; Lobo, J.M. Exoskeleton may influence the internal body temperatures of neotropical dung beetles (Coleoptera: Scarabaeinae). PeerJ 2017, 5, e3349. [Google Scholar] [CrossRef]
- Havko, N.E.; Das, M.R.; McClain, A.M.; Kapali, G.; Sharkey, T.D.; Howe, G.A. Insect herbivory antagonizes leaf cooling responses to elevated temperature in tomato. Proc. Natl. Acad. Sci. USA 2020, 117, 2211–2217. [Google Scholar] [CrossRef]
- Joseph, S.V.; Stallings, J.W.; Leskey, T.C.; Krawczyk, G.; Polk, D.; Butler, B.; Bergh, J.C. Spatial distribution of brown marmorated stink bug (Hemiptera: Pentatomidae) injury at harvest in Mid-Atlantic apple orchards. J. Econ. Entomol. 2014, 107, 1839–1848. [Google Scholar] [CrossRef]
- Acebes-Doria, A.L.; Agnello, A.M.; Alston, D.G.; Andrews, H.; Beers, E.H.; Bergh, J.C.; Bessin, R.; Blaauw, B.R.; Buntin, G.D.; Burkness, E.C.; et al. Season-long monitoring of the brown marmorated stink bug (Hemiptera: Pentatomidae) throughout the United States using commercially available traps and lures. J. Econ. Entomol. 2020, 113, 159–171. [Google Scholar] [CrossRef]
- Zhu, G.; Gutierrez Illan, J.; Crowder, D.W. The use of insect life tables in optimizing invasive pest distributional models. Ecography 2021, 44, 1501–1510. [Google Scholar] [CrossRef]
- Otieno, M.H.J.; Ayieko, M.A.; Niassy, S.; Salifu, D.; Abdelmutalab, A.G.A.; Fathiya, K.M.; Subramanian, S.; Fiaboe, K.K.M.; Roos, N.; Ekesi, S.; et al. Integrating temperature-dependent life table data into insect life cycle model for predicting the potential distribution of Scapsipedus icipe Hugel & Tanga. PLoS ONE 2019, 14, e0222941. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Temperature (°C) | Egg to Adult (Days) | Adult to Mortality (Days ± SD) | EFD (Mean Eggs Laid per Female per Day) | EF (Mean Eggs per Female) | N |
|---|---|---|---|---|---|
| Oregon | |||||
| 15 | - | 55.5 ± 47.0 | - | - | 25 |
| 18 | - | 75.1 ± 57.3 | 0.10 | 7.8 | 44 |
| 22 | - | 43.8 ± 32.9 | 0.77 | 33.9 | 42 |
| 25 | - | 28.3 ± 17.6 | 1.7 | 48.6 | 50 |
| 27 | - | 22.8 ± 20.5 | 0.93 | 21.1 | 44 |
| 30 | - | 13.8 ± 10.7 | 0.87 | 11.9 | 50 |
| 32 | - | 15.7 ± 12.8 | 0.13 | 2.1 | 44 |
| New Jersey | |||||
| 17 | 121.5 ± 0.5 ¢ | - | - | - | - |
| 20 | 81.2 ± 0.8 ¢ | 52.7 ± 28.4 | 1.0 | 54.4 | 61 |
| 22 | - | 42.2 ± 23.4 | 1.1 | 48.5 | 60 |
| 25 | 44.9 ± 0.8 ¢ | 47.97 | - | 212.3 | 28 |
| 27 | 35.8 ± 0.5 ¢ | 32.8 ± 21.3 | 2.2 | 72.3 | 59 |
| 30 | 33.4 ± 0.5 ¢ | 31.1 ± 15.5 | 2.4 | 73.6 | 60 |
| 33 | 37.8 ± 0.9 ¢ | 24.9 ± 13.6 | 0.04 | 1.1 | 7 |
| Population | Model Fit | R2 | p |
|---|---|---|---|
| Oregon | y = 0.00000156 (6.513) (42.0577) − x)(1/0.253) | 0.92 | 0.002 |
| New Jersey | y = 1/(3.14 × (4.61591) × (1 + ((x − (9.30887))/(4.61591))2)) (538.946) | 0.79 | 0.001 |
| Temperature (°C) | Ro | T | rm |
|---|---|---|---|
| 18 | 7.6 † | 142 † | 0.02 † |
| 20 | 68.8 §, 54.4 ¥ | 145 §, 140 ¥ | 0.028 §, 0.028 ¥ |
| 22 | 48.5 ¥, 33 † | 111 ¥, 123.1† | 0.06 ¥, 0.04 † |
| 23 | 128 § | 79 § | 0.06 § |
| 25 | 49 †, 60 ¢ | 103.7 †, 60 ¢ | 0.065 †, 0.07 ¢ |
| 27 | 151 §, 72.3 ¥, 21.1 † | 56.8 §, 69 ¥, 72.1† | 0.087 §, 0.063 ¥, 0.074 † |
| 30 | 58.12 §, 74.6 ¥, 11† | 50.94 §, 33.1 ¥, 37.7† | 0.079 §, 0.053 ¥, 0.02 † |
| 32 | 2 † | 48.5 † | 0.001 † |
| 33 | 6.8 §, 2.05 ¥ | 51.2 §, 44 ¥ | 0.0339 §, 0.016 ¥ |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mermer, S.; Maslen, E.A.; Dalton, D.T.; Nielsen, A.L.; Rucker, A.; Lowenstein, D.; Wiman, N.; Bhattarai, M.; Soohoo-Hui, A.; Harris, E.T.; et al. Temperature-Dependent Life Table Parameters of Brown Marmorated Stink Bug, Halyomorpha halys (Stål) (Hemiptera: Pentatomidae) in the United States. Insects 2023, 14, 248. https://doi.org/10.3390/insects14030248
Mermer S, Maslen EA, Dalton DT, Nielsen AL, Rucker A, Lowenstein D, Wiman N, Bhattarai M, Soohoo-Hui A, Harris ET, et al. Temperature-Dependent Life Table Parameters of Brown Marmorated Stink Bug, Halyomorpha halys (Stål) (Hemiptera: Pentatomidae) in the United States. Insects. 2023; 14(3):248. https://doi.org/10.3390/insects14030248
Chicago/Turabian StyleMermer, Serhan, Erika A. Maslen, Daniel T. Dalton, Anne L. Nielsen, Ann Rucker, David Lowenstein, Nik Wiman, Mukesh Bhattarai, Alexander Soohoo-Hui, Edwin T. Harris, and et al. 2023. "Temperature-Dependent Life Table Parameters of Brown Marmorated Stink Bug, Halyomorpha halys (Stål) (Hemiptera: Pentatomidae) in the United States" Insects 14, no. 3: 248. https://doi.org/10.3390/insects14030248