
Review of the Neotropical Species of the Elachista praelineata Species Group (Lepidoptera, Elachistidae, Elachistinae) with Identification Keys and Description of a New Species from Bolivia

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Morphological Study
2.2. DNA Extraction, Amplification, Sequencing and Data Analysis
3. Results
3.1. Taxonomy
- Valva straight, not curved towards cucullus ([9]: Figure 24) ……………………………………………………………………………………………………………… E. lata
- -
- Valva strongly curved and broadened towards cucullus …………………………………………………………………………………………………………………… 2
- 2.
- Sclerotized phallic tube straight ([8]: Figure 14B) ………………………………………………………………………………………………………………… E. albisquamella
- -
- Sclerotized phallic tube curved …………………………………………………………………………………………………………………………………………………… 3
- 3.
- Sclerotized phallic tube gradually bent, s-shaped, apex blunt ([9]: Figures 11, 15 and 16) ……………………………………………………………………… E. adunca
- -
- Sclerotized phallic tube curved at basal 1/2 and 4/5, apex tapered ([9]: Figures 18 and 21) ……………………………………………………………………………… E. laxa
- Corpus bursae densely covered with distinct internal spines (this paper, Figure 1E) ……………………………………………………………………………… E. albisquamella
- -
- Corpus bursae without internal spines …………………………………………………………………………………………………………………………………………………… 2
- 2.
- Ostium bursae occupying 1/2 width between apophyses anteriores; signum angulate ([9]: Figures 38 and 40) ………………………………………………………… E. lata
- -
- Ostium bursae occupying 3/5 width between apophyses anteriores; signum straight (this paper, Figure 1D,E) ……………………………………………… E. stonisi sp. nov.

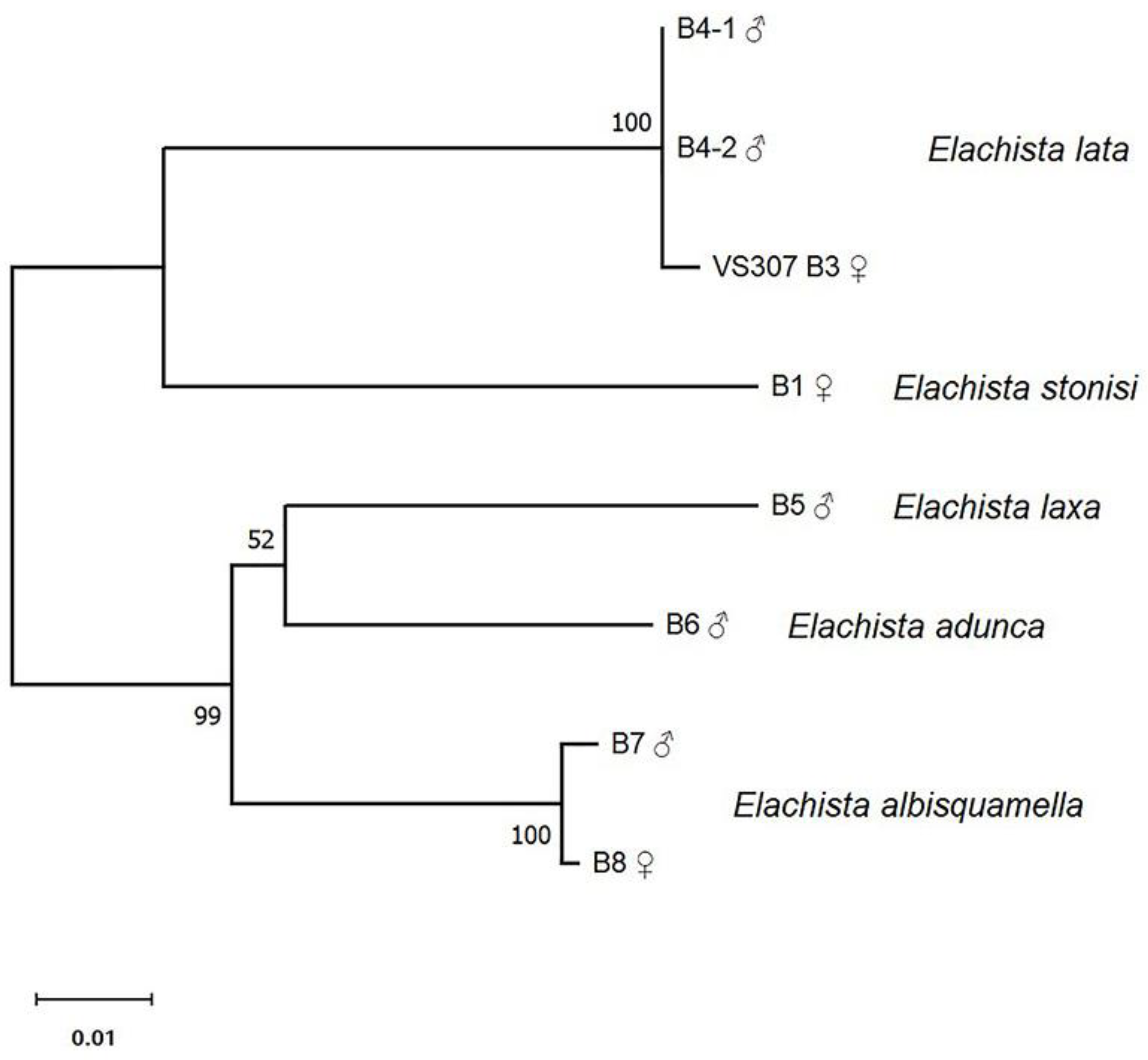
3.2. COI Fragment Analysis
4. Discussion
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kaila, L. Elachistine moths of Australia (Lepidoptera: Gelechioidea: Elachistidae). Monographs on Australian Lepidoptera; CSIRO Publishing: Melbourne, Australia, 2011; Volume 11, 443p. [Google Scholar]
- Traugott-Olsen, E.; Nielsen, E.S. The Elachistidae (Lepidoptera) of Fennoscandia and Denmark. Fauna Entomol. Scand. 1977, 6, 1–299. [Google Scholar]
- Parenti, U.; Varalda, P.J. Gli Elachistidi (Lepidoptera, Elachistidae) e le loro piante ospiti. Boll. Mus. Reg. Sci. Nat. Torino 1994, 12, 73–136. [Google Scholar]
- Kaila, L. An annotated catalogue of Elachistinae of the World (Lepidoptera: Gelechioidea: Elachistidae). Zootaxa 2019, 4632, 1–231. [Google Scholar] [CrossRef] [PubMed]
- Kaila, L. Phylogeny and classification of the Elachistidae s.s. (Lepidoptera: Gelechioidea). Syst. Entomol. 1999, 24, 139–169. [Google Scholar] [CrossRef]
- Kaila, L.; Sugisima, K. 1. Phylogeny, subfamily definition and generic classification. In Elachistine moths of Australia (Lepidoptera: Gelechioidea: Elachistidae). Monographs on Australian Lepidoptera; Kaila, L., Ed.; CSIRO Publishing: Melbourne, Australia, 2011; Volume 11, pp. 7–22. [Google Scholar]
- Kaila, L. A revision of the Nearctic species of the genus Elachista s. l. III. The bifasciella, praelineata, saccharella and freyerella groups (Lepidoptera, Elachistidae). Acta Zool. Fenn. 1999, 211, 1–235. [Google Scholar]
- Kaila, L. A review of the South American Elachistidae s. str. (Lepidoptera, Gelechioidea), with descriptions of 15 new species. Steenstrupia 2000, 25, 159–193. [Google Scholar]
- Sruoga, V. The Elachistinae (Lepidoptera: Gelechioidea: Elachistidae) of Ecuador with descriptions of five new species. Zootaxa 2010, 2524, 33–50. [Google Scholar] [CrossRef]
- Zeller, P.C. Exotische Microlepidoptera. Hor. Ent. Ross. 1877, 13, 447–448. [Google Scholar]
- Braun, A.F. Life histories of North American Tineina. Can. Entomol. 1915, 47, 104–108. [Google Scholar] [CrossRef]
- Braun, A.F. Elachistidae of North America (Microlepidoptera). Mem. Am. Entomol. Soc. 1948, 13, 1–110. [Google Scholar]
- Forbes, W.T.M. The Lepidoptera of New York and Neighboring States. Primitive Forms, Microlepidoptera, Pyraloids, Bombyces. Cornell University. Agricultural Experiment Station. Memoir 68; Cornell University: Ithaca, NY, USA, 1923; p. 222. [Google Scholar]
- Braun, A.F. Microlepidoptera: Notes and new species. Can. Entomol. 1922, 54, 90–94. [Google Scholar] [CrossRef] [Green Version]
- Chambers, V.T. Descriptions of new Tineina from Texas, and others from more northern localities. Bull. U. S. Geol. Geogr. Surv. Terr. 1878, 4, 79–106. [Google Scholar]
- Parenti, U. Elachistidi del Giappone (Lepidoptera, Elachistidae). Boll. Mus. Reg. Sci. Nat. Torino 1983, 1, 1–20. [Google Scholar]
- Sugisima, K. A revision of the Elachista praelineata-group (Lepidoptera, Elachistidae) in Japan, with comments on morphology of the pupa in Elachista. Tijdschr. Entomol. 2005, 148, 1–19. [Google Scholar] [CrossRef]
- Sruoga, V.; Kaila, L.; Rocienė, A. The Elachistinae (Lepidoptera: Gelechioidea, Elachistidae) of Thailand, with description of eight new species. Eur. J. Taxon. 2019, 574, 1–34. [Google Scholar] [CrossRef]
- Chrétien, P. Contribution à la connaissance des Lépidoptères du Nord de l’Afrique. Ann. Soc. Entomol. Fr. 1915, 84, 289–374. [Google Scholar]
- Parenti, U. Revisione degli Elachistidi (Lepidoptera, Elachistidae) paleartici. I. Tipi di Elachistidi del Museo di Storia naturale di Parigi. Boll. Mus. Zool. Univ. Torino 1972, 2, 29–56. [Google Scholar]
- Sinev, S.Y.; Sruoga, V.A. Family Elachistidae (Lepidoptera). In Key to the Insects of Russian Far East. Vol. V. Trichoptera and Lepidoptera. Part. 1; Ler, P.A., Ed.; Dal’nauka: Vladivostok, Russia, 1997; Volume 5, pp. 491–502. (In Russian) [Google Scholar]
- Nielsen, E.S.; Traugott-Olsen, E. Four new West Palaearctic species of Elachistidae (Lepidoptera). Entomol. Gaz. 1987, 38, 103–113. [Google Scholar]
- Klimesch, J. Beitraege zur Kenntnis der Microlepidopterenfauna des Kanarischen Archipels. 10. Beitraeg: Elachistidae. Vieraea 1990, 19, 185–192. [Google Scholar]
- Kaila, L.; Karsholt, O. Contribution to the Lepidoptera fauna of the Madeira island Part 3. Elachistidae. Beitr. Ent. 2002, 52, 225–233. [Google Scholar]
- Koo, J.M.; Kim, J.D.; Kim, D.S.; Park, K.T. Three Species of Microlepidoptera (Lepidoptera) New to Korea. Anim. Syst. Evol. Divers. 2021, 37, 96–100. [Google Scholar] [CrossRef]
- Sruoga, V. New species and records of Elachistinae (Lepidoptera: Gelechioidea, Elachistidae) from Nepal. Zootaxa 2022, 5100, 573–584. [Google Scholar] [CrossRef] [PubMed]
- Traugott-Olsen, E. Elachista nepalensis Traugott-Olsen, sp. n., new species from Nepal (Lepidoptera: Elachistidae). SHILAP Revta. lepid. 1999, 27, 419–422. [Google Scholar]
- Parenti, U. Nuove specie paleartiche del Genere Elachista Treitschke (Lepidoptera, Elachistidae). Boll. Mus. Zool. Univ. Torino 1978, 4, 15–26. [Google Scholar]
- Parenti, U.; Varalda, P.G. Elachista littoricola Le Marchand ed Elachista sicula Parenti: Note biologiche. (Lepidoptera, Elachistidae). Boll. Mus. Reg. Sci. Nat. Torino 1991, 9, 427–435. [Google Scholar]
- Amsel, H.G. Neue palästinensische Lepidopteren. Mitt. Zool. Mus. Berl. 1935, 20, 271–319. [Google Scholar]
- Parenti, U. Revisioni degli Elachistidi (Lepidoptera, Elachistidae) paleartici. III. Le specie di Elachistidi descritte da HG Amsel e F. Hartig. Boll. Mus. Zool. Univ. Torino 1973, 3, 41–58. [Google Scholar]
- Sruoga, V.; De Prins, J. The Elachistinae (Lepidoptera: Elachistidae) of Kenya with descriptions of eight new species. Zootaxa 2009, 2172, 1–31. [Google Scholar] [CrossRef]
- Meyrick, E. Descriptions of South-African Microlepidoptera. Ann. S. Afr. Mus. 1920, 17, 273–318. [Google Scholar] [CrossRef]
- Parenti, U. About some African and Asiatic species of the family Elachistidae (Lepidoptera) described by E. Meyrick. Stapfia 1988, 16, 185–198. [Google Scholar]
- Meyrick, E. Exotic Microlepidoptera, 2nd ed.; Taylor and Francis: London, UK, 1914; Volume 1, pp. 257–288. [Google Scholar] [CrossRef]
- Sruoga, V. Re-descriptions of two Meyrick’s types of Elachista (Lepidoptera, Elachistidae) from the Sub-Saharan Africa. Dtsch. Entomol. Z. 2002, 49, 137–141. [Google Scholar] [CrossRef]
- Wollaston, T.V. Notes on the Lepidoptera of St. Helena, with descriptions of new species. Ann. Mag. Nat. Hist. Ser. V 1879, 3, 415–441. [Google Scholar] [CrossRef] [Green Version]
- Sinev, S.Y. World Catalogue of cosmopterigid moths (Lepidoptera: Cosmopterigidae). Proc. ZIN 2002, 293, 1–184. [Google Scholar]
- Fowler, L.; Karisch, T. Elachista trifasciata (Wollaston, 1879) on St Helena Island (Lepidoptera, Elachistidae, Elachistinae). Metamorphosis 2020, 31, 28–32. [Google Scholar] [CrossRef]
- Meyrick, E. Descriptions of South African Micro-Lepidoptera. Ann. Transvaal Mus. 1921, 8, 49–148. [Google Scholar] [CrossRef]
- Sruoga, V. Redescriptions of two of Meyrick’s types of Elachistidae (Lepidoptera: Gelechioidea) from India and Sri Lanka. Acta Zool. Litu. 2005, 15, 58–61. [Google Scholar] [CrossRef]
- Robinson, G.S. The preparation of slides of Lepidoptera genitalia with special reference to the Microlepidoptera. Entomol. Gaz. 1976, 27, 127–132. [Google Scholar]
- Kristensen, N.P. (Ed.) Skeleton and muscles: Adults. In Lepidoptera, Moths and Butterflies. Vol. 2. Morphology, Physiology, and Development; Walter de Gruyter: Berlin, Germany; New York, NY, USA, 2003; pp. 39–122. [Google Scholar] [CrossRef]
- Hajibabaei, M.; Janzen, D.H.; Burns, J.M.; Hallwachs, W.; Hebert, P.D.N. DNA barcodes distinguish species of tropical Lepidoptera. Proc. Natl. Acad. Sci. USA 2006, 103, 968–971. [Google Scholar] [CrossRef] [Green Version]
- Wahlberg, N.; Wheat, C.W. Genomic outposts serve the phylogenomic pioneers: Designing novel nuclear markers for genomic DNA extractions of Lepidoptera. Syst. Biol. 2008, 57, 231–242. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Tamura, K.; Stecher, G.; Kumar, S. MEGA11: Molecular Evolutionary Genetics Analysis Version 11. Mol. Biol. Evol. 2021, 38, 3022–3027. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Sanchez, E.; Tyrrell, C.D.; Londono, X.; Oliveira, R.P.; Clark, L.G. Diversity, distribution, and classification of Neotropical woody bamboos (Poaceae: Bambusoideae) in the 21st Century. Bot. Sci. 2021, 99, 198–228. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Elachista Species | Host Plant | Distribution | References |
|---|---|---|---|
| Neotropical Region | |||
| adunca Sruoga, 2010 | Unknown | Ecuador | [9] |
| albisquamella Zeller, 1877 | Bambusoideae sp. | Colombia; Bolivia | [8,10], present study |
| lata Sruoga, 2010 | Unknown | Ecuador | [9] |
| laxa Sruoga, 2010 | Unknown | Ecuador | [9] |
| stonisi sp. nov. | Poaceae sp. | Bolivia | Present study |
| Nearctic Region | |||
| aranella Kaila, 1999 | Unknown | Canada | [7] |
| aristoteliella Kaila, 1999 | Unknown | USA | [7] |
| eilinella Kaila, 1999 | Unknown | USA | [7] |
| guilinella Kaila, 1999 | Unknown | USA | [7] |
| ibunella Kaila, 1999 | Unknown | USA | [7] |
| miriella Kaila, 1999 | Unknown | USA | [7] |
| nienorella Kaila, 1999 | Unknown | USA | [7] |
| praelineata Braun, 1915 | Hystrix patula Moench | USA | [7,11,12,13] |
| radiantella Braun, 1922 | Panicum clandestinum (L.), P. dichotomum (L.) | USA | [7,12,14] |
| serindella Kaila, 1999 | Unknown | USA | [7] |
| solitaria Braun, 1922 | Panicum sp. | USA | [7,12,14] |
| staintonella Chambers, 1878 | Unknown | USA | [7,12,15] |
| turinella Kaila, 1999 | Unknown | USA | [7] |
| Palearctic Region | |||
| amamii Parenti, 1983 | Digitaria timorensis (Kunth) Balansa, D. adscendens (H. B. K.) Henr., Thuerea involuta (Forst.) R. Br. | Japan; Taiwan; Thailand | [16,17,18] |
| bromella Chrétien, 1915 | Bromus madritensis L. | Algeria; Morocco | [4,19,20] |
| caliginosa Parenti, 1983 | Eccoilopus cotulifer A. Camus, Spodiopogon sibiricus Trin. | Russia; Japan | [16,17,21] |
| canariella Nielsen & Traugott-Olsen, 1987 | Bromus sp. | Canary Islands | [22,23] |
| encumeadae Kaila & Karsholt, 2002 | Festuca donax Lowe | Madeira | [24] |
| fasciocaliginosa Sugisima, 2005 | Miscanthus sinensis Anderss. | Japan | [17] |
| kurokoi Parenti, 1983 | Oplismenus undatifolius (Ard.) P. Beauv. | Japan; Korea; Nepal | [16,17,25,26] |
| minor Sruoga, 2022 | Unknown | Nepal | [26] |
| miscanthi Parenti, 1983 | Miscanthus sinensis Anderss. | Japan; Taiwan | [16,17] |
| nepalensis Traugott-Olsen, 1999 | Unknown | Nepal | [27] |
| sicula Parenti, 1978 | Briza maxima L., Bromus erectus Hudson, Lagurus ovatus L. | Italy | [28,29] |
| simulans Sruoga, 2022 | Unknown | Nepal | [26] |
| tabghaella Amsel, 1935 | Unknown | Israel | [30,31] |
| Afrotropical Region | |||
| kakamegensis Sruoga & De Prins, 2009 | Unknown | Kenya | [32] |
| merimnaea Meyrick, 1920 | Unknown | South Africa | [33,34] |
| semophanta Meyrick, 1914 | Unknown | Malawi | [35,36] |
| trifasciata (Wollaston, 1879) | Unknown | Saint-Helena | [37,38,39] |
| Oriental Region | |||
| brachyplectra Meyrick, 1921 | Unknown | Sri Lanka; Indonesia | [40,41] |
| Australasian Region | |||
| aurita Kaila, 2011 | Oplismenus sp. | Australia | [1] |
| Species | DNA Isolate | GenBank Accession/BOLD Process ID/Length (bp) |
|---|---|---|
| Elachista stonisi; female; paratype | B1 | OP955854/EPRAE001-23/685 |
| Elachista sp. VS307; female | B3 | OP955855/EPRAE002-23/318 |
| Elachista lata; male; holotype | B4-1 | OP955856/EPRAE003-23/318 |
| Elachista lata; male; holotype | B4-2 | OP955857/EPRAE004-23/318 |
| Elachista laxa; male; holotype | B5 | OP955858/EPRAE005-23/318 |
| Elachista adunca; male; holotype | B6 | OP955859/EPRAE006-23/318 |
| Elachista albisquamella; male | B7 | OP955860/EPRAE007-23/685 |
| Elachista albisquamella; female | B8 | OP955861/EPRAE008-23/685 |
| B1 | B3 | B4-1 | B4-2 | B5 | B6 | B7 | B8 | |
|---|---|---|---|---|---|---|---|---|
| E. stonisi; B1 | ||||||||
| Elachista sp. VS307; B3 | 9.75 | |||||||
| E. lata; B4-1 | 9.43 | 0.31 | ||||||
| E. lata; B4-2 | 9.43 | 0.31 | 0.00 | |||||
| E. laxa; B5 | 12.58 | 12.26 | 11.95 | 11.95 | ||||
| E. adunca; B6 | 11.95 | 11.64 | 11.32 | 11.32 | 7.23 | |||
| E. albisquamella; B7 | 11.68 | 11.01 | 10.69 | 10.69 | 8.18 | 5.97 | ||
| E. albisquamella; B8 | 11.39 | 10.69 | 10.38 | 10.38 | 8.18 | 6.29 | 0.44 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sruoga, V.; Havelka, J. Review of the Neotropical Species of the Elachista praelineata Species Group (Lepidoptera, Elachistidae, Elachistinae) with Identification Keys and Description of a New Species from Bolivia. Insects 2023, 14, 62. https://doi.org/10.3390/insects14010062
Sruoga V, Havelka J. Review of the Neotropical Species of the Elachista praelineata Species Group (Lepidoptera, Elachistidae, Elachistinae) with Identification Keys and Description of a New Species from Bolivia. Insects. 2023; 14(1):62. https://doi.org/10.3390/insects14010062
Chicago/Turabian StyleSruoga, Virginijus, and Jekaterina Havelka. 2023. "Review of the Neotropical Species of the Elachista praelineata Species Group (Lepidoptera, Elachistidae, Elachistinae) with Identification Keys and Description of a New Species from Bolivia" Insects 14, no. 1: 62. https://doi.org/10.3390/insects14010062