
Fitness Costs and Incomplete Resistance Associated with Delayed Evolution of Practical Resistance to Bt Crops

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Literature Review
2.2. Fitness Costs
2.3. Incomplete Resistance
2.4. Computer Simulations of Evolution of Resistance
3. Results
3.1. Comparison of Costs between Cases with and without Practical Resistance
3.1.1. Costs in Resistant Strains
3.1.2. Costs in F1 Progeny
3.1.3. Frequency and Dominance of Costs in Unrelated vs. Related Strains
3.1.4. Hybrid Vigor and Higher Fitness in Resistant Than Susceptible Strains in Unrelated vs. Related Strains
3.2. Comparison of Incomplete Resistance between Cases with and without Practical Resistance
3.3. Modeling the Effects of Costs and Incomplete Resistance on Evolution of Practical Resistance
4. Conclusion and Future Prospects
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Shelton, A.M.; Hossain, M.J.; Paranjape, V.; Azad, A.K.; Rahman, M.L.; Khan, A.S.M.M.R.; Prodhan, M.Z.H.; Rashid, M.A.; Majumder, R.; Hossain, M.A.; et al. Bt eggplant project in Bangladesh: History, present status, and future direction. Front. Bioeng. Biotechnol. 2018, 6, 106. [Google Scholar] [CrossRef] [Green Version]
- Barrero, J.M.; MacKenzie, D.J.; Goodman, R.E.; Higgins, T.J.V. A biotech shield for cowpea against the major insect pests. Open Access News. 2021. Available online: https://www.openaccessgovernment.org/major-insect-pests/111590/ (accessed on 12 February 2023).
- ISAAA. Global Status of Commercialized Biotech/GM Crops in 2019: Biotech Crops Drive Socio-Economic Development and Sustainable Environment in the New Frontier. ISAAA Brief No. 55; ISAAA: Ithaca, NY, USA, 2019. [Google Scholar]
- Van den Berg, J.; Prasanna, B.M.; Midega, C.A.O.; Ronald, P.C.; Carrière, Y.; Tabashnik, B.E. Managing fall armyworm in Africa: Can Bt maize sustainably improve control? J. Econ. Entomol. 2021, 114, 1934–1949. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, W.S.; Sakuno, C.I.R.; Miraldo, L.I.; Tavares, M.A.G.C.; Komada, K.M.A.; Teresani, D.; Santos, J.L.X.; Huang, F. Varied frequencies of resistance alleles to Cry1Ab and Cry1Ac among Brazilian populations of the sugarcane borer, Diatraea saccharalis (F.). Pest Manag. Sci. 2022, 12, 5150–5163. [Google Scholar] [CrossRef] [PubMed]
- Cattaneo, M.G.; Yafuso, C.; Schmidt, C.; Huang, C.; Rahman, M.; Olson, C.; Ellers-Kirk, C.; Orr, B.J.; Marsh, S.E.; Antilla, L.; et al. Farm-scale evaluation of transgenic cotton impacts on biodiversity, pesticide use, and yield. Proc. Natl. Acad. Sci. USA 2006, 103, 7571–7576. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bird, L.J. Pyrethroid and carbamate resistance in Australian Helicoverpa armigera (Lepidoptera: Noctuidae) from 2008 to 2015: What has changed since the introduction of Bt cotton? Bull. Entomol. Res. 2018, 108, 781–791. [Google Scholar] [CrossRef]
- Dively, G.P.; Venugopal, P.D.; Bean, D.; Whalen, J.; Holmstrom, K.; Kuhar, T.P.; Doughty, H.B.; Patton, T.; Cissel, W.; Hutchison, W.D. Regional pest suppression associated with widespread Bt maize adoption benefits vegetable growers. Proc. Natl. Acad. Sci. USA 2018, 115, 3320–3325. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Lu, Y.; van der Werf, W.; Huang, J.; Wu, F.; Zhou, K.; Deng, X.; Jiang, Y.; Wu, K.; Rosegrant, M.W. Multidecadal, county-level analysis of the effects of land use, Bt cotton, and weather on cotton pests in China. Proc. Natl. Acad. Sci. USA 2018, 115, E7700–E7709. [Google Scholar] [CrossRef] [Green Version]
- Carrière, Y.; Ellers-Kirk, C.; Sisterson, M.; Antilla, L.; Whitlow, M.; Dennehy, T.J.; Tabashnik, B.E. Long-term regional suppression of pink bollworm by Bacillus thuringiensis cotton. Proc. Natl. Acad. Sci. USA 2003, 100, 1519–1523. [Google Scholar] [CrossRef] [Green Version]
- Wu, K.M.; Lu, Y.H.; Feng, H.Q.; Jiang, Y.Y.; Zhao, J.Z. Suppression of cotton bollworm in multiple crops in China in areas with Bt toxin-containing cotton. Science 2008, 321, 1676–1678. [Google Scholar] [CrossRef] [Green Version]
- Hutchison, W.D.; Burkness, E.C.; Mitchell, P.D.; Moon, R.D.; Leslie, T.W.; Fleischer, S.J.; Abrahamson, M.; Hamilton, K.L.; Steffey, K.L.; Gray, M.E.; et al. Areawide suppression of European corn borer with Bt maize reaps savings to non-Bt maize growers. Science 2010, 330, 222–225. [Google Scholar] [CrossRef] [Green Version]
- Tabashnik, B.E.; Liesner, L.R.; Ellsworth, P.C.; Unnithan, G.C.; Fabrick, J.A.; Naranjo, S.E.; Li, X.; Dennehy, T.J.; Antilla, L.; Staten, R.T.; et al. Genetically engineered cotton synergizes eradication of the pink bollworm a century after its invasion of the United States. Proc. Natl. Acad. Sci. USA 2021, 118, e2019115118. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Fabrick, J.A.; Carrière, Y. Global patterns of insect resistance to transgenic Bt crops: The first 25 years. J. Econ. Entomol. 2023. [Google Scholar] [CrossRef] [PubMed]
- Carrière, Y.; Brown, Z.S.; Downes, S.J.; Gujar, G.; Epstein, G.; Omoto, C.; Storer, N.P.; Mota-Sanchez, D.; Søgaard Jørgensen, P.; Carroll, S.P. Governing evolution: A socio-ecological comparison of resistance management for insecticidal transgenic Bt crops among four countries. AMBIO 2020, 49, 1–16. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabashnik, B.E.; Carrière, Y. Global patterns of resistance to Bt crops highlighting pink bollworm in the United States, China, and India. J. Econ. Entomol. 2019, 112, 2513–2523. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Mota-Sanchez, D.; Whalon, M.E.; Hollingworth, R.M.; Carrière, Y. Defining terms for proactive management of resistance to Bt crops and pesticides. J. Econ. Entomol. 2014, 107, 496–507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tabashnik, B.E.; Gould, F.; Carrière, Y. Delaying evolution of resistance to transgenic crops by decreasing dominance and heritability. J. Evol. Biol. 2004, 17, 904–912. [Google Scholar] [CrossRef] [PubMed]
- Carrière, Y.; Crickmore, N.; Tabashnik, B.E. Optimizing pyramided transgenic Bt crops for sustainable pest management. Nat. Biotechnol. 2015, 33, 161–168. [Google Scholar] [CrossRef]
- Carrière, Y.; Fabrick, J.A.; Tabashnik, B.E. Can pyramids and seed mixtures delay resistance to Bt crops? Trends Biotechnol. 2016, 34, 291–302. [Google Scholar] [CrossRef] [Green Version]
- Comins, H.N. Tactics for resistance management using multiple pesticides. Agric. Ecosyst. Environ. 1986, 16, 129–148. [Google Scholar] [CrossRef]
- Roush, R.T. Two-toxin strategies for management of insecticidal transgenic crops: Can pyramiding succeed where pesticide mixtures have not? Phil. Trans. Roy. Soc. London B 1998, 353, 1777–1786. [Google Scholar] [CrossRef] [Green Version]
- Tabashnik, B.E.; Gassmann, A.J.; Crowder, D.W.; Carrière, Y. Insect resistance to Bt crops: Evidence versus theory. Nat. Biotechnol. 2008, 26, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Carrière, Y.; Crowder, D.W.; Tabashnik, B.E. Evolutionary ecology of adaptation to Bt crops. Evol. Appl. 2010, 3, 561–573. [Google Scholar] [CrossRef] [PubMed]
- Tabashnik, B.E.; Brévault, T.; Carrière, Y. Insect resistance to Bt crops: Lessons from the first billion acres. Nat. Biotechnol. 2013, 31, 510–520. [Google Scholar] [CrossRef]
- Huang, F. Dominance and fitness costs of insect resistance to genetically modified Bacillus thuringiensis crops. GM Crop. Food 2021, 12, 192–211. [Google Scholar] [CrossRef]
- Carrière, Y.; Tabashnik, B.E. Reversing insect adaptation to transgenic insecticidal plants. Proc. Roy. Soc. Lond. 2001, 268, 1475–1480. [Google Scholar] [CrossRef] [Green Version]
- Gassmann, A.J.; Carrière, Y.; Tabashnik, B.E. Fitness costs of insect resistance to Bacillus thuringiensis. Ann. Rev. Entomol. 2009, 54, 147–163. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Carrière, Y. Surge in insect resistance to transgenic crops and prospects for sustainability. Nat. Biotechnol. 2017, 35, 926–935. [Google Scholar] [CrossRef]
- Crowder, D.W.; Carrière, Y. Comparing the refuge strategy for managing the evolution of insect resistance under different reproductive strategies. J. Theor. Biol. 2009, 261, 423–430. [Google Scholar] [CrossRef]
- Tabashnik, B.E.; Dennehy, T.J.; Carrière, Y. Delayed resistance to transgenic cotton in pink bollworm. Proc. Natl. Acad. Sci. USA 2005, 43, 15389–15393. [Google Scholar] [CrossRef] [Green Version]
- Gould, F.; Cohen, M.B.; Bentur, J.S.; Kennedy, G.G.; Van Duyn, J. Impact of small fitness costs on pest adaptation to crop varieties with multiple toxins: A heuristic model. J. Econ. Entomol. 2006, 99, 2091–2099. [Google Scholar] [CrossRef] [PubMed]
- Alphey, N.; Coleman, P.G.; Bonsall, M.B.; Alphey, L. Proportions of different habitat types are critical to the fate of a resistance allele. Theor. Ecol. 2008, 1, 103–115. [Google Scholar] [CrossRef]
- Wan, P.; Xu, D.; Cong, S.-B.; Jiang, Y.-Y.; Huang, Y.; Wang, J.-T.; Wu, H.-H.; Wang, L.; Wu, K.-M.; Carrière, Y.; et al. Hybridizing transgenic Bt plants with non-Bt plants counters insect resistance. Proc. Natl. Acad. Sci. USA 2017, 114, 5413–5418. [Google Scholar] [CrossRef] [Green Version]
- Santos-Amaya, O.F.; Tavares, C.S.; Rodrigues, J.V.C.; Campos, S.O.; Guedes, R.N.C.; Alves, A.P.; Pereira, E.J.G. Fitness costs and stability of Cry1Fa resistance in Brazilian populations of Spodoptera frugiperda. Pest. Manag. Sci. 2017, 73, 35–43. [Google Scholar] [CrossRef] [Green Version]
- Bird, L.J.; Akhurst, J.A. Relative fitness of Cry1A-resistant and -susceptible Helicoverpa armigera (Lepidoptera: Noctuidae) on conventional and transgenic cotton. J. Econ. Entomol. 2004, 97, 1699–1709. [Google Scholar] [CrossRef]
- Bird, L.J.; Akhurst, J.A. Fitness of Cry1A-resistant and -susceptible Helicoverpa armigera (Lepidoptera: Noctuidae) on transgenic cotton with reduced levels of Cry1Ac. J. Econ. Entomol. 2005, 98, 1311–1319. [Google Scholar] [CrossRef]
- Chen, X.; Head, G.P.; Price, P.; Kerns, D.L.; Rice, M.E.; Huang, F.; Gilreath, R.T.; Yang, F. Fitness costs of Vip3A resistance in Spodoptera frugiperda on different hosts. Pest Manag. Sci. 2019, 75, 1074–1080. [Google Scholar] [CrossRef]
- Gulzar, A.; Pickett, B.; Sayyed, A.H.; Wright, D.J. Effect of temperature on the fitness of a Vip3A resistant population of Heliothis virescens (Lepidoptera: Noctuidae). J. Econ. Entomol. 2012, 105, 964–970. [Google Scholar] [CrossRef]
- Shrestha, R.B.; Gassmann, A.J. Inheritance and fitness costs of Cry3Bb1 resistance in diapausing field strains of western corn rootworm (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2020, 113, 2873–2882. [Google Scholar] [CrossRef]
- Ghimire, M.N.; Huang, F.; Leonard, R.; Head, G.P.; Yang, Y. Susceptibility of Cry1Ab-susceptible and -resistant sugarcane borer to transgenic corn plants containing single or pyramided Bacillus thuringiensis genes. Crop. Protect. 2011, 30, 74e81. [Google Scholar] [CrossRef]
- Hoffmann, M.; French, B.W.; Hellmich, R.L.; Lauter, N.; Gassmann, A.J. Fitness costs of resistance to Cry3Bb1 maize by western corn rootworm. J. Appl. Entomol. 2015, 139, 403–415. [Google Scholar] [CrossRef] [Green Version]
- Carrière, Y.; Degain, B.; Unnithan, G.C.; Harpold, V.S.; Li, X.; Tabashnik, B.E. Seasonal declines in Cry1Ac and Cry2Ab concentration in maturing cotton favor faster evolution of resistance to pyramided Bt cotton in Helicoverpazea. J. Econ. Entomol. 2019, 112, 2907–2914. [Google Scholar] [CrossRef] [PubMed]
- Petzold-Maxwell, J.L.; Siegfried, B.D.; Hellmich, R.L.; Abel, C.A.; Coates, B.S.; Spencer, T.A.; Horikoshi, R.J.; Gassmann, A.J. Fitness costs associated with Cry1F resistance in the European corn borer. J. Appl. Entomol. 2017, 141, 67–79. [Google Scholar] [CrossRef] [Green Version]
- Charlesworth, D.; Willis, J.H. The genetics of inbreeding depression. Nat. Rev. Genet. 2009, 10, 783–796. [Google Scholar] [CrossRef] [PubMed]
- Carrière, Y.; Ellers Kirk, C.; Biggs, R.W.; Nyboer, M.E.; Unnithan, G.C.; Dennehy, T.J.; Tabashnik, B.E. Cadherin-based resistance to Bt cotton in hybrid strains of pink bollworm: Fitness costs and incomplete resistance. J. Econ. Entomol. 2006, 99, 1925–1935. [Google Scholar] [CrossRef]
- Brévault, T.; Heuberger, S.; Zhang, M.; Ellers-Kirk, C.; Ni, X.; Masson, L.; Li, X.; Tabashnik, B.E.; Carrière, Y. Potential shortfall of pyramided Bt cotton for resistance management. Proc. Natl. Acad. Sci. USA 2013, 110, 5806–5811. [Google Scholar] [CrossRef] [Green Version]
- Jin, L.; Wang, J.; Guan, F.; Zhang, J.; Yu, S.; Liu, S.; Xue, Y.; Lia, L.; Wu, S.; Wang, X.; et al. Dominant point mutation in a tetraspanin gene associated with field-evolved resistance of cotton bollworm to transgenic Bt cotton. Proc. Natl. Acad. Sci. USA 2018, 115, 11760–11765. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.; Chen, W.; Song, X.; Ma, X.; Cotto-Rivera, R.O.; Kain, W.; Chu, H.; Chen, Y.-R.; Fei, Z.; Wang, P. Mutation of ABC transporter ABCA2 confers resistance to Bt toxin Cry2Ab in Trichoplusiani. Insect Biochem. Mol. Biol. 2019, 112, 103209. [Google Scholar] [CrossRef]
- Banerjee, R.; De Bortoli, C.P.; Huang, F.; Lamour, K.; Meagher, R.; Buntin, D.; Ni, X.; Reay-Jones, F.P.F.; Stewart, S.; Jurat-Fuentes, J.L. Large genomic deletion linked to field-evolved resistance to Cry1Fcorn in fall armyworm (Spodoptera frugiperda) from Florida. Sci. Rep. 2022, 12, 13580. [Google Scholar] [CrossRef]
- Fabrick, J.A.; Li, X.; Carrière, Y.; Tabashnik, B.E. Molecular genetic basis of lab- and field-selected Bt resistance in pink bollworm. Insects 2023, 14, 201. [Google Scholar] [CrossRef]
- Liu, Y.B.; Tabashnik, B.E. Inheritance of resistance to Bacillus thuringiensis toxin Cry1C in the diamondback moth. Appl. Environ. Microbiol. 1997, 63, 2218–2223. [Google Scholar] [CrossRef] [Green Version]
- Mahon, R.J.; Olsen, K.M. Limited survival of a Cry2Ab-resistant strain of Helicoverpa armigera (Lepidoptera: Noctuidae) on Bollgard II. J. Econ. Entomol. 2009, 102, 708–716. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Leonard, B.R.; Chen, M.; Clark, T.; Anilkumar, K.; Huang, F. Fitness costs and stability of Cry1Ab resistance in sugarcane borer, Diatraea saccharalis (F.). J. Invertebr. Pathol. 2014, 117, 26–32. [Google Scholar] [CrossRef] [PubMed]
- Dangal, V.; Huang, F. Fitness costs of Cry1F resistance in two populations of fall armyworm, Spodoptera frugiperda (J.E. Smith), collected from Puerto Rico and Florida. J. Invertebr. Pathol. 2015, 127, 81–86. [Google Scholar] [CrossRef]
- Jurat-Fuentes, J.L.; Heckel, D.G.; Ferré, J. Mechanisms of resistance to insecticidal proteins from Bacillus thuringiensis. Annu. Rev. Entomol. 2021, 66, 121–140. [Google Scholar] [CrossRef]
- Gassmann, A.J.; Petzold-Maxwell, J.L.; Clifton, E.H.; Dunbar, M.W.; Hoffmann, A.M.; Ingber, D.A.; Keweshan, R.S. Field-evolved resistance by western corn rootworm to multiple Bacillus thuringiensis toxins in transgenic maize. Proc. Natl. Acad. Sci. USA 2014, 111, 5141–5146. [Google Scholar] [CrossRef] [Green Version]
- Cao, G.; Feng, H.; Guo, F.; Wu, K.; Li, X.; Liang, G.; Desneux, N. Quantitative analysis of fitness costs associated with the development of resistance to the Bt toxin Cry1Ac in Helicoverpa armigera. Sci. Rep. 2014, 4, 5629. [Google Scholar] [CrossRef] [Green Version]
- Carrière, Y.; Onstad, D.W. The role of landscapes in insect resistance management. In Insect Resistance Management: Biology, Economics and Prediction, 3rd ed.; Onstad, D.W., Knolhoff, L.M., Eds.; Academic Press: London, UK, 2023; pp. 329–380. [Google Scholar]
- Fitt, G.; Andow, D.A.; Carrière, Y.; Moar, W.A.; Schuler, T.; Omoto, C.; Kanya, J.; Okech, M.; Arama, P.; Maniania, N.K. Resistance risks and management associated with Bt maize in Kenya. In Environmental Risk Assessment of Genetically Modified Organisms, Volume 1: A Case Study of Bt Maize in Kenya; Hilbeck, A., Andow, D., Eds.; CABI International: Wallingford, UK, 2004; pp. 209–250. [Google Scholar]
- Acharya, B.; Head, G.P.; Price, P.A.; Huang, F. Fitness costs and inheritance of Cry2Ab2 resistance in Spodoptera frugiperda (J.E. Smith). J. Invertebr. Pathol. 2017, 149, 8–14. [Google Scholar] [CrossRef]
- Akhurst, R.J.; James, W.; Bird, L.J.; Beard, C. Resistance to the Cry1Ac-Endotoxin of Bacillus thuringiensis in the cotton bollworm, Helicoverpa armigera (Lepidoptera: Noctuidae). J. Econ. Entomol. 2003, 96, 1290–1299. [Google Scholar] [CrossRef]
- Bernardi, O.; Bernardi, D.; Horikoshi, R.J.; Okuma, D.M.; Miraldo, L.L.; Fatoretto, J.; Medeiros, F.C.L.; Burd, T.; Omoto, C. Selection and characterization of resistance to the Vip3Aa20 protein from Bacillus thuringiensis in Spodoptera frugiperda. Pest. Manag. Sci. 2016, 72, 1794–1802. [Google Scholar] [CrossRef]
- Carrière, Y.; Ellers-Kirk, C.; Liu, Y.-B.; Sims, M.A.; Patin, A.L.; Dennehy, T.J.; Tabashnik, B.E. Fitness costs and maternal effects associated with resistance to transgenic cotton in the pink bollworm (Lepidoptera: Gelechiidae). J. Econ. Entomol. 2001, 94, 1571–1576. [Google Scholar] [CrossRef]
- Carrière, Y.; Ellers-Kirk, C.; Biggs, R.; Higginson, D.M.; Dennehy, T.J.; Tabashnik, B.E. Effects of gossypol on fitness costs associated with resistance to Bt cotton in pink bollworm. J. Econ. Entomol. 2004, 97, 1710–1718. [Google Scholar] [CrossRef] [PubMed]
- Carrière, Y.; Ellers-Kirk, C.; Biggs, R.; Degain, B.; Holley, D.; Yafuso, C.; Evans, P.; Dennehy, T.J.; Tabashnik, B.E. Effects of cotton cultivar on fitness costs associated with resistance of pink bollworm (Lepidoptera: Gelechiidae) to Bt cotton. J. Econ. Entomol. 2005, 98, 947–954. [Google Scholar] [CrossRef] [PubMed]
- Carrière, Y.; Showalter, A.M.; Fabrick, J.A.; Sollome, J.; Ellers-Kirk, C.; Tabashnik, B.E. Cadherin gene expression and effects of Bt resistance on sperm transfer in pink bollworm. J. Ins. Physiol. 2009, 55, 1058–1064. [Google Scholar] [CrossRef] [PubMed]
- Carrière, Y.; Degain, B.A.; Unnithan, G.C.; Harpold, V.S.; Heuberger, S.; Li, X.; Tabashnik, B.E. Effects of seasonal changes in cotton plants on the evolution of resistance to pyramided cotton producing the Bt toxins Cry1Ac and Cry1F in Helicoverpazea. Pest. Manag. Sci. 2018, 74, 627–637. [Google Scholar] [CrossRef] [PubMed]
- Carrière, Y.; Degain, B.A.; Tabashnik, B.E. Effects of gene flow between Bt and non-Bt plants in a seed mixture of Cry1A.105 + Cry2Ab corn on performance of corn earworm in Arizona. Pest. Manag. Sci. 2021, 77, 2106–2113. [Google Scholar] [CrossRef]
- Crespo, A.L.B.; Spenser, T.A.; Tan, S.Y.; Siegfried, B.D. Fitness Costs of Cry1Ab Resistance in a Field-Derived Strain of Ostrinia nubilalis (Lepidoptera: Crambidae). J. Econ. Entomol. 2010, 103, 1386–1393. [Google Scholar] [CrossRef]
- Crickmore, N.; Berry, S.; Panneerselvam, S.; Mishra, R.; Connor, T.R.; Bonning, B.C. A structure-based nomenclature for Bacillus thuringiensis and other bacteria-derived pesticidal proteins. J. Invertebr. Pathol. 2020, 6, 107438. [Google Scholar] [CrossRef]
- de Souza Ribas, N.; McNeil, J.N.; Araújo, H.D.; de Souza Ribas, B.; Lima, E. The Effect of resistance to Bt corn on the reproductive output of Spodoptera frugiperda (Lepidoptera: Noctuidae). Insects 2022, 13, 196. [Google Scholar] [CrossRef]
- Geisert, R.W.; Hibbard, B.E. Evaluation of potential fitness costs associated with eCry3.1Ab resistance in Diabrotica virgifera virgifera (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2016, 109, 1853–1858. [Google Scholar] [CrossRef] [Green Version]
- Higginson, D.M.; Morin, S.; Nyboer, M.E.; Biggs, R.W.; Tabashnik, B.E.; Carrière, Y. Evolutionary trade-offs of insect resistance to Bacillus thuringiensis crops: Fitness cost affecting paternity. Evolution 2005, 59, 915–920. [Google Scholar] [CrossRef]
- Horikoshi, R.J.; Bernardi, O.; Bernardi, D.; Okuma, D.M.; Farias, J.R.; Miraldo, L.L.; Amaral, F.S.A.; Omoto, C. Near-isogenic Cry1F-resistant strain of Spodoptera frugiperda (Lepidoptera: Noctuidae) to investigate fitness cost associated with resistance in Brazil. J. Econ. Entomol. 2016, 109, 854–859. [Google Scholar] [CrossRef] [PubMed]
- Huang, F.; Chen, M.; Gowda, A.; Clark, T.L.; McNulty, B.C.; Yang, F.; Niu, Y. Identification, inheritance, and fitness costs of Cry2Ab2 resistance in a field-derived population of sugarcane borer, Diatraea saccharalis (F.) (Lepidoptera: Crambidae). J. Invertebr. Pathol. 2015, 130, 116–123. [Google Scholar] [CrossRef] [PubMed]
- Ingber, D.A.; Gassmann, A.J. Inheritance and fitness costs of resistance to Cry3Bb1 corn by western corn rootworm (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2015, 108, 2421–2432. [Google Scholar] [CrossRef] [PubMed]
- Jackson, R.E.; Bradley, J.R.; Van Duyn, J.W. Performance of feral and Cry1Ac-selected Helicoverpa zea (Lepidoptera: Noctuidae) strains on transgenic cottons expressing one or two Bacillus thuringiensis ssp. Kurstaki proteins under greenhouse conditions. J. Entomol. Sci. 2004, 39, 46–55. [Google Scholar] [CrossRef]
- Jakka, S.R.K.; Knight, V.R.; Jurat-Fuentes, J.L. Fitness costs associated with field-evolved resistance to Bt maize in Spodoptera frugiperda (Lepidoptera: Noctuidae). J. Econ. Entomol. 2014, 107, 342–351. [Google Scholar] [CrossRef] [PubMed]
- Kruger, M.; Van Rensburg, J.B.J.; Van den Berg, J. No fitness costs associated with resistance of Busseola fusca (Lepidoptera: Noctuidae) to genetically modified Bt maize. Crop Protect. 2014, 55, 1–6. [Google Scholar] [CrossRef]
- Lefko, S.A.; Nowatzki, T.M.; Thompson, S.D.; Binning, R.R.; Pascual, M.A.; Peters, M.L.; Simbro, E.J.; Stanley, B.H. Characterizing laboratory colonies of western corn rootworm (Coleoptera: Chrysomelidae) selected for survival on maize containing event DAS-59122-7. J. Appl. Entomol. 2008, 132, 189–204. [Google Scholar] [CrossRef]
- Leite, N.A.; Mendes, S.M.; Santos-Amaya, O.F.; Santos, C.A.; Teixeira, T.P.M.; Guedes, R.N.C.; Pereira, E.J.G. Rapid selection and characterization of Cry1F resistance in a Brazilian strain of fall armyworm. Entomol. Exp. Appl. 2016, 158, 236–247. [Google Scholar] [CrossRef]
- Liang, G.-M.; Wu, K.-M.; Rector, B.; Guo, Y.-Y. Diapause, cold hardiness and flight ability of Cry1Ac-resistant and –susceptible strains of Helicoverpa armigera (Lepidoptera: Noctuidae). Eur. J. Entomol. 2007, 104, 699–704. [Google Scholar] [CrossRef] [Green Version]
- Liang, G.-M.; Wu, K.-M.; Yu, H.K.; Li, K.-K.; Feng, X.; Guo, Y.-Y. Changes of inheritance mode and fitness in Helicoverpa armigera (Hubner) (Lepidoptera: Noctuidae) along with its resistance evolution to Cry1Ac toxin. J. Invertebr. Pathol. 2008, 97, 142–149. [Google Scholar] [CrossRef]
- Lin, S.; Head, G.; Price, P.; Niu, Y.; Huang, F. Relative fitness of susceptible and Cry1A.105/Cry2Ab2-single-/dual-protein-resistant Helicoverpa zea (Boddie)(Lepidoptera: Noctuidae) on non-Bt diet and a diet containing a low concentration of two proteins. Ins. Sci. 2022, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-B.; Tabashnik, B.E.; Dennehy, T.J.; Patin, A.L.; Bartlett, A.C. Development time and resistance to Bt crops. Nature 1999, 400, 519. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.-B.; Tabashnik, B.E.; Dennehy, T.J.; Patin, A.L.; Sims, M.A.; Meyer, S.K.; Carrière, Y. Effects of Bt cotton and Cry1Ac toxin on survival and development of pink bollworm (Lepidoptera: Gelechiidae). J. Econ. Entomol. 2001, 94, 1237–1242. [Google Scholar] [CrossRef] [PubMed]
- Malthankar, P.A.; Gujar, G.T. Toxicity of Bacillus thuringiensis Cry2Ab and inheritance of Cry2Ab resistance in the pink bollworm, Pectinophora gossypiella (Saunders). Indian. J. Exp. Biol. 2016, 54, 586–596. [Google Scholar] [PubMed]
- Meihls, L.N.; Higdon, M.L.; Ellersieck, M.R.; Tabashnik, B.E.; Hibbard, B.E. Greenhouse-selected resistance to Cry3Bb1-producing corn in three western corn rootworm populations. PLoS ONE 2012, 7, e51055. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meihls, L.N.; Frank, D.L.; Ellersieck, L.R.; Hibbard, B.E. Development and characterization of MIR604 resistance in a western corn rootworm population (Coleoptera: Chrysomelidae). Environ. Entomol. 2016, 45, 526–536. [Google Scholar] [CrossRef] [Green Version]
- Mittal, A.; Ningthoukongjan, J.; Malthankar, P.A.; Ningthoujam, K.; Kalia, V.; Gujar., G.T. Toxicity of Cry1Ac, inheritance of Cry1Ac resistance and fitness costs association in the pink bollworm, Pectinophora gossypiella (Lepidoptera: Gelechiidae). Biopestic. Int. 2016, 12, 127–138. [Google Scholar]
- Niu, Y.; Head, G.P.; Price, P.A.; Huang, F. Inheritance and fitness costs of Cry1A.105 resistance in two strains of Spodoptera frugiperda (J.E. Smith). Crop Prot. 2018, 110, 229–235. [Google Scholar] [CrossRef]
- Orpet, R.J.; Degain, B.A.; Unnithan, G.C.; Welch, K.L.; Tabashnik, B.E.; Carrière, Y. Effects of dietary protein to carbohydrate ratio on Bt toxicity and fitness costs of resistance in Helicoverpa zea. Entomol. Exp. Appl. 2015, 156, 28–36. [Google Scholar] [CrossRef]
- Oswald, K.J.; French, B.W.; Nielson, C.; Bagley, M. Assessment of fitness costs in Cry3Bb1-resistant and susceptible western corn rootworm (Coleoptera: Chrysomelidae) laboratory colonies. J. Appl. Entomol. 2012, 136, 730–740. [Google Scholar] [CrossRef]
- Paddock, K.J.; Hibbard, B.E.; Barry, J.; Sethi, A.; Mueller, A.L.; Shelbyd, K.J.; Pereira, A.E. Restoration of susceptibility following removal of selection for Cry34/35Ab1 resistance documents fitness costs in resistant population of western corn rootworm, Diabrotica virgifera virgifera. Pest. Manag. Sci. 2021, 77, 2385–2394. [Google Scholar] [CrossRef] [PubMed]
- Paolino, A.R.; Gassmann, A.J. Assessment of inheritance and fitness costs associated with field-evolved resistance to Cry3Bb1 maize by western corn rootworm. Toxins 2017, 9, 159. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pereira, E.J.G.; Storer, N.P.; Siegfried, B.D. Fitness costs of Cry1F resistance in laboratory-selected European corn borer (Lepidoptera: Crambidae). J. Appl. Entomol. 2011, 135, 17–24. [Google Scholar] [CrossRef]
- Petzold-Maxwell, J.L.; Cibils-Stewart, X.; Wade French, B.; Gassmann, A.J. Adaptation by western corn rootworm (Coleoptera: Chrysomelidae) to Bt maize: Inheritance, fitness costs, and feeding preference. J. Econ. Entomol. 2012, 105, 1407–1418. [Google Scholar] [CrossRef] [Green Version]
- Petzold-Maxwell, J.L.; Siegfried, B.D.; Hellmich, R.L.; Abel, C.A.; Coates, B.S.; Spencer, T.A.; Gassmann, A.J. Effect of maize lines on larval fitness costs of Cry1F resistance in the European corn borer (Lepidoptera: Crambidae). J. Econ. Entomol. 2014, 107, 764–772. [Google Scholar] [CrossRef] [Green Version]
- Santos-Amaya, O.F.; Tavares, C.S.; Monteiro, H.M.; Teixeira, T.P.M.; Guedes, R.N.C.; Alves, A.P.; Pereira, E.J.G. Genetic basis of Cry1F resistance in two Brazilian populations of fall armyworm, Spodoptera frugiperda. Crop Prot. 2016, 81, 154–162. [Google Scholar] [CrossRef]
- van Rensburg, J.B.J. First report of field resistance by the stem borer, Busseola fusca (Fuller) to Bt-transgenic maize. South Afr. J. Plant and Soil. 2007, 24, 147–151. [Google Scholar] [CrossRef] [Green Version]
- Vélez, A.M.; Spencer, T.A.; Alves, A.P.; Crespo, A.L.B.; Siegfried, B.D. Fitness costs of Cry1F resistance in fall armyworm, Spodoptera frugiperda. J. Appl. Entomol. 2014, 138, 315–325. [Google Scholar] [CrossRef]
- Wade French, B.; Hammack, L.; Tallamy, D.W. Mating success, longevity, and fertility of Diabrotica virgifera virgifera LeConte (Chrysomelidae: Coleoptera) in relation to body size and Cry3Bb1-resistant and Cry3Bb1-susceptible genotypes. Insects 2015, 6, 943–960. [Google Scholar] [CrossRef] [Green Version]
- Wangila, D.S.; Leonard, B.R.; Bai, Y.; Head, G.P.; Huang, F. Larval survival and plant injury of Cry1Ab-susceptible, -resistant, and -heterozygous genotypes of the sugarcane borer on transgenic corn containing single or pyramided Bt genes. Crop Prot. 2012, 42, 108–115. [Google Scholar] [CrossRef] [Green Version]
- Williams, J.L.; Ellers-Kirk, C.; Orth, R.G.; Gassmann, A.J.; Head, G.; Tabashnik, B.E.; Carrière, Y. Fitness cost of resistance to Bt cotton linked with increased gossypol content in pink bollworm larvae. PLoS ONE 2010, 6, e21863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; Huang, F.; Leonard, B.R.; Ghimire, M. Growth and development of Bacillus thuringiensis Cry1Ab-susceptible and Cry1Ab-resistant sugarcane borer on diet and conventional maize plants. Entomol. Exp. Appl. 2009, 133, 199–207. [Google Scholar] [CrossRef]
- Yang, Y.-H.; Yang, Y.-J.; Gao, W.-Y.; Guo, J.-J.; Wu, Y.-H.; Wu, Y.D. Introgression of a disrupted cadherin gene enables susceptible Helicoverpa armigera to obtain resistance to Bacillus thuringiensis toxin Cry1Ac. Bull. Entomol. Res. 2009, 99, 175–181. [Google Scholar] [CrossRef]
- Zhang, W.; Ma, L.; Zhong, F.; Wang, Y.; Guo, Y.; Lua, Y.; Liang, G. Fitness costs of reproductive capacity and ovarian development in a Bt-resistant strain of the cotton bollworm Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). Pest Manag. Sci. 2015, 71, 870–877. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.N.; Ma, L.; Wang, B.J.; Chen, L.; Khaing, M.M.; Lu, Y.H.; Liang, G.M.; Guo, Y.Y. Reproductive cost associated with juvenile hormone in Bt-resistant strains of Helicoverpa armigera (Lepidoptera: Noctuidae). J. Econ. Entomol. 2016, 109, 2534–2542. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, X.C.; Wu, K.M.; Liang, G.M.; Guo, Y.Y. Modified female calling behaviour in Cry1Ac-resistant Helicoverpa armigera (Lepidoptera: Noctuidae). Pest Manag. Sci. 2009, 65, 353–357. [Google Scholar] [CrossRef]


{kind=link}
| Species | Country a | Food | WR/S b | Cost for Resistant Strain (%) | WRS/S b | Cost for F1 Progeny (%) | Practical Resistance c |
|---|---|---|---|---|---|---|---|
| C. virescens | US | Diet | 0.74 (0.25–1.37) | 26 | -- | -- | No |
| D. saccharalis | US | Diet | 1.09 (0.78–1.44) | 0 | 1.08 (0.95–1.24) | 0 | No |
| H. armigera | CH | Diet | 0.53 (0.35–0.73) | 47 | -- | -- | No |
| H. zea | US | Diet | 1.08 (0.73–1.46) | 0 | 1.09 (0.93–1.28) | 0 | Yes |
| O. nubilalis | US | Diet | 0.64 (0.36–0.98) | 36 | 1.12 (0.99–1.29) | 0 | No |
| P. gossypiella | IN | Diet | 0.78 (0.41–1.23) | 22 | 0.92 (0.79–1.08) | 8 | Yes |
| P. gossypiella | US | Diet | 0.79 (0.58–1.00) | 21 | 0.94 (0.86–1.03) | 6 | No |
| S. frugiperda | BR | Diet | 0.42 (0.00–0.96) | 58 | -- | -- | Yes |
| S. frugiperda | US | Diet | 0.89 (0.67–1.13) | 11 | 1.03 (0.94–1.12) | 0 | Yes |
| B. fusca | SA | Corn | 0.93 (0.53–1.39) | 7 | -- | -- | Yes |
| D. saccharalis | US | Corn | 1.14 (0.84–1.45) | 0 | 1.10 (0.99–1.23) | 0 | No |
| D. v. virgifera | US | Corn | 0.85 (0.71–0.99) | 15 | 0.99 (0.85–1.16) | 1 | Yes |
| O. nubilalis | US | Corn | 0.76 (0.26–1.38) | 24 | 0.96 (0.77–1.20) | 4 | No |
| S. frugiperda | BR | Corn | 0.94 (0.73–1.14) | 6 | 1.08 (1.00–1.16) | 0 | Yes |
| S. frugiperda | US | Corn | 0.92 (0.69–1.16) | 8 | 1.06 (0.95–1.18) | 0 | Yes |
| H. armigera | AU | Cotton | 0.64 (0.36–0.96) | 36 | 0.88 (0.75–1.03) | 12 | No |
| H. zea | US | Cotton | 0.92 (0.66–1.20) | 8 | 0.97 (0.86–1.09) | 3 | Yes |
| P. gossypiella | US | Cotton | 0.80 (0.60–1.00) | 20 | 0.90 (0.83–0.97) | 10 | No |
| S. frugiperda | BR | Cotton | 0.74 (0.38–1.17) | 26 | 1.00 (0.86–1.17) | 0 | Yes |
| S. frugiperda | US | Cotton | 0.99 (0.58–1.46) | 1 | 1.10 (0.93–1.29) | 0 | Yes |
| Outcome | Strain Relatedness | |
|---|---|---|
| Related | Not Related | |
| Recessive cost a | 27 (8) | 8 (3) |
| Nonrecessive cost b | 13 (4) | 24 (9) |
| No cost c | 33 (10) | 26 (10) |
| Hybrid vigor d | 17 (5) | 13 (5) |
| High fitness in resistant strain e | 7 (2) | 24 (9) |
| Fitness: F1 < susceptible and resistant > susceptible f | 3 (1) | 5 (2) |
| Species | Country a | Food | IR b | Practical Resistance c |
|---|---|---|---|---|
| B. fusca | SA | Corn | 0.93 (0.38–1.60) | Yes |
| D. saccharalis | US | Corn | 0.34 (0.10–0.64) | No |
| D. v. virgifera | US | Corn | 0.69 (0.54–0.86) | Yes |
| S. frugiperda | BR | Corn | 1.00 (0.76–1.27) | Yes |
| H. armigera | AU | Cotton | 0.48 (0.25–0.75) | No |
| H. zea | US | Cotton | 0.41 (0.10–0.79) | Yes |
| P. gossypiella | US | Cotton | 0.46 (0.03–1.02) | No |
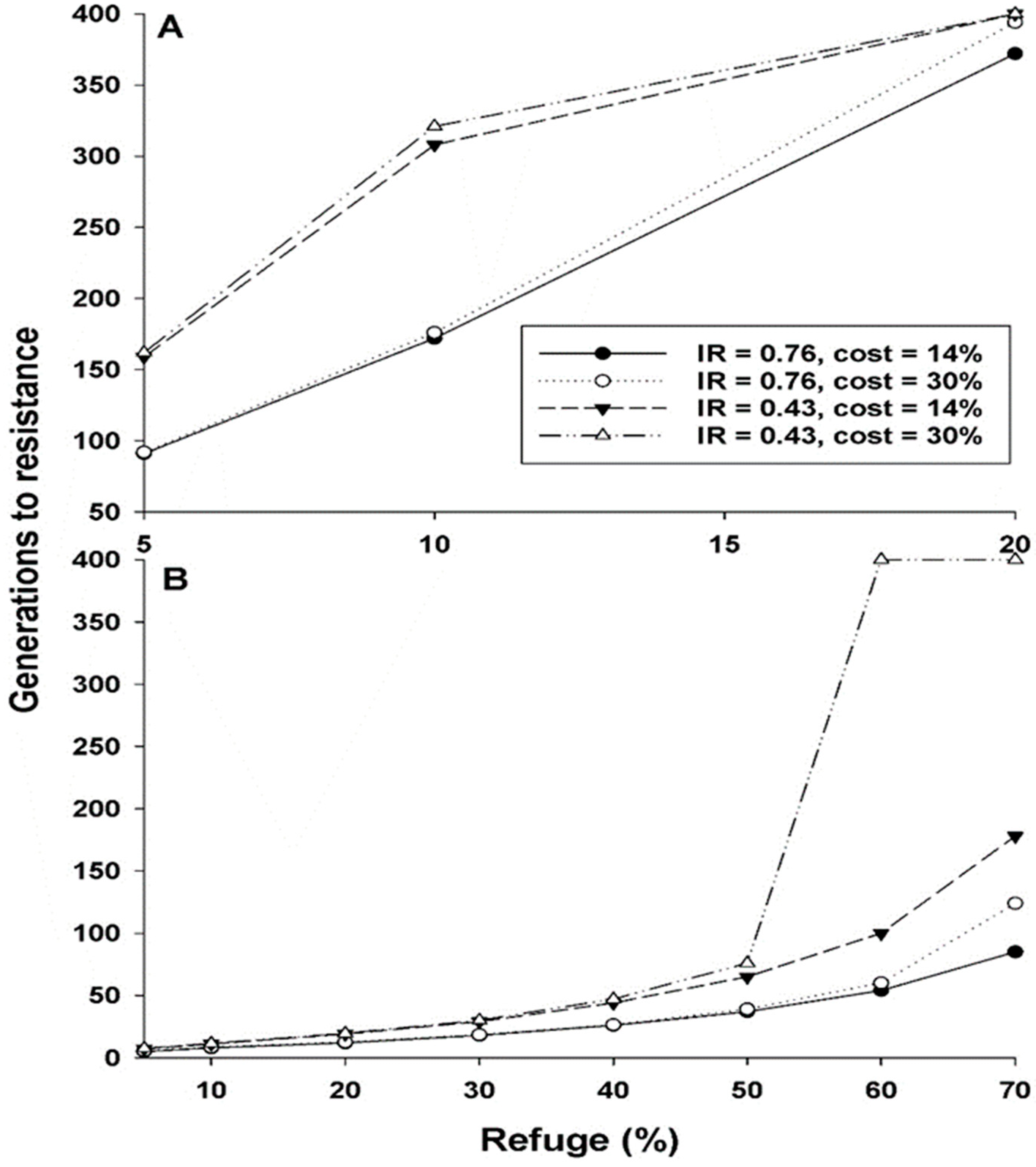
| Refuge% | No Cost | Cost = 14% | Cost = 30% | ||||||
|---|---|---|---|---|---|---|---|---|---|
| IR = | 1 | 0.76 | 0.43 | 1 | 0.76 | 0.43 | 1 | 0.76 | 0.43 |
| h = 0 | |||||||||
| 5 | 70 | 90 | 156 | 70 | 91 | 159 | 71 | 92 | 162 |
| 10 | 129 | 169 | 296 | 131 | 172 | 308 | 133 | 176 | 321 |
| 20 | 270 | 355 | >400 | 280 | 372 | >400 | 292 | 394 | >400 |
| h = 0.26 | |||||||||
| 5 | 5 | 5 | 7 | 5 | 5 | 7 | 5 | 5 | 7 |
| 10 | 7 | 8 | 11 | 7 | 8 | 11 | 7 | 8 | 11 |
| 20 | 10 | 12 | 19 | 10 | 12 | 19 | 10 | 12 | 19 |
| 30 | 15 | 18 | 29 | 15 | 18 | 29 | 15 | 18 | 30 |
| 40 | 21 | 25 | 42 | 21 | 26 | 44 | 21 | 26 | 47 |
| 50 | 29 | 35 | 60 | 30 | 37 | 65 | 31 | 39 | 76 |
| 60 | 41 | 50 | 88 | 43 | 54 | 100 | 46 | 60 | >400 |
| 70 | 61 | 76 | 133 | 66 | 85 | 178 | 77 | 124 | >400 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carrière, Y.; Tabashnik, B.E. Fitness Costs and Incomplete Resistance Associated with Delayed Evolution of Practical Resistance to Bt Crops. Insects 2023, 14, 214. https://doi.org/10.3390/insects14030214
Carrière Y, Tabashnik BE. Fitness Costs and Incomplete Resistance Associated with Delayed Evolution of Practical Resistance to Bt Crops. Insects. 2023; 14(3):214. https://doi.org/10.3390/insects14030214
Chicago/Turabian StyleCarrière, Yves, and Bruce E. Tabashnik. 2023. "Fitness Costs and Incomplete Resistance Associated with Delayed Evolution of Practical Resistance to Bt Crops" Insects 14, no. 3: 214. https://doi.org/10.3390/insects14030214