
Thermal Oviposition Performance of the Ladybird Stethorus gilvifrons Preying on Two-Spotted Spider Mites
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Rearing and Experimental Conditions
2.2. Non-Linear Models
2.3. Enkegaard Model
2.4. Bieri-1 Model
2.5. Bieri-2 Model
2.6. Analytis Model
2.7. Model Evaluation
- The coefficient of determination (R2), where RSS is the residual sum of squares and TSS is the total sum of squares. A higher value of R2 indicates a better fit.
- The residual sum of squares (RSS): A lower value of RSS indicates a better fit. The R2 is not an appropriate criterion to discriminate between models with different numbers of parameters (models with more parameters usually provide a better fit).
2.8. Statistical Analysis
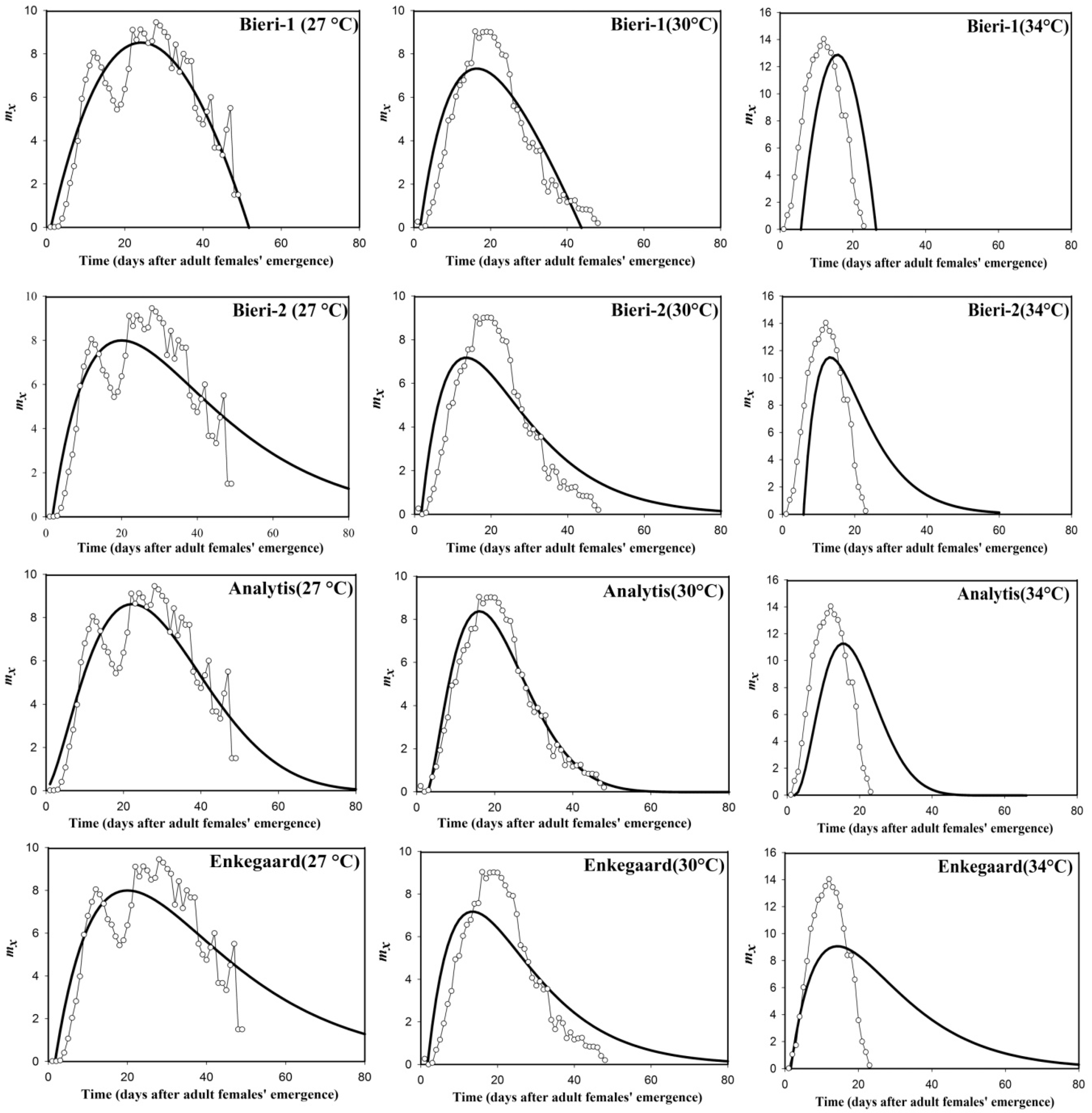
3. Results and Discussion
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Jeppson, L.R.; Keifer, H.H.; Baker, E.W. Mites Injurious to Economic Plants; University of California Press: Berkeley, CA, USA, 1975. [Google Scholar]
- Mossadegh, M.S.; Kocheili, F. A Semi Descriptive Checklist of Identified Species of Arthropods (Agricultural, Medical) and Other Pests from Khuzestan, Iran; Shahid Chamran University Press: Ahvaz, Iran, 2003. [Google Scholar]
- Van Leeuwen, T.; Vontas, J.; Tsagkarakou, A.; Dermauw, W.; Tirry, L. Acaricide resistance mechanisms in the two-spotted spider mite Tetranychus urticae and other important Acari: A review. Insect Biochem. Mol. Biol. 2010, 40, 563–572. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Helle, W.; Sabelis, M.W. Spider Mites: Their Biology, Natural Enemies, and Control, Volume 1A; Elsevier: Amsterdam, The Netherlands, 1985. [Google Scholar]
- Knapp, M.; van Houten, Y.; van Baal, E.; Groot, T. Use of predatory mites in commercial biocontrol: Current status and future prospects. Acarologia 2018, 58, 72–82. [Google Scholar] [CrossRef]
- Mathur, L.M.L. Bionomics of Stethorus gilvifrons Mulsant (Coleoptera: Coccinellidae). Madras Agric. J. 1969, 56, 7–11. [Google Scholar]
- Roy, M.; Brodeur, J.; Cloutier, C. Seasonal activity of the spider mite predators Stethorus punctillum (Coleoptera: Coccinellidae) and Neoseiulus fallacis (Acarina: Phytoseiidae) in raspberry, two predators of Tetranychus mcdanieli (Acarina: Tetranychidae). Biol. Control 2005, 34, 47–57. [Google Scholar] [CrossRef]
- McMurtry, J.A.; Huffaker, C.B.; Van de Vrie, M. Ecology of tetranychid mites and their natural enemies: A review. I. Tetranychid enemies: Their biological characters and the impact of spray practices. Hilgardia. 1970, 40, 331–390. [Google Scholar] [CrossRef] [Green Version]
- Roy, M.; Brodeur, J.; Cloutier, C. Effect of temperature on intrinsic rates of natural increase (rm) of a coccinellid and its spider mite prey. BioControl 2003, 48, 7–72. [Google Scholar] [CrossRef]
- Putman, W.L. The bionomics of Stethorus punctillum Weise (Coleoptera: Coccinellidae) in Ontario. Can. Entomol. 1955, 87, 9–33. [Google Scholar] [CrossRef] [Green Version]
- Roy, M.; Brodeur, J.; Cloutier, C. Relationship between temperature and developmental rate of Stethorus punctillum (Coleoptera: Coccinellidae) and its prey Tetranychus mcdanieli (Acarina: Tetranychidae). Environ. Èntomol. 2002, 31, 177–187. [Google Scholar] [CrossRef] [Green Version]
- Riddick, E.W.; Wu, Z. Lima bean–lady beetle interactions: Hooked trichomes affect survival of Stethorus punctillum larvae. BioControl 2011, 56, 5–63. [Google Scholar] [CrossRef]
- Houck, M.A. Prey preference in Stethorus punctum (Coleoptera: Coccinellidae). Environ. Èntomol. 1986, 15, 967–970. [Google Scholar] [CrossRef]
- Chazeau, J. Developpement et fecondite de Stethorus madecassus Chazeau (Coleopteres, Coccinellidae) eleve en conditions exterieures dans le sud-ouest de Madagascar. Cah. ORSTOM Ser. Biol. 1974, 25, 27–33. [Google Scholar]
- Kapur, A.P. On the Old-World species of the genus Stethorus Weise (Coleoptera, Coccinellidae). Bull. Entomol. Res. 1948, 37, 297–320. [Google Scholar] [CrossRef] [PubMed]
- Haji-Zadeh, J.; Kamali, G.K.; Assadi, H.B. Investigations on the functional response and populations fluctuations of Stethorus gilvifrons on red spider mite, Panonychus ulmi (Koch) in Karaj vicinity [Iran]. Appl. Entomol. Phytopathol. 1993, 61, 32–34. (In Persian) [Google Scholar]
- Mehrkhou, F.; Fathipour, Y.; Talebi, A.A.; Kamali, K.; Naseri, B. Population density and spatial distribution patterns of Tetranychus urticae (Acari, Tetranychidae) and its predator Stethorus gilvifrons (Coleoptera: Coccinellidae) on different agricultural crops. J. Entomol. Res. Soc. 2008, 10, 23–36. [Google Scholar]
- Biddinger, D.J.; Weber, D.C.; Hull, L.A. Coccinellidae as predators of mites: Stethorini in biological control. Biol. Control 2009, 51, 268–283. [Google Scholar] [CrossRef] [Green Version]
- Farahbakhsh, G. A Checklist of Important Insects and Other Enemies of Plants and Agricultural Products in Iran; Iranian Ministry of Agriculture: Tehran, Iran, 1961.
- Sarmah, M.; Bhattacharyya, B. Biology and feeding potential of Stethorus gilvifrons Mulsant (Coccinellidae: Coleoptera) on tea red spider mite, Oligonychus coffeae Neitner. Shashpa 2002, 9, 23–26. [Google Scholar]
- Afshari, M.; Mossadegh, S.; Soleyman-Nejadian, E.; Kamali, K. Geographical distribution and host plants of Stethorus gilvifrons (Mulsant) (Col.: Coccinellidae) and its biology under laboratory conditions in Khuzestan province. J. Agri. Sci. Nat. Res. 2007, 14, 201–210. [Google Scholar]
- Sohrabi, F.; Shishehbor, P. Functional and numerical responses of Stethorus gilvifrons Mulsant feeding on strawberry spider mite, Tetranychus turkestani Ugarov and Nikolski. Pak. J. Biol. Sci. 2007, 10, 4563–4566. [Google Scholar] [CrossRef]
- Taghizadeh, R.; Fathipour, Y.; Kamali, K. Influence of temperature on life-table parameters of Stethorus gilvifrons (Mulsant) (Coleoptera: Coccinellidae) fed on Tetranychus urticae Koch. J. Appl. Entomol. 2008, 132, 638–645. [Google Scholar] [CrossRef]
- Taghizadeh, R.; Fathipour, Y.; Kamali, K. Temperature-dependent development of acarophagous ladybird, Stethorus gilvifrons (Mulsant) (Coleoptera: Coccinellidae). J. Asia Pac. Entomol. 2008, 11, 145–148. [Google Scholar] [CrossRef]
- Jafari, M.; Goldasteh, S.; Ranjbar Aghdam, H.; Zamani, A.A.; Soleyman-Nejadian, E. Effect of temperature on two-sex life table parameters of Stethorus gilvifrons (Coleoptera: Coccinellidae) feeding on Tetranychus urticae (Acari: Tetranychidae). J. Entomol. Soc. Iran. 2020, 40, 65–82. (In Persian) [Google Scholar]
- Perumalsamy, K.; Selvasundaram, R.; Roobakkumar, A.; Rahman, V.J.; Muraleedharan, N. Life table and predatory efficiency of Stethorus gilvifrons (Coleoptera: Coccinellidae), an important predator of the red spider mite, Oligonychus coffeae (Acari: Tetranychidae), infesting tea. Exp. Appl. Acarol. 2010, 50, 141. [Google Scholar] [CrossRef] [PubMed]
- Jafari, M.; Goldasteh, S.; Ranjbar Aghdam, H.; Zamani, A.A.; Soleyman-Nejadian, E. Effect of solitary and group rearing on demographic parameters of Stethorus gilvifrons (Col.: Coccinellidae) feeding on Tetranychus urticae. J. Entomol. Res. 2019, 11, 197–217. [Google Scholar]
- Jafari, M.; Goldasteh, S.; Ranjbar Aghdam, H.; Zamani, A.A.; Soleyman-Nejadian, E. Effects of prey species and host plants on development and life history of Stethorus gilvifrons (Coleoptera: Coccinellidae). J. Agric. Sci. Technol. 2022, 24, 635–648. [Google Scholar]
- Cossins, A.R.; Bowler, K. Temperature Biology of Animals; Chapman and Hall: London, UK, 1987. [Google Scholar]
- Colinet, H.; Sinclair, B.J.; Vernon, P.; Renault, D. Insects in fluctuating thermal environments. Annu. Rev. Entomol. 2015, 60, 123–140. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.S.; Lee, J.H. Oviposition model of overwintered adult Tetranychus urticae (Acari: Tetranychidae) and mite phenology on the ground cover in apple orchards. Exp. Appl. Acarol. 2003, 31, 191–208. [Google Scholar] [CrossRef] [PubMed]
- Huffaker, C.B.; Berryman, A.; Turchin, P. Dynamics and regulation of insect populations. Ecol. Entomol. 1999, 2, 269–305. [Google Scholar]
- McCalla, K.A.; Keçeci, M.; Milosavljević, I.; Ratkowsky, D.A.; Hoddle, M.S. The influence of temperature variation on life history parameters and thermal performance curves of Tamarixia radiata (Hymenoptera: Eulophidae), a parasitoid of the Asian citrus psyllid (Hemiptera: Liviidae). J. Econ. Èntomol. 2019, 112, 1560–1574. [Google Scholar] [CrossRef]
- Milosavljević, I.; McCalla, K.A.; Ratkowsky, D.A.; Hoddle, M.S. Effects of constant and fluctuating temperatures on development rates and longevity of Diaphorencyrtus aligarhensis (Hymenoptera: Encyrtidae). J. Econ. Èntomol. 2019, 112, 1062–1072. [Google Scholar] [CrossRef]
- Von Schmalensee, L.; Gunnarsdottir, K.H.; Näslund, J.; Gotthard, K.; Lehmann, P. Thermal performance under constant temperatures can accurately predict insect development times across naturally variable microclimates. Ecol. Lett. 2021, 24, 1633–1645. [Google Scholar] [CrossRef] [PubMed]
- Jafari, M.; Goldasteh, S.; Aghdam, H.R.; Zamani, A.A.; Soleyman-Nejadian, E.; Schausberger, P. Modeling thermal developmental trajectories and thermal requirements of the ladybird Stethorus gilvifrons. Insects 2023, 14, 11. [Google Scholar] [CrossRef] [PubMed]
- Bieri, M.; Baumgartner, J.; Bianchi, G.; Delucchi, V.; Arx, R.V. Development and fecundity of pea aphid (Acyrthosiphon pisum Harris) as affected by constant temperatures and by pea varieties. Mitt. Schweiz. Entomol Ges. 1983, 56, 163–171. [Google Scholar]
- Enkegaard, A. The poinsettia strain of the cotton whitefly, Bemisia tabaci (Homoptera: Aleyrodidae), biological and demographic parameters on poinsettia (Euphorbia pulcherrima) in relation to temperature. Bull. Entomol. Res. 1993, 83, 535–546. [Google Scholar] [CrossRef]
- Lanzoni, A.; Accinelli, G.; Bazzocchi, G.G.; Burgio, G. Biological traits and life table of the exotic Harmonia axyridis compared with Hippodamia variegata, and Adalia bipunctata (Col., Coccinellidae). J. Appl. Entomol. 2004, 128, 298–306. [Google Scholar] [CrossRef]
- Kontodimas, D.C.; Milonas, P.G.; Stathas, G.J.; Economou, L.P.; Kavallieratos, N.G. Life table parameters of the pseudococcid predators Nephus includens and Nephus bisignatus (Coleoptera: Coccinellidae). Eur. J. Entomol. 2007, 104, 407. [Google Scholar] [CrossRef] [Green Version]
- Kontodimas, D.C. Nonlinear models for describing development and fecundity of the pseudococcid predators Nephus includens and Nephus bisignatus. Entomol. Hellenica 2012, 21, 13–24. [Google Scholar] [CrossRef] [Green Version]
- Arbab, A.; Kontodimas, D.C.; Sahragard, A. Estimating development of Aphis pomi (De Geer) (Homoptera: Aphididae) using linear and nonlinear models. Environ. Èntomol. 2006, 35, 1208–1215. [Google Scholar] [CrossRef]
- Zamani, A.A.; Talebi, A.; Fathipour, Y.; Baniameri, V. Effect of temperature on life history of Aphidius colemani and Aphidius matricariae (Hymenoptera: Braconidae), two parasitoids of Aphis gossypii and Myzus persicae (Homoptera: Aphididae). Environ. Èntomol. 2007, 36, 263–271. [Google Scholar] [CrossRef]
- Aghdam, H.R.; Fathipour, Y.; Radjabi, G.; Rezapanah, M. Temperature-dependent development and temperature thresholds of codling moth (Lepidoptera: Tortricidae) in Iran. Environ. Èntomol. 2009, 38, 885–895. [Google Scholar] [CrossRef]
- Aghdam, H.R.; Fathipour, Y.; Kontodimas, D.C. Evaluation of non-linear models to describe development and fertility of codling moth at constant temperatures. Entomol. Hell. 2011, 20, 3–16. [Google Scholar] [CrossRef] [Green Version]
- Analytis, S. Obtaining of sub-models for modeling the entire life cycle of a pathogen/Über die Erlangung von Sub-Modellen, die zur Beschreibung eines gesamten Lebenszyklus eines Krankheitserregers dienen. Z. Pflanzenkrankh. Pflanzenschutz/J. Plant Dis. Prot. 1980, 87, 371–382. [Google Scholar]
- Analytis, S. Relationship between temperature and development times in phytopathogenic fungus and in plant pests: A mathematical model. J. Agric. Res. 1981, 5, 133–159. [Google Scholar]
- Chi, H.; Liu, H. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Chi, H. TWOSEX-MS Chart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis. 2017. Available online: http://140.120.197.173/Ecology/ (accessed on 16 September 2017).
- Marquardt, D.V. An algorithm for least squares estimation of nonlinear parameters. J. Soc. Ind. Appl. Math. 1963, 11, 431–441. [Google Scholar] [CrossRef]
- Aksit, T.; Cakmak, I.; Ozer, G. Effect of temperature and photoperiod on development and fecundity of an acarophagous ladybird beetle, Stethorus gilvifrons. Phytoparasitica 2007, 35, 357–366. [Google Scholar] [CrossRef]
- Mori, K.; Nozawa, M.; Arai, K.; Gotoh, T. Life-history traits of the acarophagous lady beetle, Stethorus japonicus at three constant temperatures. BioControl 2005, 50, 35–51. [Google Scholar] [CrossRef]
- Wang, S.; Tan, X.L.; Michaud, J.P.; Zhang, F.; Guo, X. Light intensity and wavelength influence development, reproduction and locomotor activity in the predatory flower bug Orius sauteri (Poppius) (Hemiptera: Anthocoridae). BioControl 2013, 58, 667–674. [Google Scholar] [CrossRef] [Green Version]
- Emana, G.D. Comparative studies of the influence of relative humidity and temperature on the longevity and fecundity of the parasitoid, Cotesia flavipes. J. Insect Sci. 2007, 7, 19. [Google Scholar]
- Kim, D.S.; Lee, J.H. Oviposition model of Carposina sasakii (Lepidoptera: Carposinidae). Ecol. Modell. 2003, 162, 145–153. [Google Scholar] [CrossRef]
- Tougeron, K.; Ferrais, L.; Renard, M.-E.; Hance, T. Effects of constant versus fluctuating temperatures on fitness indicators of the aphid Dysaphis plantaginea and the parasitoid Aphidius matricariae. Insects 2021, 12, 855. [Google Scholar] [CrossRef] [PubMed]
- Rismayani; Ullah, S.M.; Chi, H.; Gotoh, T. Impact of constant and fluctuating temperatures on population characteristics of Tetranychus pacificus (Acari: Tetranychidae). J. Econ. Entomol. 2021, 114, 639–651. [Google Scholar] [CrossRef] [PubMed]
- Hoskins, J.L.; Janion-Scheepers, C.; Ireland, E.; Monro, K.; Chown, S.L. Constant and fluctuating temperature acclimations have similar effects on phenotypic plasticity in springtails. J. Therm. Biol. 2020, 93, 102690. [Google Scholar] [CrossRef] [PubMed]
- Lee, J.H.; Ahn, J.J. Temperature effects on development, fecundity, and life table parameters of Amblyseius womersleyi (Acari: Phytoseiidae). Environ. Entomol. 2000, 29, 265–271. [Google Scholar] [CrossRef]
- Nguyen, T.V.; Shih, C.I.T. Development of Neoseiulus womersleyi (Schicha) and Euseius ovalis (Evans) feeding on four tetranychid mites (Acari: Phytoseiidae, Tetranychidae) and pollen. J. Asia Pac. Entomol. 2010, 13, 289–296. [Google Scholar] [CrossRef]
- McMurtry, J.A.; De Moraes, G.J.; Sourassou, N.F. Revision of the lifestyles of phytoseiid mites (Acari: Phytoseiidae) and implications for biological control strategies. Syst. Appl. Acarol. 2013, 18, 297–320. [Google Scholar] [CrossRef] [Green Version]



{kind=link}
{kind=link}
| Model | N | Parameters | 15 °C | 20 °C | 25 °C | 27 °C | 30 °C | 34 °C |
|---|---|---|---|---|---|---|---|---|
| Bieri-1 | 4 | a | −0.1824 | −1.099 | −0.1881 | −1.2885 | −0.418 | −18.2343 |
| b | 0.9844 | 0.9849 | 0.9726 | 0.9773 | 0.9094 | 0.9868 | ||
| xmax | 99.17 | 96.8981 | 84.5666 | 78.6850 | 44.4851 | 92.1040 | ||
| xmin | 184.95 | 309.2969 | 100.8353 | 221.4891 | 32.0370 | 562.1977 | ||
| R2 | 0.6935 | 0.8804 | 0.8055 | 0.8180 | 0.8251 | 0.3507 | ||
| R2adj | 0.6664 | 0.8729 | 0.7969 | 0.8059 | 0.8132 | 0.2482 | ||
| RSS(10−4) | 2.84 | 21.80 | 16.65 | 68.80 | 78.56 | 1136.86 | ||
| Bieri-2 | 3 | a | 0.1121 | 0.6027 | 0.3509 | 1.1872 | 1.6660 | 4.2338 |
| b | 1.033 | 1.050 | 1.0437 | 1.0560 | 1.0890 | 1.1448 | ||
| xmin | 0.2814 | 2.4778 | 2.3966 | 1.6891 | 1.7523 | 5.8095 | ||
| R2 | 0.6915 | 0.6878 | 0.7219 | 0.7525 | 0.7677 | 0.4052 | ||
| R2adj | 0.6739 | 0.6656 | 0.7139 | 0.7418 | 0.7574 | 0.3457 | ||
| RSS(10−4) | 2.87 | 163.53 | 25.47 | 102.32 | 123.33 | 5238.56 | ||
| Analytis | 5 | a | 3.17 × 10−16 | 1.47 × 10−14 | 1.72 × 10−14 | 8.53 × 10−15 | 2.18 × 10−12 | 8.86 × 10−17 |
| Dmin | 1.7993 | −4.1012 | −1.9677 | −0.3099 | 1.6813 | 1.8743 | ||
| Dmax | 168.9471 | 81.0560 | 126.9143 | 111.8141 | 63.1247 | 66.1766 | ||
| n | 6.4882 | 5.6469 | 5.7270 | 6.5432 | 6.1459 | 8.5172 | ||
| m | 1.1649 | 3.2071 | 1.9327 | 1.6488 | 2.0061 | 2.2784 | ||
| R2 | 0.6737 | 0.9152 | 0.8095 | 0.8014 | 0.9454 | 0.3349 | ||
| R2adj | 0.6329 | 0.9080 | 0.7982 | 0.7833 | 0.9401 | 0.1784 | ||
| RSS(10−4) | 2.7520 | 15.89 | 16.36 | 76.64 | 24.11 | 390.37 | ||
| Enkegaard | 4 | a | −0.8739 | −9.8291 | −5.4350 | −12.8136 | −19.7534 | −19.7533 |
| b | 0.6820 | 3.9667 | 2.2659 | 7.5857 | 11.2724 | 12.7650 | ||
| c | 1.7624 | 1.7624 | 1.7624 | 1.7624 | 1.7624 | 1.7624 | ||
| d | 0.0333 | 0.0491 | 0.0428 | 0.0545 | 0.0853 | 0.0785 | ||
| R2 | 0.6915 | 0.6786 | 0.7219 | 0.7525 | 0.7677 | 0.3921 | ||
| R2adj | 0.6643 | 0.6586 | 0.7097 | 0.7360 | 0.7519 | 0.2961 | ||
| RSS(10−4) | 2.87 | 63.53 | 25.47 | 102.32 | 123.33 | 333.64 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jafari, M.; Aghdam, H.R.; Zamani, A.A.; Goldasteh, S.; Soleyman-Nejadian, E.; Schausberger, P. Thermal Oviposition Performance of the Ladybird Stethorus gilvifrons Preying on Two-Spotted Spider Mites. Insects 2023, 14, 199. https://doi.org/10.3390/insects14020199
Jafari M, Aghdam HR, Zamani AA, Goldasteh S, Soleyman-Nejadian E, Schausberger P. Thermal Oviposition Performance of the Ladybird Stethorus gilvifrons Preying on Two-Spotted Spider Mites. Insects. 2023; 14(2):199. https://doi.org/10.3390/insects14020199
Chicago/Turabian StyleJafari, Maryam, Hossein Ranjbar Aghdam, Abbas Ali Zamani, Shila Goldasteh, Ebrahim Soleyman-Nejadian, and Peter Schausberger. 2023. "Thermal Oviposition Performance of the Ladybird Stethorus gilvifrons Preying on Two-Spotted Spider Mites" Insects 14, no. 2: 199. https://doi.org/10.3390/insects14020199