
An LW-Opsin Mutation Changes the Gene Expression of the Phototransduction Pathway: A Cryptochrome1 Mutation Enhances the Phototaxis of Male Plutella xylostella (Lepidoptera: Plutellidae)

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Strains and Rearing
2.2. RNA Extraction and Transcriptome-Sequencing
2.3. Bioinformatic Analyses
2.4. Validation of Differentially-Expressed Genes by RT-qPCR
2.5. Phototactic Behavioral Assays
2.6. Statistical Analysis
3. Results
3.1. Transcriptome-Sequencing Data Analysis
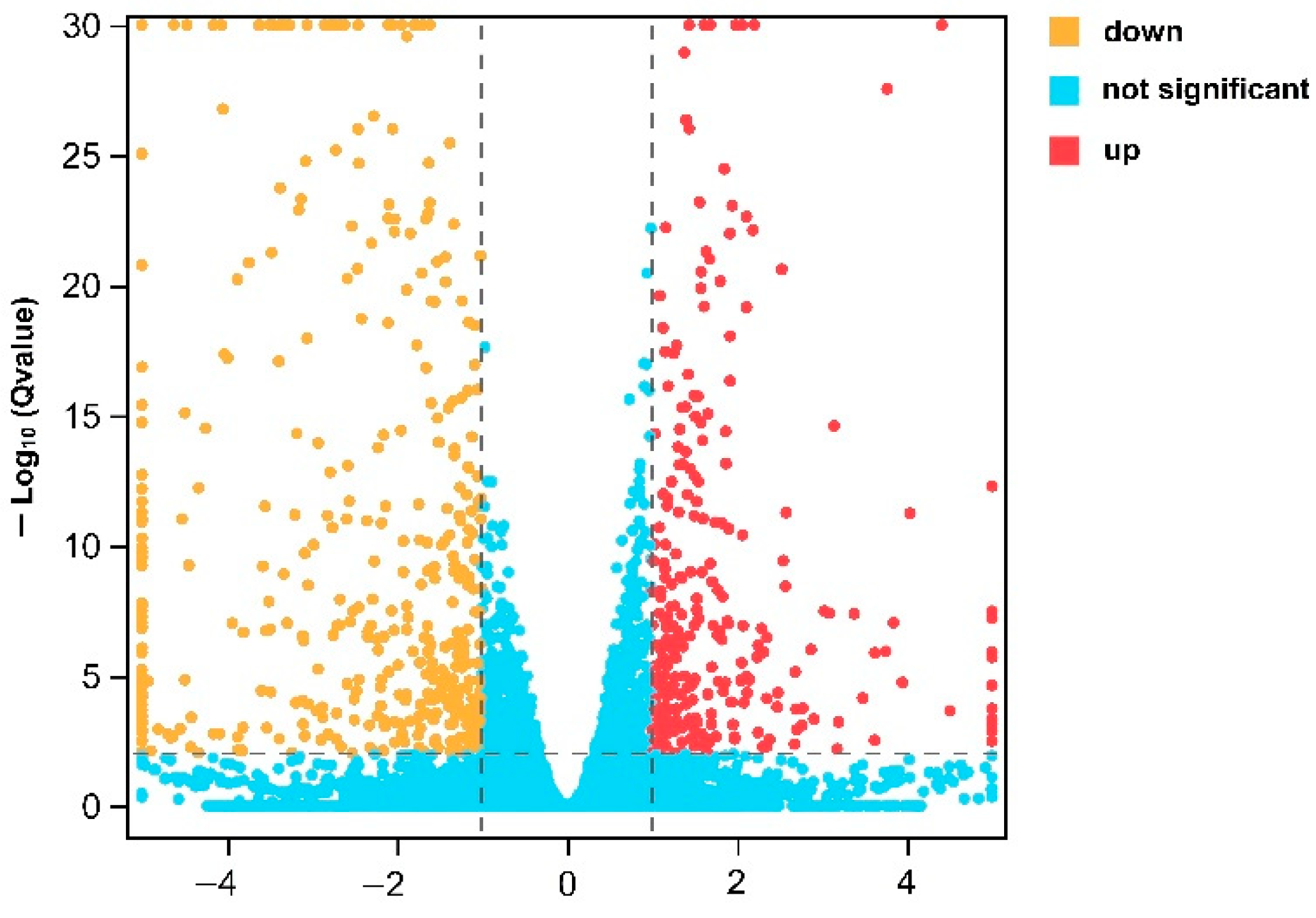
3.2. Identification of DEGs
3.3. Gene Ontology of DEGs
3.4. KEGG Pathway of DEGs
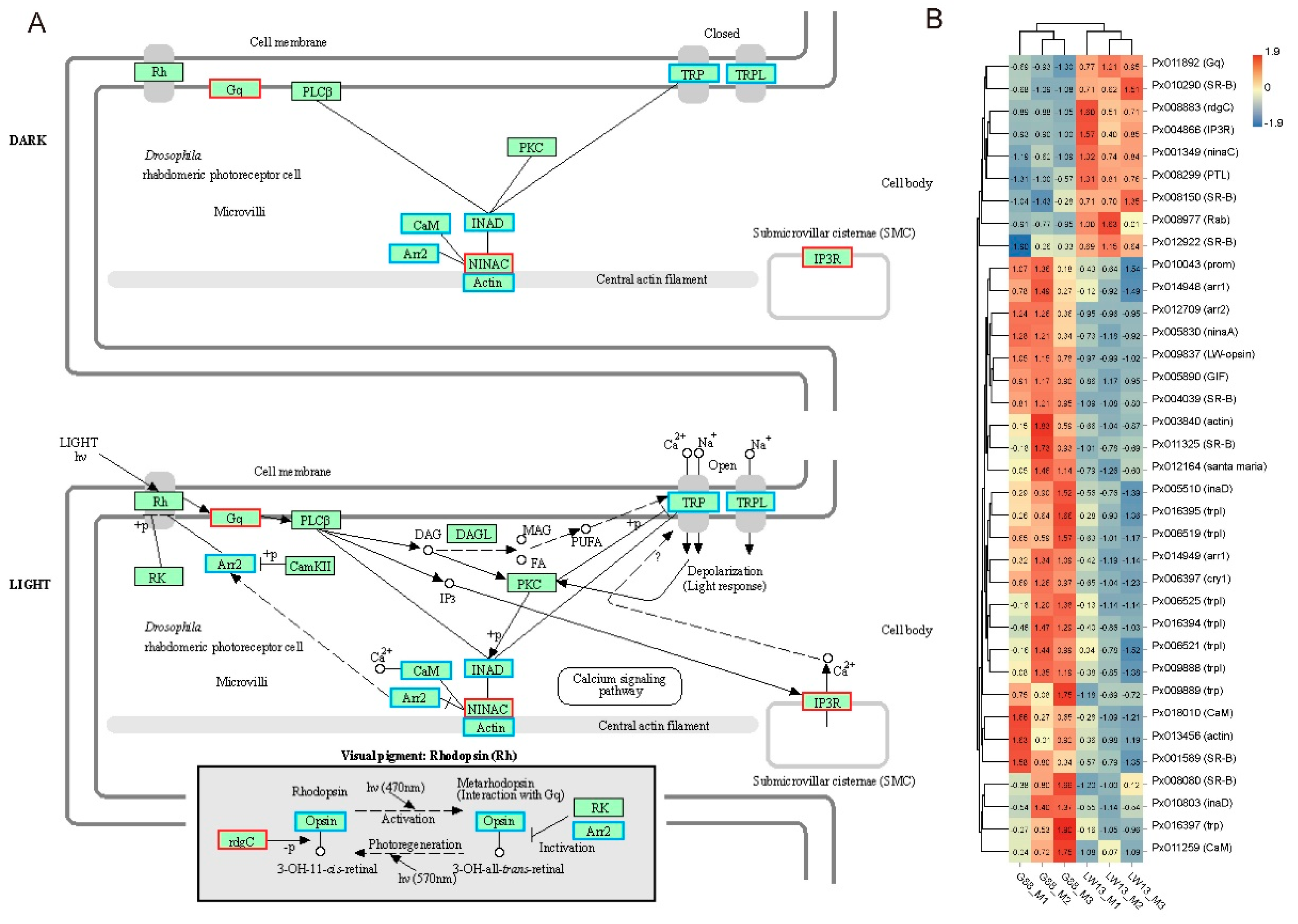
3.5. Influences of LW-Opsin Mutation on the Expression of Genes in the Phototransduction Pathway
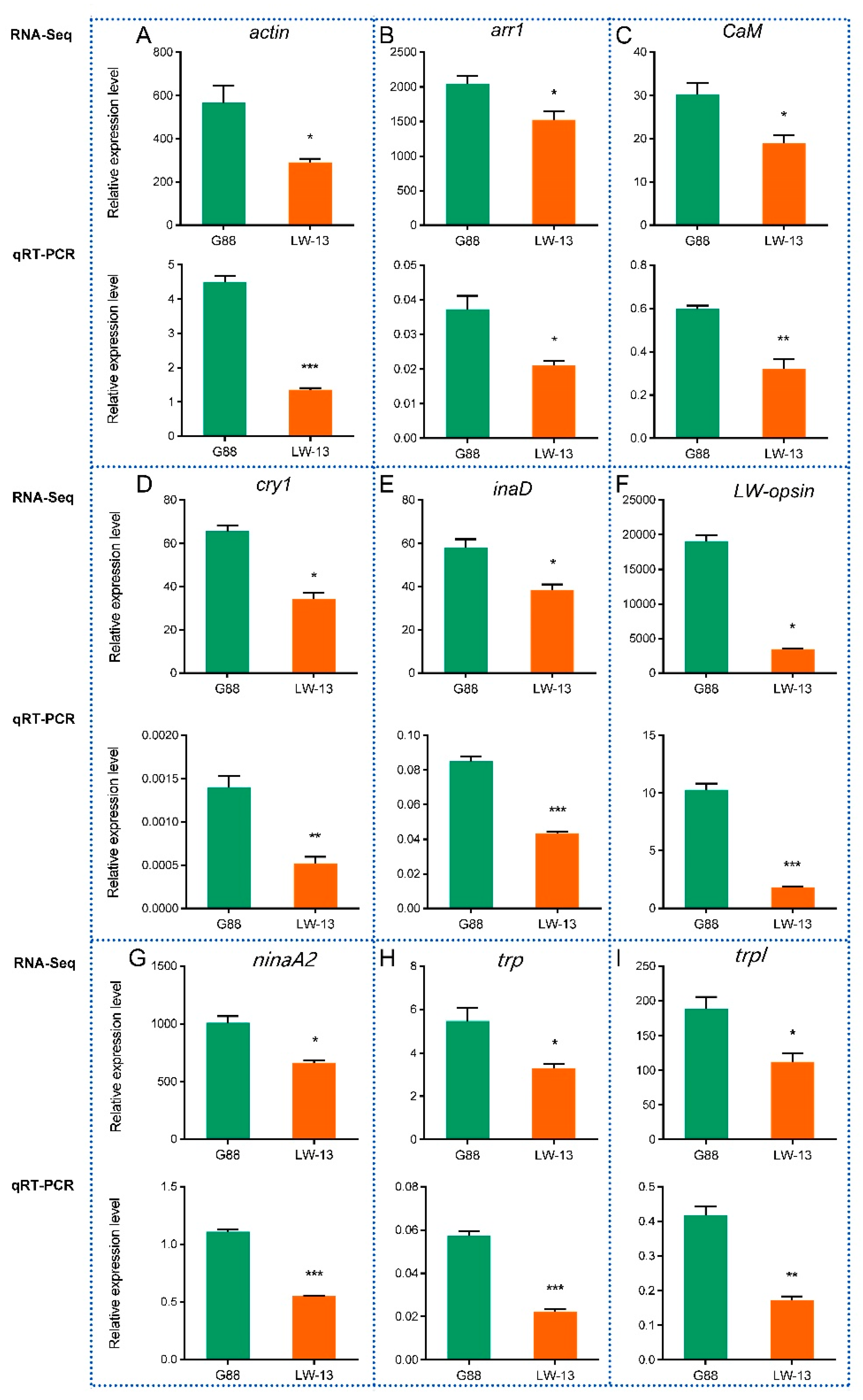
3.6. RT-qPCR Validation of DEGs in the Phototransduction Pathway
3.7. The Effects of cry1 Mutation on Phototactic Behavior of P. xylostella
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fain, G.L.; Hardie, R.; Laughlin, S.B. Phototransduction and the evolution of photoreceptors. Curr. Biol. 2010, 20, 114–124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hardie, R.C.; Juusola, M. Phototransduction in Drosophila. Curr. Opin. Neurobiol. 2015, 34, 37–45. [Google Scholar] [CrossRef] [Green Version]
- Katz, B.; Minke, B. The Drosophila light-activated TRP and TRPL channels—Targets of the phosphoinositide signaling cascade. Prog. Retin. Eye Res. 2018, 66, 200–219. [Google Scholar] [CrossRef] [PubMed]
- Landry, C.R.; Castillo-Davis, C.I.; Ogura, A.; Liu, J.S.; Hartl, D.L. Systems-level analysis and evolution of the phototransduction network in Drosophila. Proc. Natl. Acad. Sci. USA 2007, 104, 3283–3288. [Google Scholar] [CrossRef] [Green Version]
- Montell, C. Drosophila visual transduction. Trends Neurosci. 2012, 35, 356–363. [Google Scholar] [CrossRef] [Green Version]
- Paulsen, R.; Schwemer, J. Biogenesis of blowfly photoreceptor membranes is regulated by 11-cis-retinal. Eur. J. Biochem. 1983, 137, 609–614. [Google Scholar] [CrossRef] [PubMed]
- Yau, K.W.; Hardie, R.C. Phototransduction motifs and variations. Cell 2009, 139, 246–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Friedrich, M.; Chen, R.; Daines, B.; Bao, R.; Caravas, J.; Rai, P.K.; Zagmajster, M.; Peck, S.B. Phototransduction and clock gene expression in the troglobiont beetle Ptomaphagus hirtus of Mammoth cave. J. Exp. Biol. 2011, 214, 3532–3541. [Google Scholar] [CrossRef] [Green Version]
- Macias-Munoz, A.; Smith, G.; Monteiro, A.; Briscoe, A.D. Transcriptome-wide differential gene expression in Bicyclus anynana butterflies: Female vision-related genes are more plastic. Mol. Biol. Evol. 2016, 33, 79–92. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.; Gong, Z.J.; Wu, R.H.; Miao, J.; Jiang, Y.L.; Li, T.; Wu, X.B.; Wu, Y.Q. Transcriptome analysis of molecular mechanisms responsible for light-stress response in Mythimna separata (Walker). Sci. Rep. 2017, 7, 45188. [Google Scholar] [CrossRef]
- Macias-Muñoz, A.; Rangel Olguin, A.G.; Briscoe, A.D.; Li, W.H. Evolution of phototransduction genes in lepidoptera. Genome Biol. Evol. 2019, 11, 2107–2124. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feuda, R.; Marletaz, F.; Bentley, M.A.; Holland, P.W.H. Conservation, duplication, and divergence of five opsin genes in insect evolution. Genome Biol. Evol. 2016, 8, 579–587. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yamaguchi, S.; Desplan, C.; Heisenberg, M. Contribution of photoreceptor subtypes to spectral wavelength preference in Drosophila. Proc. Natl. Acad. Sci. USA 2010, 107, 5634–5639. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wakakuwa, M.; Stewart, F.; Matsumoto, Y.; Matsunaga, S.; Arikawa, K. Physiological basis of phototaxis to near-infrared light in Nephotettix cincticeps. J. Comp. Physiol. A 2014, 200, 527–536. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.J.; Yan, S.; Shen, Z.J.; Li, Z.; Zhang, X.F.; Liu, X.M.; Zhang, Q.W.; Liu, X.X. The expression of three opsin genes and phototactic behavior of Spodoptera exigua (Lepidoptera: Noctuidae): Evidence for visual function of opsin in phototaxis. Insect Biochem. Mol. Biol. 2018, 96, 27–35. [Google Scholar] [CrossRef] [PubMed]
- Li, C.; Tian, F.; Lin, T.; Wang, Z.; Liu, J.; Zeng, X. The expression and function of opsin genes related to the phototactic behavior of Asian citrus psyllid. Pest Manag. Sci. 2020, 76, 1578–1587. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.P.; Liu, Z.X.; Chen, Y.T.; Wang, Y.; Chen, J.Z.; Fu, S.; Ma, W.F.; Xia, S.; Liu, D.; Wu, T.; et al. CRISPR/Cas9-mediated knockout of LW-opsin reduces the efficiency of phototaxis in the diamondback moth Plutella xylostella. Pest Manag. Sci. 2021, 77, 3519–3528. [Google Scholar] [CrossRef]
- Chen, S.P.; Wang, D.F.; Ma, W.F.; Lin, X.L.; Yang, G. Knockout of cryptochrome1 disturbs the locomotor circadian rhythm and development of Plutella xylostella. Insect Sci. 2022. [Google Scholar] [CrossRef]
- Ma, X.L.; He, W.Y.; Wang, P.; You, M.S. Cell lines from diamondback moth exhibiting differential susceptibility to baculovirus infection and expressing midgut genes. Insect Sci. 2019, 26, 251–262. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Baik, L.S.; Fogle, K.J.; Roberts, L.; Galschiodt, A.M.; Chevez, J.A.; Recinos, Y.; Nguy, V.; Holmes, T.C. CRYPTOCHROME mediates behavioral executive choice in response to UV light. Proc. Natl. Acad. Sci. USA 2017, 114, 776–781. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bouly, J.P.; Schleicher, E.; Dionisio-Sese, M.; Vandenbussche, F.; Van Der Straeten, D.; Bakrim, N.; Meier, S.; Batschauer, A.; Galland, P.; Bittl, R.; et al. Cryptochrome blue light photoreceptors are activated through interconversion of flavin redox states. J. Biol. Chem. 2007, 282, 9383–9391. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- You, M.; Yue, Z.; He, W.; Yang, X.; Yang, G.; Xie, M.; Zhan, D.; Baxter, S.W.; Vasseur, L.; Gurr, G.M.; et al. A heterozygous moth genome provides insights into herbivory and detoxification. Nat. Genet. 2013, 45, 220–225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colley, N.J.; Cassill, J.A.; Baker, E.K.; Zuker, C.S. Defective intracellular transport is the molecular basis of rhodopsin-dependent dominant retinal degeneration. Proc. Natl. Acad. Sci. USA 1995, 92, 3070–3074. [Google Scholar] [CrossRef] [Green Version]
- Kumar, J.P.; Ready, D.F. Rhodopsin plays an essential structural role in Drosophila photoreceptor development. Development 1995, 121, 4359–4370. [Google Scholar] [CrossRef]
- Wang, T.; Montell, C. Phototransduction and retinal degeneration in Drosophila. Pflug. Arch. Eur. J. Physiol. 2007, 454, 821–847. [Google Scholar] [CrossRef] [Green Version]
- Zanini, D.; Giraldo, D.; Warren, B.; Katana, R.; Andrés, M.; Reddy, S.; Pauls, S.; Schwedhelm-Domeyer, N.; Geurten, B.R.H.; Göpfert, M.C. Proprioceptive opsin functions in Drosophila larval locomotion. Neuron 2018, 98, 67–74. [Google Scholar] [CrossRef] [Green Version]
- Scott, K.; Becker, A.; Sun, Y.; Hardy, R.; Zuker, C. Gqα protein function in vivo: Genetic dissection of its role in photoreceptor cell physiology. Neuron 1995, 15, 919–927. [Google Scholar] [CrossRef] [Green Version]
- Dolph, P.J.; Ranganathan, R.; Colley, N.J.; Hardy, R.W.; Socolich, M.; Zuker, C.S. Arrestin function in inactivation of G protein-coupled receptor rhodopsin in vivo. Science 1993, 260, 1910–1916. [Google Scholar] [CrossRef]
- Chevesich, J.; Kreuz, A.J.; Montell, C. Requirement for the PDZ domain protein, INAD, for localization of the TRP store-operated channel to a signaling complex. Neuron 1997, 18, 95–105. [Google Scholar] [CrossRef]
- Li, H.S.; Montell, C. TRP and the PDZ protein, INAD, form the core complex required for retention of the signalplex in Drosophila photoreceptor cells. J. Cell Biol. 2000, 150, 1411–1421. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wes, P.D.; Xu, X.Z.S.; Li, H.S.; Chien, F.; Doberstein, S.K.; Montell, C. Termination of phototransduction requires binding of the NINAC myosin III and the PDZ protein INAD. Nat. Neurosci. 1999, 2, 447–453. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.Z.S.; Choudhury, A.; Li, X.; Montell, C. Coordination of an array of signaling proteins through homo- and heteromeric interactions between PDZ domains and target proteins. J. Cell Biol. 1998, 142, 545–555. [Google Scholar] [CrossRef] [PubMed]
- Hardie, R.C.; Minke, B. The trp gene is essential for a light-activated Ca2+ channel in Drosophila photoreceptors. Neuron 1992, 8, 643–651. [Google Scholar] [CrossRef]
- Leung, H.T.; Geng, C.; Pak, W.L. Phenotypes of trpl mutants and interactions between the transient receptor potential (TRP) and TRP-like channels in Drosophila. J. Neurosci. 2000, 20, 6797–6803. [Google Scholar] [CrossRef] [Green Version]
- Dewett, D.; Lam-Kamath, K.; Poupault, C.; Khurana, H.; Rister, J. Mechanisms of vitamin A metabolism and deficiency in the mammalian and fly visual system. Dev. Biol. 2021, 476, 68–78. [Google Scholar] [CrossRef]
- Dewett, D.; Labaf, M.; Lam-Kamath, K.; Zarringhalam, K.; Rister, J. Vitamin A deficiency affects gene expression in the Drosophila melanogaster head. G3 Genes Genomes Genet. 2021, 11, jkab297. [Google Scholar] [CrossRef]
- Harris, W.; Ready, D.; Lipson, E.; Hudspeth, A.; Stark, W. Vitamin A deprivation and Drosophila photopigments. Nature 1977, 266, 648–650. [Google Scholar] [CrossRef]
- Wang, T.; Jiao, Y.; Montell, C. Dissection of the pathway required for generation of vitamin A and for Drosophila phototransduction. J. Cell Biol. 2007, 177, 305–316. [Google Scholar] [CrossRef] [Green Version]
- Lane, M.A.; Bailey, S.J. Role of retinoid signalling in the adult brain. Prog. Neurobiol. 2005, 75, 275–293. [Google Scholar] [CrossRef]
- Travis, G.H.; Golczak, M.; Moise, A.R.; Palczewski, K. Diseases caused by defects in the visual cycle: Retinoids as potential therapeutic agents. Annu. Rev. Pharmacol. Toxicol. 2007, 47, 469–512. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baik, L.S.; Recinos, Y.; Chevez, J.A.; Au, D.D.; Holmes, T.C. Multiple phototransduction inputs integrate to mediate UV light-evoked avoidance/attraction behavior in Drosophila. J. Biol. Rhythm. 2019, 34, 391–400. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Yang, G.; Chen, W. Diel and circadian patterns of locomotor activity in the adults of diamondback moth (Plutella xylostella). Insects 2021, 12, 727. [Google Scholar] [CrossRef] [PubMed]
- Mazzotta, G.; Rossi, A.; Leonardi, E.; Mason, M.; Bertolucci, C.; Caccin, L.; Spolaore, B.; Martin, A.J.M.; Schlichting, M.; Grebler, R.; et al. Fly cryptochrome and the visual system. Proc. Natl. Acad. Sci. USA 2013, 110, 6163–6168. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Q30 (%) | Total Mapped | Multiple Mapped | Uniquely Mapped |
|---|---|---|---|---|
| G88 M1 | 94.61 | 23,206,466 (55.63%) | 1,350,166 (3.24%) | 21,856,300 (52.39%) |
| G88 M2 | 94.67 | 25,289,426 (55.93%) | 1,448,410 (3.20%) | 23,841,016 (52.72%) |
| G88 M3 | 94.86 | 25,923,219 (56.01%) | 1,521,146 (3.29%) | 24,402,073 (52.72%) |
| LW-13 M1 | 94.67 | 22,971,282 (54.15%) | 1,367,482 (3.22%) | 21,603,800 (50.92%) |
| LW-13 M2 | 94.81 | 23,454,846 (54.49%) | 1,393,748 (3.24%) | 22,061,098 (51.25%) |
| LW-13 M3 | 94.76 | 26,024,544 (54.10%) | 1,496,406 (3.11%) | 24,528,138 (50.99%) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chen, S.-P.; Lin, X.-L.; Qiu, R.-Z.; Chi, M.-X.; Yang, G. An LW-Opsin Mutation Changes the Gene Expression of the Phototransduction Pathway: A Cryptochrome1 Mutation Enhances the Phototaxis of Male Plutella xylostella (Lepidoptera: Plutellidae). Insects 2023, 14, 72. https://doi.org/10.3390/insects14010072
Chen S-P, Lin X-L, Qiu R-Z, Chi M-X, Yang G. An LW-Opsin Mutation Changes the Gene Expression of the Phototransduction Pathway: A Cryptochrome1 Mutation Enhances the Phototaxis of Male Plutella xylostella (Lepidoptera: Plutellidae). Insects. 2023; 14(1):72. https://doi.org/10.3390/insects14010072
Chicago/Turabian StyleChen, Shao-Ping, Xiao-Lu Lin, Rong-Zhou Qiu, Mei-Xiang Chi, and Guang Yang. 2023. "An LW-Opsin Mutation Changes the Gene Expression of the Phototransduction Pathway: A Cryptochrome1 Mutation Enhances the Phototaxis of Male Plutella xylostella (Lepidoptera: Plutellidae)" Insects 14, no. 1: 72. https://doi.org/10.3390/insects14010072