

From Field Data to Practical Knowledge: Investigating the Bioecology of the Oak Lace Bug—An Invasive Insect Species in Europe

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Area and Data Collection
2.2. Developing the Insect Development Prediction Model Based on Degree days
2.3. Constructing the Life Table of the Insect Population in a Natural Environment
2.4. Data Analyses
3. Results
3.1. Life Cycle and Number and Durations of Generations
3.2. Using Degree Days to Predict Insect Development
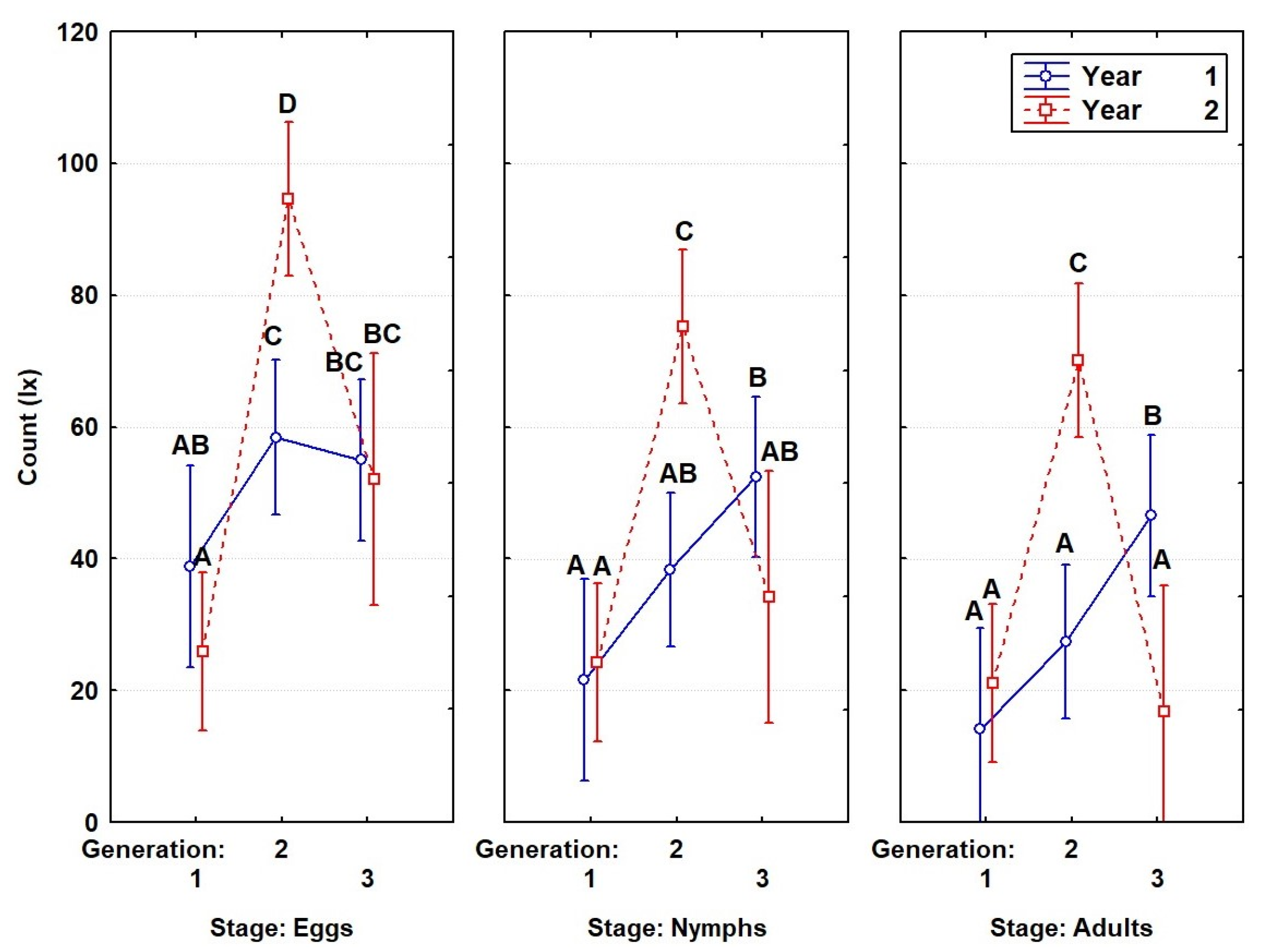
3.3. Life Table of the Insect Population in a Natural Environment
3.4. Analysis of Differences in Egg Laying Trends among Generations
4. Discussion
4.1. Life Cycle and Number and Durations of Generations
4.2. Using Degree Days to Predict Insect Development
4.3. Life Table of the Insect Population in a Natural Environment
4.4. Analysis of Differences in Egg Laying Trends among Generations
4.5. Practical Implications and Future Research Directions
- Continuing periodic observations of the insect’s biology to refine and improve our degree day–based prediction model;
- Gaining a deeper understanding of how climatic conditions affect the life cycle and population density of the OLB;
- Developing advanced forecasting methods based on newly acquired knowledge;
- Assessing the potential risk of mass infestation in various ecosystems and geographic areas.
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Index | Description | Method of Determination | Formula |
|---|---|---|---|
| N′ | The total number of pairs established and introduced into the experiment. | Directly counting the actual number of the pairs established and introduced into the experiment. | N/A (directly counted). |
| N | The number of pairs that successfully completed the development cycle of the respective generation. | Directly counting the actual number of pairs that successfully completed the development cycle. | N/A (directly counted). |
| The average number of eggs per pair. | Directly counting the actual number of eggs produced by each pair at the end of oviposition and calculating the average for a pair. | N/A (directly counted). | |
| The mortality and infertility of the eggs. | Determining the percentage of eggs that were non-viable due to natural enemies or other unknown biological causes and did not result in the development of first-instar nymphs. | ||
| The average number of first-instar nymphs. | Directly counting the number of nymphs resulting from the total number of eggs for each pair that survived until the end of the first stage of development and calculating the average for a pair. | N/A (directly counted). | |
| The mortality of first-instar nymphs. | Determining the percentage of first-instar nymphs that did not survive until the end of the first instar due to natural enemies or other unknown biological causes. | ||
| The average number of second-instar nymphs. | Directly counting the number of nymphs resulting from the first-instar nymphs that survived until the end of the second instar and calculating the average for a pair. | N/A (directly counted). | |
| The mortality of second-instar nymphs. | Determining the percentage of nymphs derived from first-instar nymphs that did not survive until the end of the second instar due to natural enemies or other unknown biological causes. | ||
| The average number of third-instar nymphs. | Directly counting the number of nymphs resulting from second-instar nymphs that survived until the end of the third instar and calculating the average for a pair. | N/A (directly counted). | |
| The mortality of third-instar nymphs. | Calculating the percentage of nymphs originating from second-instar nymphs that did not survive until the end of the third instar due to natural enemies or other unknown biological causes. | ||
| The average number of fourth-instar nymphs. | Directly counting the number of nymphs resulting from third-instar nymphs that survived until the end of the fourth instar and calculating the average for a pair. | N/A (directly counted). | |
| The mortality of fourth-instar nymphs. | Calculating the percentage of nymphs originating from third-instar nymphs that did not survive until the end of the fourth instar due to natural enemies or other unknown biological causes. | ||
| The average number of fifth-instar nymphs. | Directly counting the number of nymphs resulting from fourth-instar nymphs that survived until the end of the fifth instar and calculating the average for a pair. | N/A (directly counted). | |
| The mortality of fifth-instar nymphs. | Determining the percentage of nymphs derived from fourth-instar nymphs that did not survive until the end of the fifth instar due to natural enemies or other unknown biological causes. | ||
| The average number of adults. | Directly counting the number of adults resulting from fifth-instar nymphs and calculating the average for a pair. | ||
| The sex ratio. | The ratio of the number of females to the number of adults | ||
| GS | The survival of the generation. | the population trend index. Without the effects of fertility and mortality of individuals after reaching adulthood. |
References
- Williamson, M.; Fitter, A. The Varying Success of Invaders. Ecology 1996, 77, 1661–1666. [Google Scholar] [CrossRef]
- Richardson, D.M.; Pyšek, P.; Rejmanek, M.; Barbour, M.G.; Panetta, F.D.; West, C.J. Naturalization and Invasion of Alien Plants: Concepts and Definitions. Divers. Distrib. 2000, 6, 93–107. [Google Scholar] [CrossRef]
- Juliano, S.A.; Philip Lounibos, L. Ecology of Invasive Mosquitoes: Effects on Resident Species and on Human Health. Ecol. Lett. 2005, 8, 558–574. [Google Scholar] [CrossRef] [PubMed]
- Pyšek, P.; Richardson, D.M. Invasive Species, Environmental Change and Management, and Health. Annu. Rev. Environ. Resour. 2010, 35, 25–55. [Google Scholar] [CrossRef]
- Simberloff, D.; Martin, J.-L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M. Impacts of Biological Invasions: What’s What and the Way Forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef]
- Davis, M.A.; Grime, J.P.; Thompson, K. Fluctuating Resources in Plant Communities: A General Theory of Invasibility. J. Ecol. 2000, 88, 528–534. [Google Scholar] [CrossRef]
- Lounibos, L.P. Invasions by Insect Vectors of Human Disease. Annu. Rev. Entomol. 2002, 47, 233–266. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, T.M.; Essl, F.; Evans, T.; Hulme, P.E.; Jeschke, J.M.; Kühn, I.; Kumschick, S.; Marková, Z.; Mrugała, A.; Nentwig, W. A Unified Classification of Alien Species Based on the Magnitude of Their Environmental Impacts. PLoS Biol. 2014, 12, e1001850. [Google Scholar] [CrossRef]
- Pimentel, D.; Lach, L.; Zuniga, R.; Morrison, D. Environmental and Economic Costs of Nonindigenous Species in the United States. Bioscience 2000, 50, 53–65. [Google Scholar] [CrossRef]
- Lovell, S.J.; Stone, S.F.; Fernandez, L. The Economic Impacts of Aquatic Invasive Species: A Review of the Literature. Agric. Resour. Econ. Rev. 2006, 35, 195–208. [Google Scholar] [CrossRef]
- Meyerson, L.A.; Mooney, H.A. Invasive Alien Species in an Era of Globalization. Front. Ecol. Environ. 2007, 5, 199–208. [Google Scholar] [CrossRef]
- Vilà, M.; Basnou, C.; Pyšek, P.; Josefsson, M.; Genovesi, P.; Gollasch, S.; Nentwig, W.; Olenin, S.; Roques, A.; Roy, D. How Well Do We Understand the Impacts of Alien Species on Ecosystem Services? A Pan-European, Cross-taxa Assessment. Front. Ecol. Environ. 2010, 8, 135–144. [Google Scholar] [CrossRef]
- Vilà, M.; Espinar, J.L.; Hejda, M.; Hulme, P.E.; Jarošík, V.; Maron, J.L.; Pergl, J.; Schaffner, U.; Sun, Y.; Pyšek, P. Ecological Impacts of Invasive Alien Plants: A Meta-analysis of Their Effects on Species, Communities and Ecosystems. Ecol. Lett. 2011, 14, 702–708. [Google Scholar] [CrossRef]
- Jeschke, J.M.; Keesing, F.; Ostfeld, R.S. Novel Organisms: Comparing Invasive Species, GMOs, and Emerging Pathogens. Ambio 2013, 42, 541–548. [Google Scholar] [CrossRef] [PubMed]
- Hulme, P.E. Invasive Species Challenge the Global Response to Emerging Diseases. Trends Parasitol. 2014, 30, 267–270. [Google Scholar] [CrossRef]
- Schindler, S.; Staska, B.; Adam, M.; Rabitsch, W.; Essl, F. Alien Species and Public Health Impacts in Europe: A Literature Review. NeoBiota 2015, 27, 1. [Google Scholar] [CrossRef]
- Bellard, C.; Cassey, P.; Blackburn, T.M. Alien Species as a Driver of Recent Extinctions. Biol. Lett. 2016, 12, 20150623. [Google Scholar] [CrossRef] [PubMed]
- Blackburn, T.M.; Bellard, C.; Ricciardi, A. Alien versus Native Species as Drivers of Recent Extinctions. Front. Ecol. Environ. 2019, 17, 203–207. [Google Scholar] [CrossRef]
- Kenis, M.; Branco, M. Impact of Alien Terrestrial Arthropods in Europe. Chapter 5. BioRisk 2010, 4, 51. [Google Scholar] [CrossRef]
- Connell, W.A.; Beacher, J.H. Life History and Control of the Oak Lace Bug. Bull. Univ. Del. Agric. Exp. Stn. 1947, 265, 5–28. [Google Scholar]
- Drake, C.J.; Ruhoff, F.A. Lacebugs of the World: A Catalog (Hemiptera: Tingidae). Chinches de Encaje Del Mundo: Un Catálogo (Hemiptera: Tingitidae). Bull. United States Natl. Mus. 1965, 243, 1–634. [Google Scholar]
- Bernardinelli, I.; Zandigiacomo, P. Prima Segnalazione Di Corythucha arcuata (Say) (Heteroptera, Tingidae) in Europa. Inf. Fitopatol. 2000, 50, 47–49. [Google Scholar]
- Forster, B.; Giacalone, I.; Moretti, M.; Dioli, P.; Wermelinger, B. Die Amerikanische Eichennetzwanze Corythucha arcuata (Say) (Heteroptera, Tingidae) Hat Die Sudschweiz Erreicht. Mitteilungen-Schweiz. Entomol. Ges. 2005, 78, 317. [Google Scholar]
- Mutun, S. First Report of the Oak Lace Bug, Corythucha arcuata (Say, 1832) (Heteroptera: Tingidae), from Bolu, Turkey. Isr. J. Zool. 2003, 49, 323–324. [Google Scholar]
- Samin, N.; Linnavuori, R.E. A Contribution to the Tingidae (Heteroptera) from North and Northwestern Iran. Entomofauna 2011, 32, 373–380. [Google Scholar]
- Dobreva, M.; Simov, N.; Georgiev, G.; Mirchev, P.; Georgieva, M. First Record of Corythucha arcuata (Say) (Heteroptera: Tingidae) on the Balkan Peninsula. Acta Zool. Bulg. 2013, 65, 409–412. [Google Scholar]
- Csóka, G.; Hirka, A.; Somlyai, M. A Tölgy Csipkéspoloska (Corythuca arcuata Say, 1832–Hemiptera, Tingidae) Elsô Észlelése Magyarországon. Novenyvedelem 2013, 49, 293–296. [Google Scholar]
- Hrašovec, B.; Posarić, D.; Lukić, I.; Pernek, M. Prvi Nalaz Hrastove Mrežaste Stjenice (Corythucha arcuata) u Hrvatskoj. Sumar. List. 2013, 137, 499–503. [Google Scholar]
- Poljaković-Pajnik, L.; Drekić, M.; Pilipović, A.; Nikolić, N.; Pap, P.; Vasić, V.; Marković, M. Pojava Velikih Šteta Od Corythucha arcuata (Say)(Heteroptera: Tingidae) u Šumama Hrasta u Vojvodini. XIII Savetovanje o Zaštiti Bilja. Zb. Rad. 2015, 63, 179–181. [Google Scholar]
- Pap, P.; Drekić, M.; Poljaković-Pajnik, L.; Marković, M.; Vasić, V. Monitoring Zdravstvenog Stanja Šuma Na Teritoriji Vojvodine u 2015. Godini. Topola 2015, 195, 117–133. [Google Scholar]
- Neimorovets, V.V.; Shchurov, V.I.; Bondarenko, A.S.; Skvortsov, M.M.; Konstantinov, F. V First Documented Outbreak and New Data on the Distribution of Corythucha arcuata (Say, 1832)(Hemiptera: Tingidae) in Russia. Acta Zool. Bulg. 2017, 9, 139–142. [Google Scholar]
- Chireceanu, C.; Teodoru, A.; Chiriloaie, A. First Record of Oak Lace Bug Corythucha arcuata (Tingidae: Heteroptera) in Romania. In Proceedings of the 7th ESENIAS Workshop with Scientific Conference, Sofia, Bulgaria, 28–30 March 2017; pp. 28–30. [Google Scholar]
- Don, I.; Don, C.D.; Sasu, L.R.; Vidrean, D.; Brad, M.L. Insect Pests on the Trees and Shrubs from the Macea Botanical Garden. Stud. Univ. Vasile Goldis Arad. 2016, 11, 23–28. [Google Scholar]
- Csóka, G.; Hirka, A.; Mutun, S.; Glavendekić, M.; Mikó, Á.; Szőcs, L.; Paulin, M.; Eötvös, C.B.; Gáspár, C.; Csepelényi, M. Spread and Potential Host Range of the Invasive Oak Lace Bug [Corythucha arcuata (Say, 1832)–Heteroptera: Tingidae] in Eurasia. Agric. For. Entomol. 2019, 22, 61–74. [Google Scholar] [CrossRef]
- Jurc, M.; Jurc, D. The First Record and the Beginning the Spread of Oak Lace Bug, Corythucha arcuata (Say, 1832) (Heteroptera: Tingidae), in Slovenia. Sumar. List 2017, 141, 485–488. [Google Scholar] [CrossRef]
- Dautbašić, M.; Zahirović, K.; Mujezinović, O.; Margaletić, J. Prvi Nalaz Hrastove Mrežaste Stjenice (Corythucha arcuata) u Bosni i Hercegovini. Sumar. List. 2018, 142, 179–181. [Google Scholar] [CrossRef]
- Streito, J.-C.; Balmès, V.; Aversenq, P.; Weill, P.; Chapin, E.; Clement, M.; Piednoir, F. Corythucha arcuata (Say, 1832) et Stephanitis Lauri Rietschel, 2014, Deux Espèces Invasives Nouvelles Pour La Faune de France (Hemiptera Tingidae). L’Entomologiste 2018, 74, 133–136. [Google Scholar]
- Meshkova, V. Insecte Fitofage Invazive Din Arboretele Și Parcurile Din Ucraina. Bucov. For. 2022, 22, 29–40. [Google Scholar] [CrossRef]
- Zubrik, M.; Gubka, A.; Rell, S.; Kunca, A.; Vakula, J.; Galko, J.; Nikolov, C.; Leonotvyč, R. First Record of Corythuchaarcuata in Slovakia–Short Communication. Plant Prot. Sci. 2019, 55, 129–133. [Google Scholar] [CrossRef]
- Sallmannshofer, M.; Ette, S.; Hinterstoisser, W.; Cech, T.L.; Hoch, G. Erstnachweis Der Eichennetzwanze, Corythucha arcuata, in Österreich. Aktuell 2019, 66, 20. [Google Scholar]
- May, R.M.; Hassell, M.P.; Anderson, R.M.; Tonkyn, D.W. Density Dependence in Host-Parasitoid Models. J. Anim. Ecol. 1981, 50, 855–865. [Google Scholar] [CrossRef]
- Jarosik, V.; Dixon, A.F.G. Population Dynamics of a Tree-Dwelling Aphid: Regulation and Density-Independent Processes. J. Anim. Ecol. 1999, 68, 726–732. [Google Scholar] [CrossRef]
- Norris, R.F.; Caswell-Chen, E.P.; Kogan, M. Concepts in Integrated Pest Management; Prentice Hall: Hoboken, NJ, USA, 2002. [Google Scholar]
- Supriadi, K.; Mudjiono, G.; Abadi, A.L.; Karindah, S. The Influence of Environmental Factors to the Abundance of Scales (Hemiptera: Diaspididae) Population on Apple Crop. J. Trop. Life Sci. 2015, 5, 20–24. [Google Scholar]
- Herms, D.A. Using Degree-Days and Plant Phenology to Predict Pest Activity. In IPM (Integrated Pest Management) of Midwest Landscapes; Minnesota Agricultural Experiment Station: St Paul, MN, USA, 2004; Volume 58, pp. 49–59. Available online: http://cues.cfans.umn.edu/old/Web/049DegreeDays.pdf (accessed on 25 September 2023).
- Jones, V.P.; Brunner, J.F. Degree-Day Models. Orchard. Pest. Management. Good Fruit. Grower. 1993. pp. 45–48. Available online: https://treefruit.wsu.edu/crop-protection/opm/dd-models/ (accessed on 25 September 2023).
- Kowalsick, T.; Clark, S. Using Growing Degree-Days for Insect Pest Management. In Information Brochure; Horticulture Research Laboratory, Cornell Cooperative Extension, Cornell University: Ithaca, NY, USA, 2006; Volume 4. [Google Scholar]
- Murray, M. Using Degree Days to Time Treatments for Insect Pests. 2020. Available online: https://climate.usu.edu/includes/pestFactSheets/degree-days08.pdf (accessed on 25 September 2023).
- Kakde, A.M.; Patel, K.G.; Tayade, S. Role of Life Table in Insect Pest Management—A Review. IOSR J. Agric. Vet. Sci. 2014, 7, 40–43. [Google Scholar] [CrossRef]
- Southwood, T.R.E.; Southwood, T.R.E. The Construction, Description and Analysis of Age-Specific Life-Tables. In Ecological Methods: With Particular Reference to the Study of Insect Populations; Springer: Dordrecht, The Netherlands, 1978; pp. 356–387. [Google Scholar] [CrossRef]
- Harcourt, D. The Development and Use of Life Tables in the Study of Natural Insect Populations. Annu. Rev. Entomol. 1969, 14, 175–196. [Google Scholar] [CrossRef]
- Gibson, E.H. The Genus Corythucha Stal (Tingidae; Heteroptera). Trans. Am. Entomol. Soc. 1918, 44, 69–104. [Google Scholar]
- Mutun, S.; Ceyhan, Z.; Sözen, C. Invasion by the Oak Lace Bug, Corythucha arcuata (Say)(Heteroptera: Tingidae), in Turkey. Turk. J. Zool. 2009, 33, 263–268. [Google Scholar] [CrossRef]
- Paulin, M.J.; Eötvös, C.B.; Zabransky, P.; Csóka, G.; Schebeck, M. Cold Tolerance of the Invasive Oak Lace Bug, Corythucha arcuata. Agric. For. Entomol. 2023, 2023, 612–621. [Google Scholar] [CrossRef]
- Bernardinelli, I. Distribution of the Oak Lace Bug Corythucha arcuata (Say) in Northern Italy (Heteroptera Tingidae). Redia 2000, 83, 157–162. [Google Scholar]
- Csepelényi, M.; Csókáné Hirka, A.; Szénási, Á.; Mikó, Á.; Szőcs, L.; Csóka, G. Az Inváziós Tölgy Csipkéspoloska [Corythucha arcuata (Say, 1832)] Gyors Terjeszkedése És Tömeges Fellépése Magyarországon= Rapid Area Expansion and Mass Occurrences of the Invasive Oak Lace Bug [Corythucha arcuata (Say 1932)] in Hungary. Erdészettudományi Közlemények 2017, 7, 127–134. [Google Scholar] [CrossRef]
- Dreistadt, S.H.; Perry, E.J. Lace Bugs: Integrated Pest Management for Home Gardeners and Landscape Professionals; University of California, ANR/Communications Services: Santa Barbara, CA, USA, 2014. [Google Scholar]
- Uvarov, B.P. Insects and Climate. Trans. R. Entomol. Soc. Lond. 1931, 79 Pt 1, 1–247. [Google Scholar] [CrossRef]
- Lindsey, A.A.; Newman, J.E. Use of Official Wather Data in Spring Time: Temperature Analysis of an Indiana Phenological Record. Ecology 1956, 37, 812–823. [Google Scholar] [CrossRef]
- Kocmánková, E.; Trnka, M.; Juroch, J.; Dubrovský, M.; Semerádová, D.; Možný, M.; Žalud, Z. Impact of Climate Change on the Occurrence and Activity of Harmful Organisms. Plant Prot. Sci. 2009, 45, S48–S52. [Google Scholar] [CrossRef]
- Skendžić, S.; Zovko, M.; Živković, I.P.; Lešić, V.; Lemić, D. The Impact of Climate Change on Agricultural Insect Pests. Insects 2021, 12, 440. [Google Scholar] [CrossRef] [PubMed]
- Bălăcenoiu, F.; Simon, D.C.; Nețoiu, C.; Toma, D.; Petrițan, I.C. The Seasonal Population Dynamics of Corythucha arcuata (Say, 1832)(Hemiptera: Tingidae) and the Relationship between Meteorological Factors and the Diurnal Flight Intensity of the Adults in Romanian Oak Forests. Forests 2021, 12, 1774. [Google Scholar] [CrossRef]
- Rock, G.C.; Stinner, R.E.; Bacheler, J.E.; Hull, L.A.; Hogmire, H.W., Jr. Predicting Geographical and Within-Season Variation in Male Flights of Four Fruit Pests. Environ. Entomol. 1993, 22, 716–725. [Google Scholar] [CrossRef]
- Hawthorne, D.J.; Rock, G.C.; Stinner, R.E. Redbanded Leafroller (Lepidoptera: Tortricidae): Thermal Requirements for Development and Simulation of within-Season Phenology in North Carolina. Environ. Entomol. 1988, 17, 40–46. [Google Scholar] [CrossRef]
- Adams, N.E. Using Growing Degree Days for Insect Management. University of New Hampshire. Coop. Extension. 2017. Available online: http://ccetompkins.org/resources/usinggrowing-degree-days-for-insect-management (accessed on 25 September 2023).
- University of Massachusetts Amherst. Corythucha spp. [Fact Sheet]. 2023. Available online: https://Ag.Umass.Edu/Landscape/Publications-Resources/Insect-Mite-Guide/Corythucha-Spp (accessed on 25 September 2023).
- Cornell University. Horticulture Center Demonstration & Community Gardens at East Meadow Farm Lacebug [Fact Sheet]. 2023. Available online: https://ccenassau.org/resources/lacebug (accessed on 25 September 2023).
- Hosseini-Tabesh, B.; Sahragard, A.; Karimi-Malati, A. A Laboratory and Field Condition Comparison of Life Table Parameters of Aphis gossypii Glover (Hemiptera: Aphididae). J. Plant Prot. Res. 2015, 55, 1–7. [Google Scholar] [CrossRef]
- Khaliq, A.M.; Javed, M.; Sohail, M.; Sagheer, M. Environmental Effects on Insects and Their Population Dynamics. J. Entomol. Zool. Stud. 2014, 2, 1–7. [Google Scholar]
- Ju, R.-T.; Wang, F.; Li, B. Effects of Temperature on the Development and Population Growth of the Sycamore Lace Bug, Corythucha ciliata. J. Insect Sci. 2011, 11, 16. [Google Scholar] [CrossRef] [PubMed]
- Schowalter, T.D. Insect Ecology: An Ecosystem Approach; Academic Press: Cambridge, MA, USA, 2022; ISBN 032-3-85674-8. [Google Scholar]
- Wrensch, D.L. Evolutionary Flexibility through Haploid Males or How Chance Favors the Prepared Genome. In Evolution and Diversity of Sex Ratio in Insects and Mites; Chapman & Hall: New York, NY, USA, 1993; pp. 118–149. [Google Scholar]
- Wei, Y.J. Sex Ratio of Nysius Huttoni White (Hemiptera: Lygaeidae) in Field and Laboratory Populations. N. Z. J. Zool. 2008, 35, 19–28. [Google Scholar] [CrossRef]
- Bernardinelli, I. Potential Host Plants of Corythucha Arcuata (Het., Tingidae) in Europe: A Laboratory Study. J. Appl. Entomol. 2006, 130, 480–484. [Google Scholar] [CrossRef]
- Leather, S.R. Factors Affecting Fecundity, Fertility, Oviposition, and Larviposition in Insects. In Insect Reproduction; CRC Press: Boca Raton, FL, USA, 2018; pp. 143–174. [Google Scholar]
- Awmack, C.S.; Leather, S.R. Host Plant Quality and Fecundity in Herbivorous Insects. Annu. Rev. Entomol. 2002, 47, 817–844. [Google Scholar] [CrossRef] [PubMed]
- Wilson, L.; Barnett, W. Degree-Days: An Aid in Crop and Pest Management. Calif. Agric. 1983, 37, 4–7. [Google Scholar]
- Bălăcenoiu, F.; Nețoiu, C.; Tomescu, R.; Simon, D.C.; Buzatu, A.; Toma, D.; Petrițan, I.C. Chemical Control of Corythucha arcuata (Say, 1832), an Invasive Alien Species, in Oak Forests. Forests 2021, 12, 770. [Google Scholar] [CrossRef]
- Singh, H.; Singh, M. Role of Life Table in Insect Pest Management. Vigyan Varta 2022, 3, 82–84. [Google Scholar]





| Year | Generation | Date |
|---|---|---|
| 2020 | I | 5 May 2020 |
| II | 7 July 2020 | |
| III | 6 August 2020 | |
| 2021 | I | 1 May 2021 |
| II | 1 July 2021 | |
| III | 1 August 2021 |
| Index | The Generation of 2020 | The Generation of 2021 | ||||
|---|---|---|---|---|---|---|
| G I | G II | G III | G I | G II | G III | |
| N′ | 40 | 130 | 110 | 70 | 70 | 70 |
| N | 28 | 48 | 44 | 46 | 48 | 18 |
| 38.9 ± 5 | 58.4 ± 5.8 | 55 ± 4.2 | 25.9 ± 4.2 | 94.6 ± 10.3 | 52.1 ± 8.9 | |
| 44.5% | 34.3% | 4.7% | 6.2% | 20.5% | 34.3% | |
| 21.6 ± 2.8 | 38.4 ± 5.5 | 52.4 ± 4.4 | 24.3 ± 5.2 | 75.3 ± 8.7 | 34.2 ± 10.6 | |
| 17.9% | 11.6% | 4.5% | 6.3% | 5.1% | 9.1% | |
| 17.7 ± 2.6 | 30.7 ± 4.5 | 50.5 ± 4.6 | 22.7 ± 5.1 | 71.4 ± 8.7 | 31.1 ± 10.4 | |
| 10.1% | 9.6% | 3.6% | 5.4% | 0.9% | 10.4% | |
| 15.9 ± 2.6 | 29.2 ± 4.3 | 48.2 ± 4.6 | 21.5 ± 5 | 70.7 ± 8.6 | 27.9 ± 10.5 | |
| 6.3% | 4.6% | 1.6% | 1.2% | 0.5% | 8.4% | |
| 14.9 ± 2.7 | 28.2 ± 4.2 | 47.5 ± 4.7 | 21.3 ± 5 | 70.4 ± 8.6 | 25.6 ± 10.6 | |
| 3.3% | 3.6% | 1.3% | 0.4% | 0.2% | 2.2% | |
| 14.6 ± 2.7 | 27.4 ± 4.2 | 46.8 ± 4.8 | 21.2 ± 5 | 70.2 ± 8.6 | 25 ± 10.7 | |
| 2% | 3% | 0.6% | 0.4% | 0.2% | 32.4% | |
| 14.1 ± 2.8 | 27.4 ± 4.2 | 46.5 ± 4.8 | 21.1 ± 5 | 70.1 ± 8.6 | 16.9 ± 11.5 | |
| 0.4 | 0.5 | 0.5 | 0.5 | 0.5 | 0.5 | |
| GS | 36% | 47% | 85% | 81% | 74% | 32% |
| Characteristics | Statistical Parameters | Generation | |||||
|---|---|---|---|---|---|---|---|
| I | II | III | |||||
| 2020 | 2021 | 2020 | 2021 | 2020 | 2021 | ||
| Clusters/female | Mean | 2.65 | 2.17 | 1.95 | 3.65 | 1.55 | 2.33 |
| Minimum | 1 | 1 | 1 | 2 | 1 | 1 | |
| Maximum | 7 | 3 | 6 | 9 | 3 | 5 | |
| Standard deviation | 1.64 | 0.77 | 1.08 | 1.67 | 0.74 | 1.41 | |
| Coefficient of variation (%) | 62.68 | 35.77 | 56.09 | 45.67 | 48.07 | 60.61 | |
| Number of eggs/cluster | Mean | 11.5 | 11.9 | 24.62 | 26.79 | 31.84 | 22.90 |
| Minimum | 1 | 2 | 4 | 2 | 6 | 7 | |
| Maximum | 46 | 51 | 72 | 125 | 84 | 64 | |
| Standard deviation | 10.74 | 10.80 | 15.27 | 23.11 | 21.50 | 15.34 | |
| Coefficient of variation (%) | 94.10 | 90.71 | 62.02 | 86.27 | 67.23 | 66.97 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bălăcenoiu, F.; Toma, D.; Nețoiu, C. From Field Data to Practical Knowledge: Investigating the Bioecology of the Oak Lace Bug—An Invasive Insect Species in Europe. Insects 2023, 14, 882. https://doi.org/10.3390/insects14110882
Bălăcenoiu F, Toma D, Nețoiu C. From Field Data to Practical Knowledge: Investigating the Bioecology of the Oak Lace Bug—An Invasive Insect Species in Europe. Insects. 2023; 14(11):882. https://doi.org/10.3390/insects14110882
Chicago/Turabian StyleBălăcenoiu, Flavius, Dragoș Toma, and Constantin Nețoiu. 2023. "From Field Data to Practical Knowledge: Investigating the Bioecology of the Oak Lace Bug—An Invasive Insect Species in Europe" Insects 14, no. 11: 882. https://doi.org/10.3390/insects14110882