
Host Plant Variation and Lack of Genetic Differentiation in Populations of Dione (Agraulis) dodona Lamas & Farfán (Lepidoptera: Nymphalidae)
 , , ,
, , ,
Abstract
:Simple Summary
Abstract

1. Introduction
2. Materials and Methods
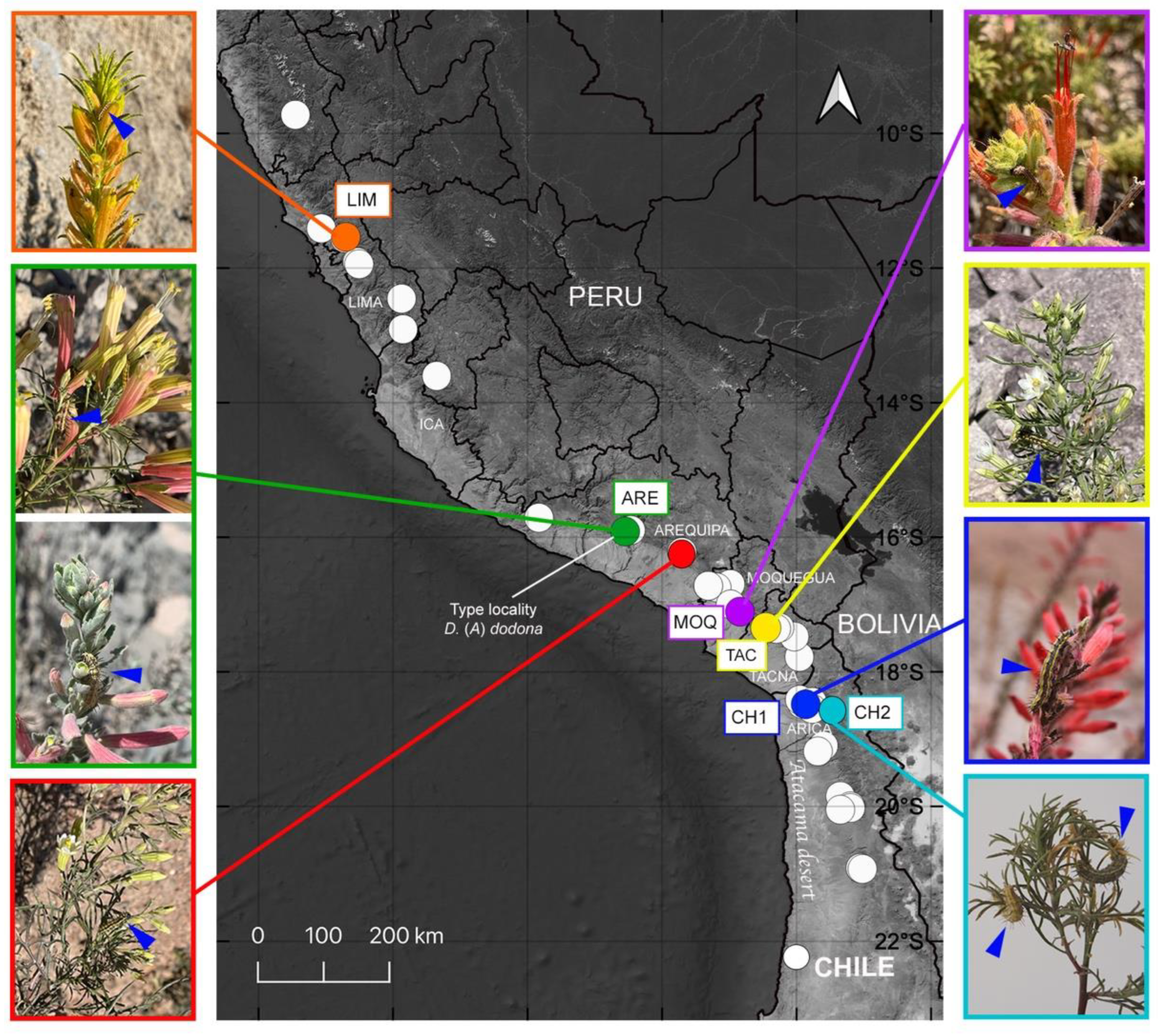
2.1. Sample Collection and Host-Plant Records
2.2. Genetic Analysis
2.2.1. DNA Extraction, Polymerase Chain Reaction Amplification, and Sequencing
2.2.2. Data Analysis
3. Results
3.1. Host-Plant Variation
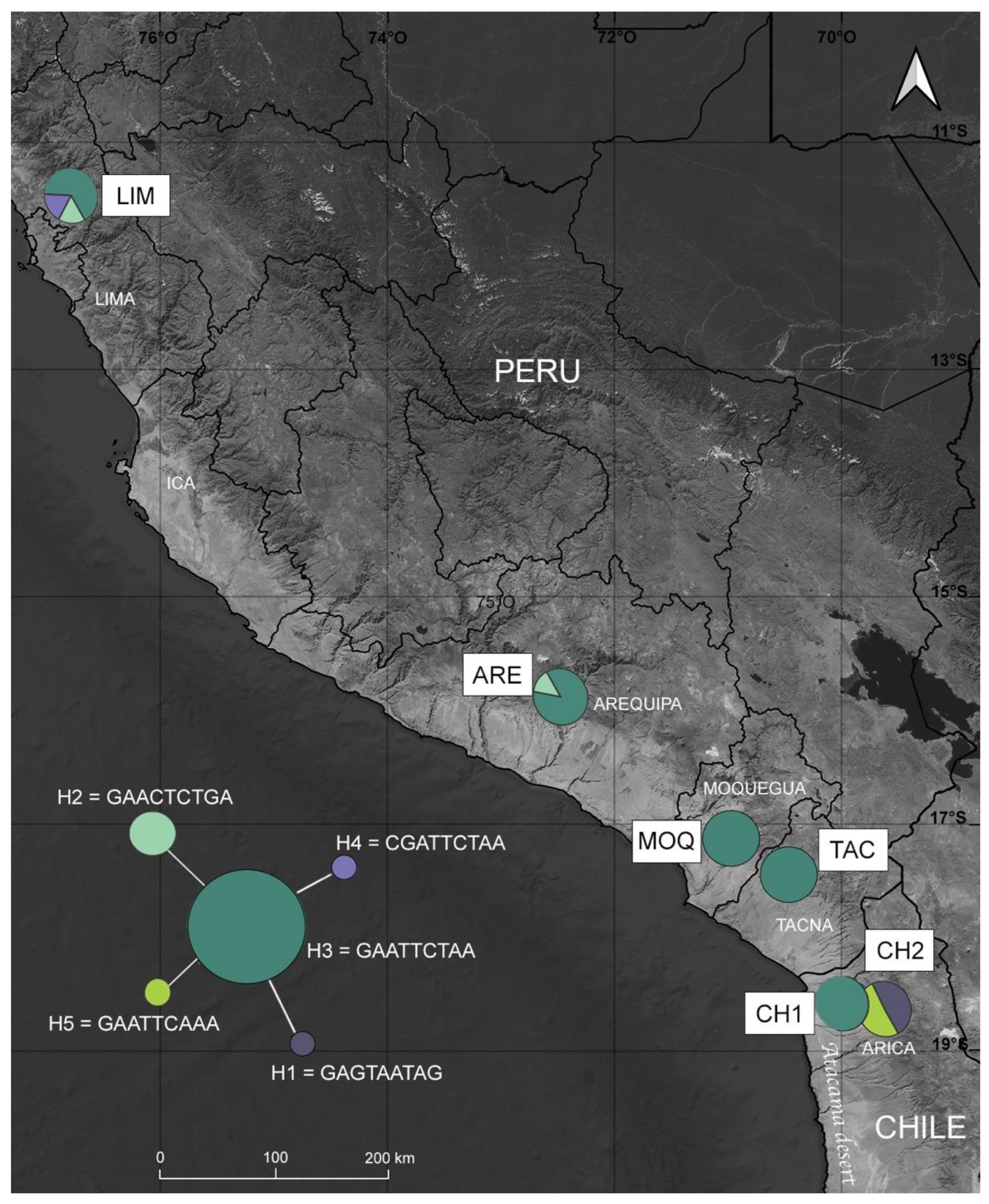
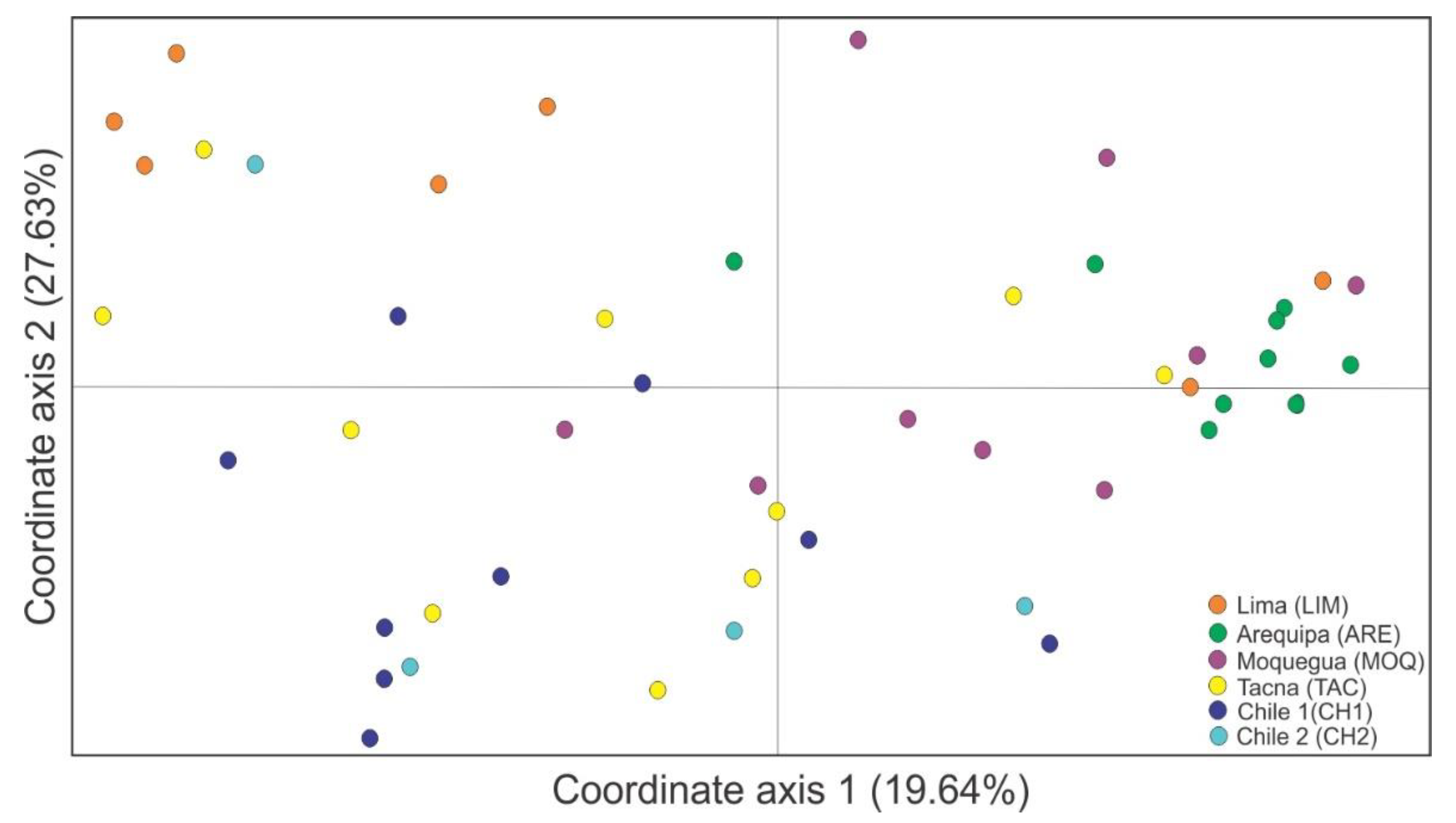
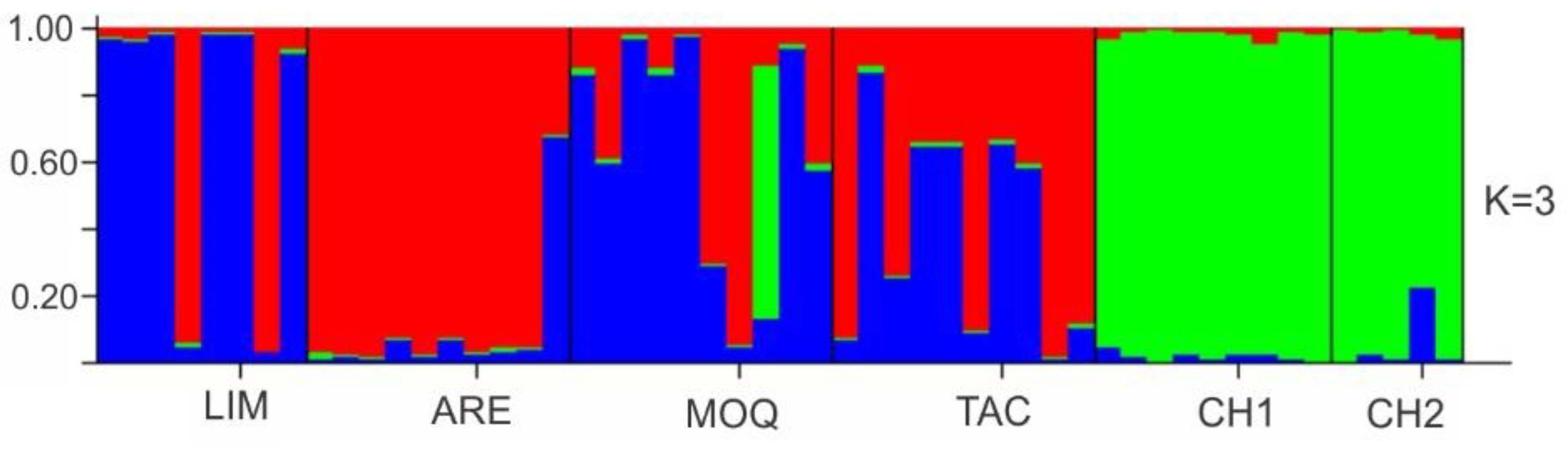
3.2. Genetic Differentiation
4. Discussion
4.1. Evolution of Heliconiinae–Passifloraceae in the Andes
4.2. Genetic Connectivity through Suitable Areas
4.3. Conservation of Suitable Areas
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Farfán, J.; Cerdeña, J.; Vargas, H.A.; Gonçalves, G.L.; Lamas, G.; Moreira, G.R.P. A peculiar new species of Dione (Agraulis) Boisduval & Le Conte (Lepidoptera: Nymphalidae: Heliconiinae) associated with Malesherbia Ruiz & Pavón (Passifloraceae) in xeric western slopes of the Andes. Zookeys 2022, 1113, 199–227. [Google Scholar]
- Gengler-Nowak, K.M. Reconstruction of the biogeographical history of Malesherbiaceae. Bot. Rev. 2002, 68, 171–188. [Google Scholar] [CrossRef]
- Weigend, M.; Jossberger, T.; Beltrán, H. Notes on Malesherbia (Passifloraceae) in Peru: A new species from southern Peru, a new record and a first report on interspecific hybridization in Malesherbia. Phytotaxa 2015, 202, 250–258. [Google Scholar] [CrossRef]
- Beltrán, H.; Roque, J.; Cáceres, C. Sinopsis del género Malesherbia (Passifloraceae) en el Perú. Rev. Peru. Biol. 2018, 25, 229–240. [Google Scholar] [CrossRef]
- Bull-Hereñu, K. The genus Malesherbia Ruiz & Pav. (Passifloraceae) in Chile. Phytotaxa 2020, 468, 1–44. [Google Scholar]
- Gengler-Nowak, K.M. Molecular phylogeny and taxonomy of Malesherbiaceae. Syst. Bot. 2003, 28, 333–344. [Google Scholar]
- Núñez, R.; Willmott, K.R.; Álvarez, Y.; Genaro, J.A.; Pérez-Asso, A.R.; Quejereta, M.; Turner, T.; Miller, J.Y.; Brévignon, C.; Lamas, G.; et al. Integrative taxonomy clarifies species limits in the hitherto monotypic passion-vine butterfly genera Agraulis and Dryas (Lepidoptera, Nymphalidae, Heliconiinae). Syst. Entomol. 2022, 47, 152–178. [Google Scholar] [CrossRef]
- Gagnaire, P.-A.; Broquet, T.; Aurelle, D.; Viard, F.; Souissi, A.; Bonhomme, F.; Arnaud-Haond, S.; Bierne, N. Using neutral, selected, and hitchhiker loci to assess connectivity of marine populations in the genomic era. Evol. Appl. 2015, 8, 769–786. [Google Scholar] [CrossRef]
- Lowe, A.J.; Harris, S.A.; Ashton, P. Ecological Genetics: Design, Analysis and Application, 1st ed.; Blackwells: Oxford, UK, 2004; p. 326. [Google Scholar]
- Allendorf, F.W. Genetics and the conservation of natural populations: Allozymes to genomes. Mol. Ecol. 2017, 26, 420–430. [Google Scholar] [CrossRef]
- Zhang, D.-X. Lepidopteran microsatellite DNA: Redundant but promising. Trends Ecol. Evol. 2004, 19, 507–509. [Google Scholar] [CrossRef]
- Flanagan, N.S.; Blum, M.J.; Davison, A.; Alamo, M.; Albarrán, R.; Faulhaber, K.; Peterson, E.; McMillan, W.O. Characterization of microsatellite loci in neotropical Heliconius butterflies. Mol. Ecol. Notes 2002, 2, 398–401. [Google Scholar] [CrossRef]
- Mavárez, J.; González, M. A set of microsatellite markers for Heliconius melpomene and closely related species. Mol. Ecol. Notes 2006, 6, 20–23. [Google Scholar] [CrossRef]
- Massardo, D.; Roratto, P.A.; Vargas, H.A.; Kronforst, M.R.; Moreira, G.R.P. Development of a microsatellite library for the passion flower butterfly Dione moneta Hübner (Lepidoptera: Nymphalidae: Heliconiinae). Conserv. Genet. Resour. 2012, 4, 719–724. [Google Scholar] [CrossRef]
- Peterson, M.A.; Denno, R.F. The influence of dispersal and diet breadth on patterns of genetic isolation by distance in phytophagous insects. Am. Nat. 1998, 152, 428–446. [Google Scholar] [CrossRef] [PubMed]
- Peterson, M.A.; Denno, R.F. Life history strategies and the genetic structure of phytophagous insect populations. In Genetic Structure and Local Adaptation in Natural Insect Populations: Effects of Ecology, Life History and Behavior, 1st ed.; Mopper, S., Strauss, S., Eds.; Chapman and Hall: New York, NY, USA, 1998; pp. 263–322. [Google Scholar]
- Drès, M.; Mallet, J. Host races in plant-feeding insects and their importance sympatric speciation. Philos. Trans. R. Soc. B Biol. Sci. 2002, 357, 471–492. [Google Scholar] [CrossRef]
- Huanca-Mamani, W.; Rivera-Cabello, D.; Maita-Maita, J. A simple, fast, and inexpensive CTAB-PVP-Silica based method for genomic DNA isolation from single, small insect larvae and pupae. Genet. Mol. Res. 2015, 14, 8001–8007. [Google Scholar] [CrossRef]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Massardo, D.; Fornel, R.; Kronforst, M.; Lopes-Goncalves, G.; Moreira, G.R.P. Diversification of the silverspot butterflies (Nymphalidae) in the Neotropics inferred from multi-locus DNA sequences. Mol. Phylogenet. Evol. 2015, 82, 156–165. [Google Scholar] [CrossRef]
- Nei, M. Estimation of average heterozygosity and genetic distance from a small number of individuals. Genetics 1978, 89, 583–590. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenALEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research-an update. Mol. Ecol. Notes 2002, 6, 288–295. [Google Scholar] [CrossRef]
- Van Oosterhout, C.; Hutchinson, W.F.; Wills, D.P.M.; Shipley, P. Micro-Checker: Software for identifying and correcting genotyping errors in microsatellite data. Mol. Ecol. Notes 2004, 4, 535–538. [Google Scholar] [CrossRef]
- Guo, S.W.; Thompson, E.A. Performing the Exact Test of Hardy-Weinberg Proportion for Multiple Alleles. Biometrics 1992, 48, 361–372. [Google Scholar] [CrossRef] [PubMed]
- Excoffier, L.; Schneiser, S. Arlequin version 3.0: An integrated software package for population genetics data analysis. Evol. Bioinform. Online 2005, 1, 47–50. [Google Scholar] [CrossRef]
- Rice, W.R. Analyzing tables of statistical tests. Evolution 1989, 43, 535–538. [Google Scholar] [CrossRef] [PubMed]
- Bandelt, H.J.; Forster, P.; Röhl, A. Median-joining networks for inferring intraspecific phylogenies. Mol. Biol. Evol. 1999, 16, 37–48. [Google Scholar] [CrossRef]
- Raymond, M.; Rousset, F. Genepop (Version 1.2): Population-Genetics Software for Exact Tests and Ecumenicism. J. Hered. 1995, 86, 248–249. [Google Scholar] [CrossRef]
- Weir, B.S.; Cockerham, C.C. Estimating F-Statistics for the Analysis of Population-Structure. Evolution 1984, 38, 1358–1370. [Google Scholar]
- Pritchard, J.K.; Stephens, M.; Donnelly, P. Inference of population structure using multilocus genotype data. Genetics 2000, 155, 945–959. [Google Scholar] [CrossRef]
- Earl, D.A.; von Holdt, B.M. STRUCTURE HARVESTER: A website and program for visualizing STRUCTURE output and implementing the Evanno method. Conserv. Genet. Resour. 2012, 4, 359–361. [Google Scholar] [CrossRef]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef]
- Rogers, A.R.; Harpending, H. Population growth makes waves in the distribution of pairwise genetic differences. Mol. Biol. Evol. 1992, 9, 552–569. [Google Scholar] [PubMed]
- Guerrero, P.C.; Rosas, M.; Arroyo, M.T.K.; Wiens, J.J. Evolutionary lag times and recent origin of the biota of an ancient desert (Atacama-Sechura). Proc. Natl. Acad. Sci. USA 2013, 110, 11469–11474. [Google Scholar] [CrossRef] [PubMed]
- Stebbins, G.L. Aridity as a stimulus to plant evolution. Am. Nat. 1952, 86, 33–44. [Google Scholar] [CrossRef]
- Solbrig, O. The origin and floristic affinities of the South American temperate desert and semiarid regions. In Evolution of Desert Biota, 1st ed.; Goodall, D., Ed.; University of Texas Press: Austin, TX, USA, 1976; pp. 7–50. [Google Scholar]
- Moreira-Muñoz, A. Plant Geography of Chile, 1st ed.; Springer: London, UK, 2011; p. 320. [Google Scholar]
- Rosser, N.; Phillimore, A.B.; Huertas, B.; Willmott, K.R.; Mallet, J. Testing historical explanations for gradients in species richness in heliconiine butterflies of tropical America. Biol. J. Linn. Soc. 2012, 105, 479–497. [Google Scholar] [CrossRef]
- Benson, W.W.; Brown, K.S.J.; Gilbert, L.E. Coevolution of plants and herbivores: Passion flower butterflies. Evolution 1975, 29, 659–680. [Google Scholar] [CrossRef]
- Brower, A.V.Z. The evolution of ecologically important characters in Heliconius butterflies (Lepidoptera: Nymphalidae): A cladistic review. Zool. J. Linn. Soc. 1997, 119, 457–472. [Google Scholar] [CrossRef]
- Merrill, R.M.; Naisbit, R.E.; Mallet, J.; Jiggins, C.D. Ecological and genetic factors influencing the transition between host-use strategies in sympatric Heliconius butterflies. J. Evol. Biol. 2013, 26, 1959–1967. [Google Scholar] [CrossRef]
- Brown, K.S., Jr. Ecologia Geográfica e Evolução Nas Florestas Neotropicais; Universidade Estadual de Campinas: Campinas, Brazil, 1979; p. 256. [Google Scholar]
- Rodrigues, D.; Moreira, G.R.P. Geographical variation in larval host-plant use by Heliconius erato (Lepidoptera: Nymphalidae) and consequences for adult life history. Braz. J. Biol. 2002, 62, 321–332. [Google Scholar] [CrossRef]
- Jorge, L.R.; Cordeiro-Estrela, P.; Klaczko, L.B.; Moreira, G.R.P.; Freitas, A.V.L. Host-plant dependent wing phenotypic variation in the neotropical butterfly Heliconius erato. Biol. J. Linn. Soc. 2011, 102, 765–774. [Google Scholar] [CrossRef]
- Silva, A.K.; Gonçalves, G.L.; Moreira, G.R.P. Larval feeding choices in heliconians: Induced preferences are not constrained by performance and host plant phylogeny. Anim. Behav. 2014, 89, 155–162. [Google Scholar] [CrossRef]
- Thiele, S.C. Uso de Passiflora Linnaeus por Dione moneta moneta Hübner (Lepidoptera: Nymphalidae: Heliconiinae) e Consequências Para a Distribuição Geográfica. Master’s Thesis, Programa de Pós Graduação em Biologia Animal. Universidade Federal do Rio Grande do Sul (UFRGS), Porto Alegre, Brazil, 2012. [Google Scholar]
- Runquist, E.B.; Forister, M.L.; Shapiro, A.M. Phylogeography at large spatial scales: Incongruent patterns of population structure and demography of Pan-American butterflies associated with weedy habitats. J. Biogeogr. 2012, 39, 382–396. [Google Scholar] [CrossRef]
- Gilbert, L. On the ecology of natural dispersal: Dione moneta poeyii in Texas (Nymphalidae). J. Lepid. Soc. 1969, 23, 177–185. [Google Scholar]
- Hanski, I. Metapopulation dynamics. Nature 1998, 396, 41–49. [Google Scholar] [CrossRef]
- Vargas-Ortiz, M.; Aliaga-Pichihua, G.; Lazo-Rivera, A.; Cerdeña, J.; Farfán, J.; Huanca-Mamani, W.; Vargas, H.A. Cryptic Diversity in the Monotypic Neotropical Micromoth Genus Angelabella (Lepidoptera: Gracillariidae) in the Peru-Chile Desert. Insects 2020, 11, 677. [Google Scholar] [CrossRef] [PubMed]
- Jaun, A.; Wymann, H.-P.; Lucek, K. Lack of genetic structure suggests high connectivity of Parnassius phoebus between nearby valleys in the Alps. Alp. Entomol. 2022, 6, 1–6. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Source of Variation | Variance | Variation (%) | F Statistic | p |
|---|---|---|---|---|
| Among groups | 0.18 | 13.5 | 0.13 | 0.1241 |
| Among populations within groups | 0.15 | 11.6 | 0.13 | 0.0000 |
| Within populations | 0.99 | 74.7 | 0.25 | 0.0000 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Farfán, J.; Cerdeña, J.; Huanca-Mamani, W.; Vargas, H.A.; Gonçalves, G.L.; Moreira, G.R.P. Host Plant Variation and Lack of Genetic Differentiation in Populations of Dione (Agraulis) dodona Lamas & Farfán (Lepidoptera: Nymphalidae). Insects 2022, 13, 819. https://doi.org/10.3390/insects13090819
Farfán J, Cerdeña J, Huanca-Mamani W, Vargas HA, Gonçalves GL, Moreira GRP. Host Plant Variation and Lack of Genetic Differentiation in Populations of Dione (Agraulis) dodona Lamas & Farfán (Lepidoptera: Nymphalidae). Insects. 2022; 13(9):819. https://doi.org/10.3390/insects13090819
Chicago/Turabian StyleFarfán, Jackie, José Cerdeña, Wilson Huanca-Mamani, Héctor A. Vargas, Gislene L. Gonçalves, and Gilson R. P. Moreira. 2022. "Host Plant Variation and Lack of Genetic Differentiation in Populations of Dione (Agraulis) dodona Lamas & Farfán (Lepidoptera: Nymphalidae)" Insects 13, no. 9: 819. https://doi.org/10.3390/insects13090819