

Baseline Susceptibility and Cross-Resistance of HearNPV in Helicoverpa armigera (Lepidoptera: Noctuidae) in Brazil

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. Baseline Susceptibility
2.3. Validation of Diagnostic Concentration
2.4. Cross-Resistance between HearNPV and Insecticides
2.5. Statistical Analysis
3. Results
3.1. Baseline Susceptibility of H. armigera to HearNPV in Droplet Feeding Bioassays
3.2. Validation of the Candidate Diagnostic Concentration for Resistance Monitoring
3.3. Cross-Resistance between HearNPV and Insecticides
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hawkins, N.J.; Bass, C.; Dixon, A.; Neve, P. The evolutionary origins of pesticide resistance. Biol. Rev. 2019, 94, 135–155. [Google Scholar] [CrossRef] [PubMed]
- Tay, W.T.; Walsh, T.K.; Downes, S.; Anderson, C.; Jermiin, L.S.; Wong, T.K.F.; Piper, M.C.; Chang, E.S.; Macedo, I.B.; Czepak, C.; et al. Mitochondrial DNA and trade data support multiple origins of Helicoverpa armigera (Lepidoptera: Noctuidae) in Brazil. Sci. Rep. 2017, 7, 45302. [Google Scholar] [CrossRef] [PubMed]
- Forrester, N.W.; Cahill, M.; Bird, L.J.; Layland, J.K. Management of pyrethroid and endosulfan resistance in Helicoverpa armigera (Lepidoptera: Noctuidae) in Australia. Bull. Entomol. Res. 1993, Supplement No. 1, 1–132. [Google Scholar]
- Durigan, M.R.; Corrêa, A.S.; Pereira, R.M.; Leite, N.A.; Amado, D.; Sousa, D.R.; Omoto, C. High frequency of CYP337B3 gene associated with control failures of Helicoverpa armigera with pyrethroid insecticides in Brazil. Pesticide Biochem. Physiol. 2017, 143, 73–80. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Qiu, X.; Ren, X.; Zhang, W.; Wang, K. Effects of spinosad on Helicoverpa armigera (Lepidoptera: Noctuidae) from China: Tolerance status, synergism and enzymatic responses. Pest Manag. Sci. 2009, 65, 1040–1046. [Google Scholar] [CrossRef]
- Ahmad, M.; Arif, M.I.; Ahmad, Z. Resistance to carbamate insecticides in Helicoverpa armigera (Lepidoptera: Noctuidae) in Pakistan. Crop Prot. 2001, 20, 427–432. [Google Scholar] [CrossRef]
- Pereira, R.M. Caracterização da Suscetibilidade a Inseticidas Diamidas e Espinosinas em Populações de Helicoverpa armigera (Lepidoptera: Noctuidae) do Brasil; Universidade de São Paulo: Piracicaba, Brazil, 2017. [Google Scholar] [CrossRef]
- Bird, L.J. Genetics, cross-resistance and synergism of indoxacarb resistance in Helicoverpa armigera (Lepidoptera: Noctuidae). Pest Manag. Sci. 2017, 73, 575–581. [Google Scholar] [CrossRef]
- Zhang, S.P.; Cheng, H.; Gao, Y.; Wang, G.; Liang, G.; Wu, K. Mutation of an aminopeptidase N gene is associated with Helicoverpa armigera resistance to Bacillus thuringiensis Cry1Ac toxin. Insect Biochem. Mol. Biol. 2009, 39, 421–429. [Google Scholar] [CrossRef]
- Czepack, C.; Albenaz, K.C.; Vivan, L.M.; Guimarães, H.O.; Carvalhais, T. First reported occurrence of Helicoverpa armigera (Hubner) (Lepidoptera: Noctuidae) in Brazil. Pesqui. Agropecuária Trop. 2013, 43, 110–113. [Google Scholar] [CrossRef]
- Tay, W.T.; Soria, M.F.; Walsh, T.; Thomazoni, D.; Silvie, P.; Behere, G.T.; Anderson, C.; Downes, S. New World for an Old World Pest: Helicoverpa armigera (Lepidoptera: Noctuidae) in Brazil. PLoS ONE 2013, 8, 0080134. [Google Scholar] [CrossRef] [Green Version]
- Kuss, C.C.; Roggia, R.C.R.K.; Basso, C.J.; de Oliveira, M.C.N.; de Pias, O.H.C.; Roggia, S. Controle de Helicoverpa armigera (Lepidoptera: Noctuidae) em soja com inseticidas químicos e biológicos. Pesqui. Agropecuária Bras. 2016, 51, 527–536. [Google Scholar] [CrossRef]
- Leite, N.A.; Alves-Pereira, A.; Corrêa, A.S.; Zucchi, M.I.; Omoto, C. Demographics and Genetic Variability of the New World Bollworm (Helicoverpa zea) and the Old World Bollworm (Helicoverpa armigera) in Brazil. PLoS ONE 2014, 9, e113286. [Google Scholar] [CrossRef]
- Moscardi, F. Assessment of the application of Baculoviruses for control of Lepidoptera. Annu. Rev. Entomol. 1999, 44, 257–289. [Google Scholar] [CrossRef] [PubMed]
- Barreto, M.R.; Guimaraes, C.T.; Teixeira, F.F.; Paiva, E.; Valicente, F.H. Effect of Baculovirus spodoptera isolates in Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) larvae and their characterization by RAPD. Neotrop. Entomol. 2005, 34, 67–75. [Google Scholar] [CrossRef]
- Bentivenha, J.P.F.; Rodrigues, J.G.; Lima, M.F.; Marçon, P.; Popham, H.J.R.; Omoto, C. Baseline Susceptibility of Spodoptera frugiperda (Lepidoptera: Noctuidae) to SfMNPV and Evaluation of Cross-Resistance to Major Insecticides and Bt Proteins. J. Econ. Entomol. 2018, 112, 91–98. [Google Scholar] [CrossRef]
- Muraro, D.S.; Giacomelli, T.; Stacke, R.F.; Godoy, D.N.; Marçon, P.; Popham, H.J.R.; Bernardi, O. Baseline susceptibility of Brazilian populations of Chrysodeixis includens (Lepidoptera: Noctuidae) to C. includens nucleopolyhedrovirus and diagnostic concentration for resistance monitoring. J. Econ. Entomol. 2019, 112, 349–354. [Google Scholar] [CrossRef]
- Roush, R.T.; Miller, G.L. Considerations for design of insecticide resistance monitoring programs. J. Econ. Entomol. 1986, 79, 293–298. [Google Scholar] [CrossRef]
- Ffrench-Constant, R.H.; Roush, R.T. Resistance detection and documentation: The relative roles of pesticidal and biochemical assays. In Pesticide Resistance in Arthropods; Roush, R.T., Tabashnik, B.E., Eds.; Springer: New York, NY, USA, 1990; pp. 4–38. [Google Scholar] [CrossRef]
- Sparks, T.C.; Crossthwaite, A.J.; Nauen, R.; Banba, S.; Cordova, D.; Earley, F.; Ebbinghaus-Kintscher, U.; Fujioka, S.; Hirao, A.; Karmon, D.; et al. Insecticides, biologics and nematicides: Updates to IRAC’s mode of action classification—A tool for resistance management. Pesticide Biochem. Physiol. 2020, 167, 104587. [Google Scholar] [CrossRef]
- Song, J.; Wang, X.; Hou, D.; Huang, H.; Liu, X.; Deng, F.; Wang, H.; Arif, B.M.; Hu, Z.; Wang, M. The host specificities of baculovirus per os infectivity factors. PLoS ONE 2016, 11, 0159862. [Google Scholar] [CrossRef] [Green Version]
- Boogaard, B.; Van Oers, M.; Van Lent, J.W.M. An advanced view on baculovirus per os infectivity factors. Insects 2018, 9, 84. [Google Scholar] [CrossRef]
- Fuxa, J.R.; Mitchell, F.L.; Richter, A.R. Resistance of Spodoptera frugiperda (Lep.: Noctuidae) to a nuclear polyhedrosis virus in the field and laboratory. Entomophaga 1988, 33, 55–63. [Google Scholar] [CrossRef]
- Abot, A.R.; Moscardi, F.; Fuxa, J.R.; Sosa-Gómez, D.R.; Richter, A.R. Development of resistance by Anticarsia gemmatalis from Brazil and the United States to a nuclear polyhedrosis virus under laboratory selection pressure. Biol. Control 1996, 7, 126–130. [Google Scholar] [CrossRef]
- Godoy, D.N.; Führ, F.M.; Stacke, R.F.; Muraro, D.S.; Marçon, P.; Popham, H.J.R.; Bernardi, O. No cross-resistance between ChinNPV and chemical insecticides in Chrysodeixis includens (Lepidoptera: Noctuidae). J. Invertebr. Pathol. 2019, 164, 66–68. [Google Scholar] [CrossRef] [PubMed]
- Windus, L.C.; Jones, A.M.; Downes, S.; Walsh, T.; Knight, K.; Kinkema, M. HearNPV susceptibility in Helicoverpa armigera and Helicoverpa punctigera strains resistant to Bt toxins Cry1Ac, Cry2Ab, and Vip3Aa. J. Invertebr. Pathol. 2021, 183, 107598. [Google Scholar] [CrossRef]
- Durigan, M.R. Resistance to Pyrethroid and Oxadiazine Insecticides in Helicoverpa armigera (Lepidoptera: Noctuidae) Populations in Brazil; Universidade de São Paulo: Piracicaba, Brazil, 2018. [Google Scholar] [CrossRef]
- Greene, G.L.; Leppla, N.; Dickerson, W.A. Velvetbean caterpillar: A rearing procedure and artificial medium. J. Econ. Entomol. 1976, 69, 487–488. [Google Scholar] [CrossRef]
- Hughes, P.R.; Van Beek, N.A.M.; Wood, H.A. A modified droplet feeding method for rapid assay of Bacillus thuringiensis and baculoviruses in noctuidae larvae. J. Invertebr. Pathol. 1986, 48, 1–9. [Google Scholar] [CrossRef]
- Harrison, R.L.; Puttler, B.; Popham, J.R. Genomic sequence analysis of a fast-killing isolate of Spodoptera frugiperda multiple nucleopolyhedrovirus. J. Gen. Virol. 2008, 89, 775–790. [Google Scholar] [CrossRef]
- SAS Institute. Statistical Analysis System: Getting Started with the SAS Learning; SAS Institute: Cary, NC, USA, 2000. [Google Scholar]
- Robertson, J.L.; Jones, M.M.; Olguin, E.; Brad Alberts, B. Bioassays with Arthropods, 3rd ed.; CRC: Boca Raton, FL, USA, 2016; p. 2012. [Google Scholar] [CrossRef]
- Sims, S.R.; John, T.; Greenplate, J.T.; Stone, T.B.; Caprio, M.A.; Gould, F.L. Monitoring strategies for early detection of Lepidoptera resistance to Bacillus thuringiensis insecticidal proteins. In Molecular Genetics and Evolution of Pesticide Resistance; Brown, T.M., Ed.; American Chemical Society: Washington, DC, USA, 1996; pp. 229–242. [Google Scholar] [CrossRef]
- Arrizubieta, M.; Trevor, W.; Primitivo, C.; Oihane, S. Selection of a nucleopolyhedrovirus isolate from Helicoverpa armigera as the basis for a biological insecticide. Pest Manag. Sci. 2014, 70, 967–976. [Google Scholar] [CrossRef]
- Grove, M.; Hoover, K. Intrastadial developmental resistance of third instar gypsy moths (Lymantria dispar L.) to L-dispar nucleopolyhedrovirus. Biol. Control 2007, 40, 355–361. [Google Scholar] [CrossRef]
- Trudeau, D.; Washburn, J.O.; Volkman, L.E. Central role of hemocytes in Autographa californica M nucleopolyhedrovirus pathogenesis in Heliothis virescens and Helicoverpa zea. J. Virol. 2001, 75, 996–1003. [Google Scholar] [CrossRef]
- Cui, L.; Wang, Q.; Qi, H.; Wang, Q.; Yuan, H.; Rui, C. Resistance selection of indoxacarb in Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae): Cross-resistance, biochemical mechanisms and associated fitness costs. Pest Manag. Sci. 2018, 74, 2636–2644. [Google Scholar] [CrossRef] [PubMed]
- Richardson, E.B.; Troczka, B.J.; Gutbrod, O.; Davies, E.G.T.; Nauen, R. Diamide resistance: 10 years of lessons from lepidopteran pests. J. Pest Sci. 2020, 93, 911–928. [Google Scholar] [CrossRef]
- Siegwart, M.; Graillot, B.; Lopez, C.B.; Besse, S.; Bardin, M.; Nicot, P.C.; Lopez-Ferber, M. Resistance to bio-insecticides or how to enhance their sustainability: A review. Front. Plant Sci. 2015, 6, 381. [Google Scholar] [CrossRef]
- Mastrangelo, T.; Paulo, D.F.; Bergamo, L.W.; Morais, E.G.F.; Silva, M.; Bezerra-Silva, G.; Azeredo-Espin, A.M.L. Detection and Genetic Diversity of a Heliothine Invader (Lepidoptera: Noctuidae) From North and Northeast of Brazil. J. Econ. Entomol. 2014, 107, 970–980. [Google Scholar] [CrossRef] [PubMed]
- Raymond, B.; Sayyed, A.H.; Wright, D.J. The compatibility of a nucleopolyhedrosis virus control with resistance management for Bacillus thuringiensis: Co-infection and cross-resistance studies with the diamondback moth Plutella xylostella. J. Invertebr. Pathol. 2006, 93, 114–120. [Google Scholar] [CrossRef]
- Tabashnik, B.E. Managing resistance with multiple pesticide tactics: Theory, evidence, and recommendations. J. Econ. Entomol. 1989, 82, 1263–1269. [Google Scholar] [CrossRef]


{kind=link}
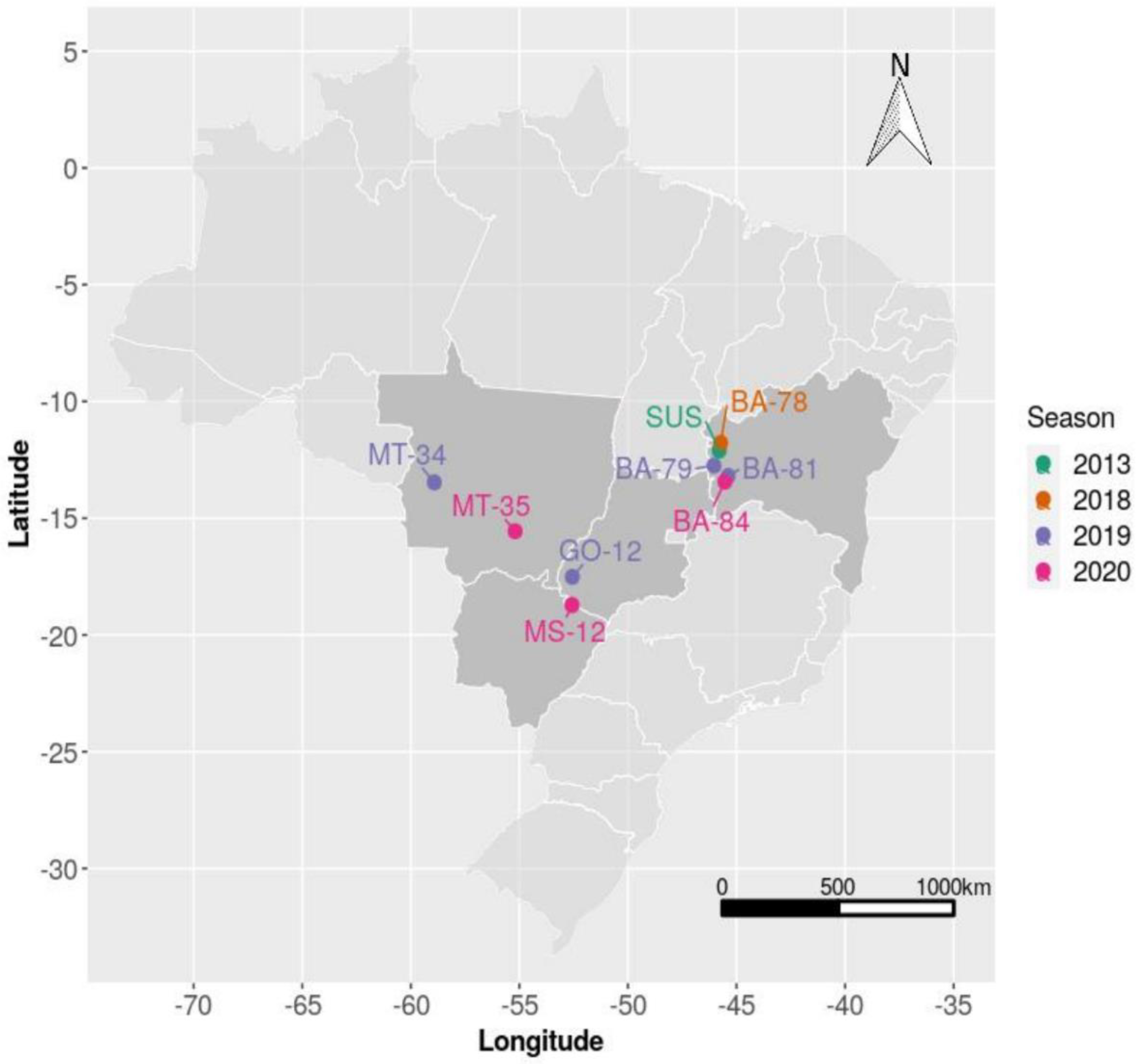
| Population Code | City, State | Host Crop | Latitude (S) | Longitude (W) | Date |
|---|---|---|---|---|---|
| SUS | Luís Eduardo Magalhães, BA | Bean | 12°05′58″ | 45°47′54″ | September 2013 |
| Season 2018 | |||||
| BA-78 | Luís Eduardo Magalhães, BA | Cotton | 11°46′33″ | 45°43′44″ | June 2018 |
| Season 2019 | |||||
| BA-79 | Roda Velha, BA | Soybean | 12°45′00″ | 46°02′25″ | December 2018 |
| BA-81 | Correntina, BA | Cotton | 13°11′34″ | 45°23′16″ | June 2019 |
| GO-12 | Mineiros, GO | Soybean | 17°30′47″ | 52°33′48″ | December 2018 |
| MT-34 | Sapezal, MT | Soybean | 13°27’55″ | 58°55’13″ | January 2019 |
| Season 2020 | |||||
| BA-84 | Correntina, BA | Soybean | 13°25′55″ | 45°32′07″ | December 2019 |
| MT-35 | Campo Verde, MT | Soybean | 15°33′29″ | 55°11′49″ | December 2019 |
| MS-12 | Chapadão do Sul-MS | Cotton | 18°43’13″ | 52°34’27″ | June 2019 |
| Population | Generation | na | Slope ± SE b | LC50 (95% CI) c | χ2 d | df e |
|---|---|---|---|---|---|---|
| SUS | F47 | 435 | 0.48 ± 0.05 | 1.1 × 106 (3.9 × 105 to 2.6 × 106) a | 4.87 | 4 |
| Season 2018 | ||||||
| BA-78 | F1 | 521 | 0.99 ± 0.16 | 7.3 × 105 (1.8 × 104 to 4.3 × 106) a | 8.55 | 4 |
| Season 2019 | ||||||
| BA-79 | F1 | 486 | 0.55 ± 0.09 | 2.5 × 105 (6.5 × 103 to 1.8 × 106) a | 9.94 | 5 |
| BA-81 | F1 | 412 | 0.49 ± 0.12 | 3.5 × 105 (5.4 × 103 to 1.7 × 106) a | 7.68 | 4 |
| GO-12 | F1 | 544 | 0.51 ± 0.09 | 4.4 × 105 (1.4 × 104 to 2.9 × 106) a | 8.91 | 4 |
| MT-34 | F1 | 543 | 0.69 ± 0.11 | 1.9 × 105 (8.6 × 104 to 4.0 × 105) a | 7.84 | 4 |
| Season 2020 | ||||||
| MT-35 | F1 | 642 | 0.53 ± 0.12 | 1.5 × 105 (6.4 × 104 to 5.1 × 105) a | 6.21 | 4 |
| Population Code | Generation | Tested | Died | % Mortality (95% CI) a |
|---|---|---|---|---|
| SUS | F47 | 450 | 445 | 98.9 (97.8–99.5) |
| BA-84 | F1 | 420 | 415 | 98.8 (97.8–99.6) |
| MT-34 | F2 | 550 | 547 | 99.5 (98.1–99.8) |
| MT-35 | F2 | 380 | 378 | 99.6 (98.5–99.8) |
| MS-12 | F1 | 450 | 450 | 100.0 (98.7–99.5) |
| Strains | Generation | n a | Slope ± SE b | LC50 (95% CI) c | χ2 d | df e | RR f |
|---|---|---|---|---|---|---|---|
| SUS | F47 | 435 | 0.48 ± 0.05 | 1.1 × 106 (3.9 × 105–2.6 × 106) a | 4.87 | 4 | - |
| FBD-R | F34 | 521 | 0.99 ± 0.16 | 7.3 × 104 (1.8 × 103–4.3 × 105) a | 8.55 | 4 | 0.06 |
| AVA-R | F18 | 458 | 0.69 ± 0.11 | 1.5 × 106 (3.5 × 103–1.4 × 106) a | 7.84 | 4 | 1.36 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Muraro, D.S.; Gonçalves, T.M.; Amado, D.; Lima, M.F.; Popham, H.J.R.; Marçon, P.G.; Omoto, C. Baseline Susceptibility and Cross-Resistance of HearNPV in Helicoverpa armigera (Lepidoptera: Noctuidae) in Brazil. Insects 2022, 13, 820. https://doi.org/10.3390/insects13090820
Muraro DS, Gonçalves TM, Amado D, Lima MF, Popham HJR, Marçon PG, Omoto C. Baseline Susceptibility and Cross-Resistance of HearNPV in Helicoverpa armigera (Lepidoptera: Noctuidae) in Brazil. Insects. 2022; 13(9):820. https://doi.org/10.3390/insects13090820
Chicago/Turabian StyleMuraro, Dionei Schmidt, Thaini M. Gonçalves, Douglas Amado, Marcelo F. Lima, Holly J. R. Popham, Paula G. Marçon, and Celso Omoto. 2022. "Baseline Susceptibility and Cross-Resistance of HearNPV in Helicoverpa armigera (Lepidoptera: Noctuidae) in Brazil" Insects 13, no. 9: 820. https://doi.org/10.3390/insects13090820