
Mosquito (Diptera: Culicidae) Diversity and Community Structure in Doi Inthanon National Park, Northern Thailand

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
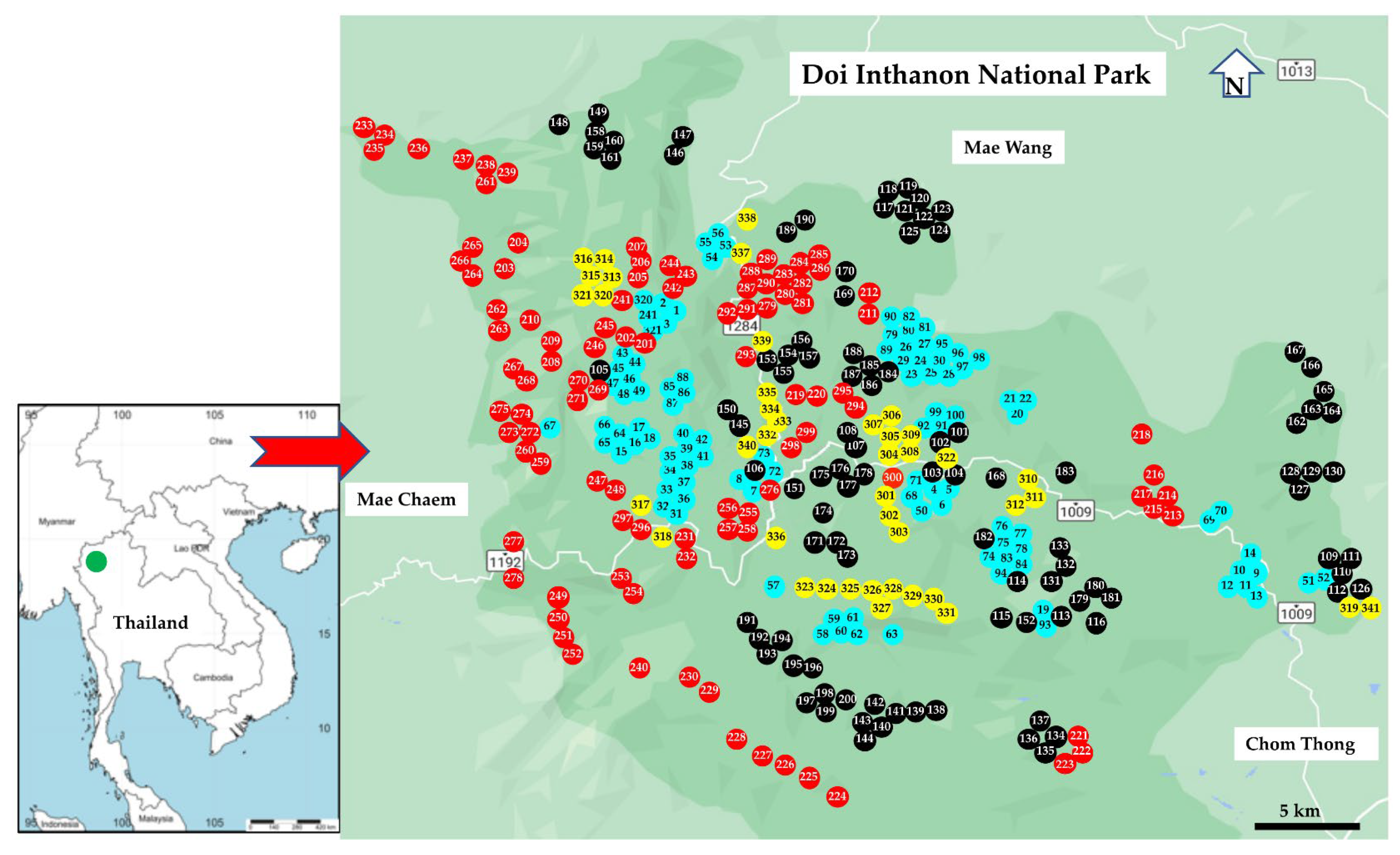
2.1. Study Area
2.2. Mosquito Collections
2.3. Larval Mosquito Habitats
2.4. Mosquito Identification
2.5. Statistical Analyses
3. Results
3.1. Species Diversity and Community Structure
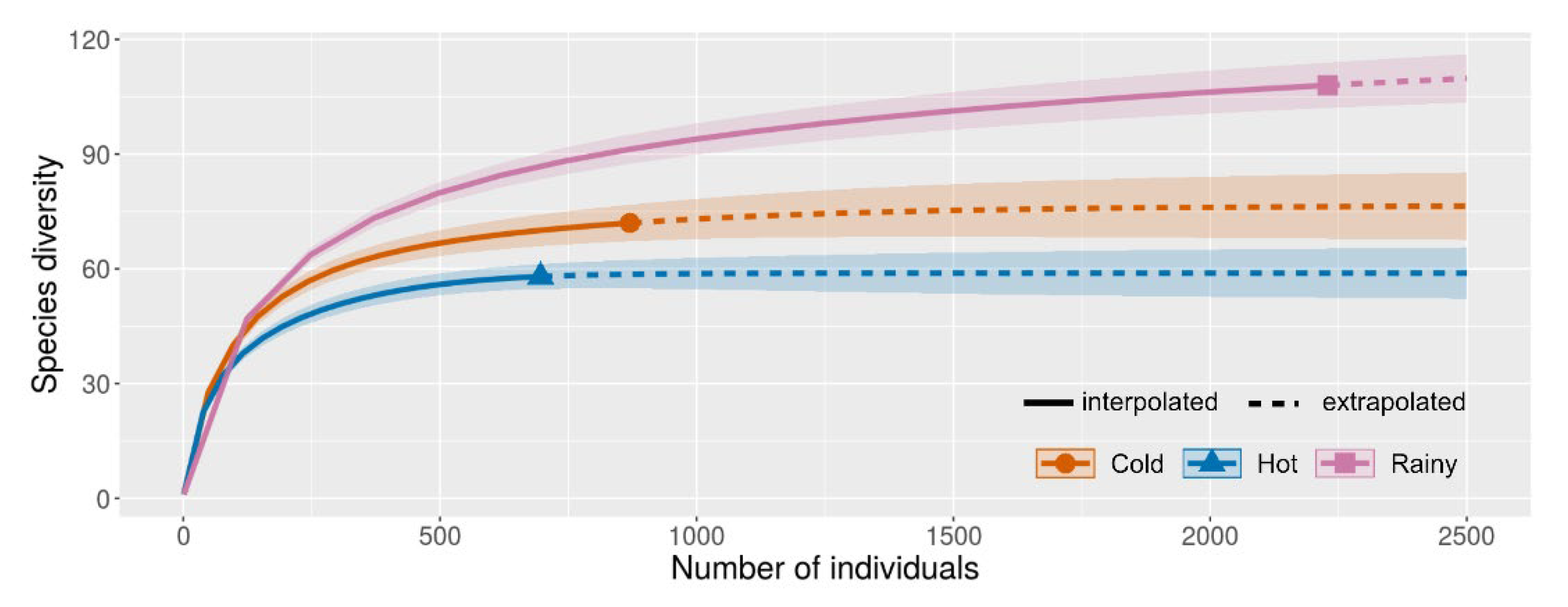
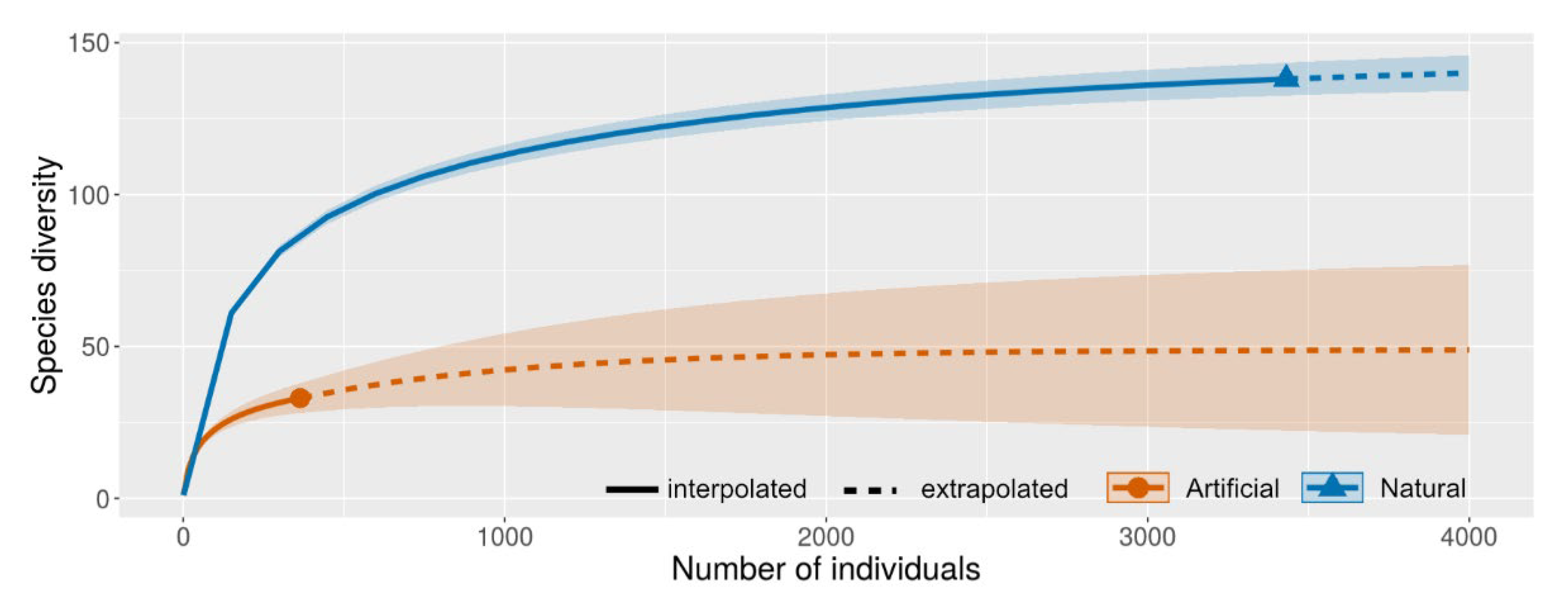
3.2. Diversity Comparisons
3.3. Aquatic Habitat Preferences
3.4. Correlation of Mosquito Species with Habitat Types
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harbach, R.E. The Culicidae (Diptera): A review of taxonomy, classification and phylogeny. Zootaxa 2007, 1668, 591–688. [Google Scholar] [CrossRef]
- World Health Organization. Dengue Bulletin, Volume 41; World Health Organization, Regional Office for South-East Asia: New Delhi, India, 2021; 202p.
- Thurman, E.B. A Contribution to a Revision of the Culicidae of Northern Thailand; University of Maryland, Agricultural Experiment Station: College Park, MD, USA, 1959; Volume A-100, pp. 1–182. [Google Scholar]
- Rattanarithikul, R.; Somboon, P.; Harbach, R.E.; Harrison, B.A. Understanding mosquitoes [in English and Thai]; Pattrara Pre-press Part., Ltd.: Chiang Mai, Thailand, 2021; Volume v18p, (In English). Volume ii19p. (In Thai) [Google Scholar]
- Bureau of Vector Borne Diseases. Annual Report 2020; Bureau of Vector Borne Diseases, Ministry of Public Health: Nonthaburi, Thailand, 2020; 146p.
- Bond, J.G.; Casas-Martínez, M.; Quiroz-Martínez, H.; Novelo-Gutiérrez, R.; Marina, C.F.; Ulloa, A.; Orozco-Bonilla, A.; Muñoz, M.; Williams, T. Diversity of mosquitoes and the aquatic insects associated with their oviposition sites along the Pacific coast of Mexico. Parasit. Vectors 2014, 7, 41. [Google Scholar] [CrossRef] [PubMed]
- Vanlalruia, K.; Senthilkumar, N.; Gurusubramanian, G. Diversity and abundance of mosquito species in relation to their larval habitats in Mizoram, North Eastern Himalayan region. Acta Trop. 2014, 137, 1–18. [Google Scholar] [CrossRef] [PubMed]
- Donald, C.L.; Siriyasatien, P.; Kohl, A. Toxorhynchites species: A review of current knowledge. Insects 2020, 11, 747. [Google Scholar] [CrossRef]
- Dexter, J.S. Mosquitoes pollinating orchids. Science 1913, 37, 867. [Google Scholar] [CrossRef]
- Thien, L.B. Mosquito pollination of Habenaria obtusata (Orchidaceae). Am. J. Bot. 1969, 56, 232–237. [Google Scholar] [CrossRef]
- Gorham, J.R. Orchid pollination by Aedes mosquitoes in Alaska. Am. Midl. Nat. 1976, 95, 208–210. [Google Scholar] [CrossRef]
- Lahondère, C.; Vinauger, C.; Okubo, R.P.; Wolff, G.H.; Chan, J.K.; Akbari, O.S.; Riffell, J.A. The olfactory basis of orchid pollination by mosquitoes. Proc. Natl. Acad. Sci. USA 2020, 117, 708–716. [Google Scholar] [CrossRef]
- Harbach, R.E. Culicidae. Mosquito Taxonomic Inventory. 2022. Available online: https://mosquito-taxonomic-inventory.myspecies.info/simpletaxonomy/term/6065 (accessed on 13 July 2022).
- Rattanarithikul, R.; Harbach, R.E.; Harrison, B.A.; Panthusiri, P.; Coleman, R.E.; Richardson, J.H. Illustrated keys to the mosquitoes of Thailand. VI. Tribe Aedini. Southeast Asian J. Trop. Med. Public Health 2010, 41 (Suppl. S1), 1–225. [Google Scholar]
- Harbach, R.E.; Kitching, I.J.; Culverwell, C.L.; Howard, T.M.; Linton, Y.-M. Nyx pholeocola, a new genus and cavernicolous species of tribe Aedini (Diptera: Culicidae) from southern Thailand based on morphological and molecular data. Zootaxa 2013, 3683, 159–177. [Google Scholar] [CrossRef]
- Harbach, R.E.; Taai, K. Nyctomyia biunguiculata, a new cavernicolous species of tribe Aedini (Diptera: Culicidae) from southern Thailand. Zootaxa 2014, 3895, 427–432. [Google Scholar] [CrossRef] [Green Version]
- Airi, M.; Kaur, S. Confirmation of Culex (Culex) tritaeniorhynchus summorosus (Diptera; Culicidae) as a separate species. J. Vector Borne Dis. 2015, 52, 219–223. [Google Scholar]
- Taai, K.; Harbach, R.E. Systematics of the Anopheles barbirostris species complex (Diptera: Culicidae: Anophelinae) in Thailand. Zool. J. Linn. Soc. 2015, 174, 244–264, 15 online tables. [Google Scholar] [CrossRef]
- Harbach, R.E.; Rattanarithikul, R.; Harrison, B.A. Anopheles prachongae, a new species of the Gigas Complex of subgenus Anopheles (Diptera: Culicidae) in Thailand, contrasted with known forms of the complex. Zootaxa 2017, 4236, 65–81. [Google Scholar] [CrossRef]
- Phanitchakun, T.; Wilai, P.; Saingamsook, J.; Namgay, R.; Drukpa, T.; Tsuda, Y.; Walton, C.; Harbach, R.E.; Somboon, P. Culex (Culiciomyia) sasai (Diptera: Culicidae), senior synonym of Cx. Spiculothorax and a new country record for Bhutan. Acta Trop. 2017, 171, 194–198. [Google Scholar] [CrossRef]
- Somboon, P.; Harbach, R.E. Lutzia (Metalutzia) chiangmaiensis n. sp. (Diptera: Culicidae), formal name for the Chiang Mai (CM) form of the genus Lutzia in Thailand. J. Med. Entomol. 2019, 56, 1270–1274. [Google Scholar] [CrossRef]
- Rattanarithikul, R.; Harrison, B.A.; Harbach, R.E.; Panthusiri, P.; Coleman, R.E. Illustrated keys to the mosquitoes of Thailand. IV. Anopheles. Southeast Asian J. Trop. Med. Public Health 2006, 37 (Suppl. S2), 1–128. [Google Scholar]
- Rattanarithikul, R.; Harrison, B.A.; Panthusiri, P.; Coleman, R.E. Illustrated keys to the mosquitoes of Thailand. I. background; geographic distribution; lists of genera, subgenera, and species; and a key to the genera. Southeast Asian J. Trop. Med. Public Health 2005, 36 (Suppl. S1), 1–80. [Google Scholar]
- Clements, A.N. The Biology of Mosquitoes. Volume 1. Development, Nutrition and Reproduction; Chapman & Hall: London, UK, 1992; Volume xxii, 509p. [Google Scholar]
- Roiz, D.; Ruiz, S.; Soriguer, R.; Figuerola, J. Climatic effects on mosquito abundance in Mediterranean wetlands. Parasit. Vectors 2014, 7, 333. [Google Scholar] [CrossRef]
- Bodharamik, T.; Juntarajumnong, W.; Apiwathanasorn, C.; Sungvornyothin, S.; Arunyawat, U. Diversity of mosquito species ovipositing in different zones of light intensity within limestone caves in Thailand. J. Am. Mosq. Control Assoc. 2018, 34, 182–189. [Google Scholar] [CrossRef]
- Oliveira-Christe, R.; Medeiros-Sousa, A.R.; Fernandes, A.; Ceretti-Júnior, W.; Marrelli, M.T. Distribution of Culex (Microculex) (Diptera: Culicidae) in forest cover gradients. Acta Trop. 2020, 202, 105264. [Google Scholar] [CrossRef] [PubMed]
- Caldwell, J.M.; LaBeaud, A.D.; Lambin, E.F.; Stewart-Ibarra, A.M.; Ndenga, B.A.; Mutuku, F.M.; Krystosik, A.R.; Ayala, E.B.; Anyamba, A.; Borbor-Cordova, M.J.; et al. Climate predicts geographic and temporal variation in mosquito-borne disease dynamics on two continents. Nat. Commun. 2021, 12, 1233. [Google Scholar] [CrossRef]
- Pernat, N.; Kampen, H.; Ruland, F.; Jeschke, J.M.; Werner, D. Drivers of spatio-temporal variation in mosquito submissions to the citizen science project ‘Mückenatlas’. Sci. Rep. 2021, 11, 1356. [Google Scholar] [CrossRef] [PubMed]
- Young, K.I.; Buenemann, M.; Vasilakis, N.; Perera, D.; Hanley, K.A. Shifts in mosquito diversity and abundance along a gradient from oil palm plantations to conterminous forests in Borneo. Ecosphere 2021, 12, e03463. [Google Scholar] [CrossRef]
- Ngom, E.H.M.; Virgillito, C.; Manica, M.; Rosà, R.; Pichler, V.; Sarleti, N.; Kassé, I.; Diallo, M.; della Torre, A.; Dia, I.; et al. Entomological survey confirms changes in mosquito composition and abundance in Senegal and reveals discrepancies among results by different host-seeking female traps. Insects 2021, 12, 692. [Google Scholar] [CrossRef] [PubMed]
- Julião, G.R.; Abad-Franch, F.; Lourenço-De-Oliveira, R.; Luz, S.L.B. Measuring mosquito diversity patterns in an Amazonian terra firme rain forest. Med. Entomol. 2010, 47, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Ngo, C.T.; Dubois, G.; Sinou, V.; Parzy, D.; Le, H.Q.; Harbach, R.E.; Manguin, S. Diversity of Anopheles mosquitoes in Binh Phuoc and Dak Nong Provinces of Vietnam and their relation to disease. Parasit. Vectors 2014, 7, 316. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Zhong, D.; Cui, L.; Lee, M.-C.; Yang, Z.; Yan, G.; Zhou, G. Population dynamics and community structure of Anopheles mosquitoes along the China-Myanmar border. Parasit. Vectors 2015, 8, 445. [Google Scholar] [CrossRef] [PubMed]
- Hutchings, R.S.G.; Hutchings, R.W.; Menezes, I.S.; Motta, M.d.A.; Sallum, M.A.M. Mosquitoes (Diptera: Culicidae) from the northwestern Brazilian Amazon: Padauari River. J. Med. Entomol. 2016, 53, 1330–1347. [Google Scholar] [CrossRef]
- Cornel, A.J.; Lee, Y.; Almeida, A.P.G.; Johnson, T.; Mouatcho, J.; Venter, M.; de Jager, C.; Braack, L. Mosquito community composition in South Africa and some neighboring countries. Parasit. Vectors 2018, 11, 331. [Google Scholar] [CrossRef]
- Almeida, J.F.; Belchior, H.C.M.; Ríos-Velásquez, C.M.; Pessoa, F.A.C. Diversity of mosquitoes (Diptera: Culicidae) collected in different types of larvitraps in an Amazon rural settlement. PloS ONE 2020, 15, e0235726. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Zhou, G.; Zhong, S.; Wang, X.; Zhong, D.; Hemming-Schroeder, E.; Yi, G.; Fu, F.; Fu, F.; Cui, L.; et al. Spatial heterogeneity and temporal dynamics of mosquito population density and community structure in Hainan Island, China. Parasit. Vectors 2020, 13, 444. [Google Scholar] [CrossRef] [PubMed]
- Junkum, A.; Pitasawat, B.; Tuetun, B.; Saeung, A.; Rattanachanpichai, E.; Jariyapan, N.; Komalamisra, N.; Mogi, M.; Chaithong, U.; Choochote, W. Seasonal abundance and biting activity of Anopheles aconitus (Diptera: Culicidae) in Chiang Mai, northern Thailand. Southeast Asian J. Trop. Med. Public Health 2007, 38 (Suppl. 1), 215–223. [Google Scholar]
- Rattanarithikul, R.; Green, C.A.; Panyim, S.; Noigamol, C.; Chanaimongkol, S.; Mahapibul, P. Laval habitats of malaria vectors and other Anopheles mosquitoes around a transmission focus in northwestern Thailand. J. Am. Mosq. Control Assoc. 1995, 11, 428–433. [Google Scholar]
- Obsomer, V.; Defourny, P.; Coosemans, M. The Anopheles dirus complex: Spatial distribution and environmental drivers. Malar. J. 2007, 6, 26. [Google Scholar] [CrossRef]
- Poolphol, P.; Harbach, R.E.; Sriwichai, P.; Srisuka, S.; Aupalee, K.; Taai, K.; Thongsahuan, S.; Uttamangkapong, S.; Phuackchantuck, R.; Morakote, N.; et al. Diversity, seasonal abundance and biting activity of Anopheles species in relation to climatic factors in northeastern Thailand. Southeast Asian J. Trop. Med. Public Health 2017, 48, 1175–1187. [Google Scholar]
- Marasri, N.; Overgaard, H.J.; Sumarnrote, A.; Thanispong, K.; Corbel, V.; Chareonviriyaphap, T. Abundance and distribution of Anopheles mosquitoes in a malaria endemic area along the Thai–Lao border. J. Vector Ecol. 2017, 42, 325–334. [Google Scholar] [CrossRef] [Green Version]
- Tainchum, K.; Ritthison, W.; Chuaycharoensuk, T.; Bangs, M.J.; Manguin, S.; Chareonviriyaphap, T. Diversity of Anopheles species and trophic behavior of putative malaria vectors in two malaria endemic areas of northwestern Thailand. J. Vector Ecol. 2014, 39, 424–436. [Google Scholar] [CrossRef]
- Tananchai, C.; Tisgratog, R.; Juntarajumnong, W.; Grieco, J.P.; Manguin, S.; Prabaripai, A.; Chareonviriyaphap, T. Species diversity and biting activity of Anopheles dirus and Anopheles baimaii (Diptera: Culicidae) in a malaria prone area of western Thailand. Parasit. Vectors 2012, 5, 211. [Google Scholar] [CrossRef]
- Tananchai, C.; Pattanakul, M.; Nararak, J.; Sinou, V.; Manguin, S.; Chareonviriyaphap, T. Diversity and biting patterns of Anopheles species in a malaria endemic area, Umphang Valley, Tak Province, western Thailand. Acta Trop. 2019, 190, 183–192. [Google Scholar] [CrossRef]
- Sumruayphol, S.; Chaiphongpachara, T.; Samung, Y.; Ruangsittichai, J.; Cui, L.; Zhong, D.; Sattabongkot, J.; Sriwichai, P. Seasonal dynamics and molecular differentiation of three natural Anopheles species (Diptera: Culicidae) of the Maculatus group (Neocellia series) in malaria hotspot villages of Thailand. Parasit. Vectors 2020, 13, 574. [Google Scholar] [CrossRef]
- Wamaket, N.; Khamprapa, O.; Chainarin, S.; Thamsawet, P.; Ninsaeng, U.; Thongsalee, S.; Suwan, V.; Sakolvaree, J.; Takhampunya, R.; Davidson, S.A.; et al. Anopheles bionomics in a malaria endemic area of southern Thailand. Parasit. Vectors 2021, 14, 378. [Google Scholar] [CrossRef]
- National Parks Wildlife and Plant Conservation Department. The Best of National Parks of Thailand; National Park: Bangkok, Thailand, 2004; 152p.
- Teejuntuk, S.; Sahunalu, P.; Sakurai, K.; Sungpalee, W. Forest structure and tree species diversity along an altitudinal gradient in Doi Inthanon National Park, northern Thailand. Tropics 2003, 12, 85–102. [Google Scholar] [CrossRef]
- Khamyong, S.; Lykke, A.M.; Seramethakun, D.; Barfod, A.S. Species composition and vegetation structure of an upper montane forest at the summit of Mt. Doi Inthanon, Thailand. Nord. J. Bot. 2004, 23, 83–97. [Google Scholar] [CrossRef]
- Sungkajanttranon, O.; Marod, D.; Thanompun, K. Diversity and distribution of family Araceae in Doi Inthanon National Park, Chiang Mai province. Agric. Nat. Resour. 2018, 52, 125–131. [Google Scholar] [CrossRef]
- Adler, P.H.; Srisuka, S.; Low, V.L.; Takaoka, H.; Saeung, A. High-elevation chromosomal diversity of black flies (Diptera: Simuliidae) in Thailand. Insect Syst. Divers. 2019, 3, 1–10. [Google Scholar] [CrossRef]
- Plant, A.R.; Bickel, D.J.; Chatelain, P.; Daugeron, C.; Srisuka, W. Composition and organization of highly speciose Empidoidea (Diptera) communities in tropical montane forests of northern Thailand. Zootaxa 2019, 4590, 1–39. [Google Scholar] [CrossRef]
- Plant, A.R.; Bickel, D.J.; Chatelain, P.; Daugeron, C.; Srisuka, W. Endemism, similarity and difference in montane evergreen forest biodiversity hotspots: Comparing communities of Empidoidea (Insecta: Diptera) in the summit zones of Doi Inthanon and Doi Phahompok, Thailand. Trop. Nat. Hist. 2020, 20, 16–27. [Google Scholar]
- Betz, O.; Srisuka, W.; Puthz, V. Elevational gradients of species richness, community structure, and niche occupation of tropical rove beetles (Coleoptera: Staphylinidae: Steninae) across mountain slopes in Northern Thailand. Evol. Ecol. 2020, 34, 193–216. [Google Scholar] [CrossRef]
- Srisuka, W.; Sulin, C.; Aupalee, K.; Phankaen, T.; Taai, K.; Thongsahuan, S.; Saeung, A.; Takaoka, H. Community structure, biodiversity and spatiotemporal distribution of the black flies (Diptera: Simuliidae) using malaise traps on the highest mountain in Thailand. Insects 2021, 12, 504. [Google Scholar] [CrossRef]
- Kuraji, K.; Punyatrong, K. Long-term observation of an altitudinal increase in rainfall over Doi Inthanon and the Mae Chaem watershed, northern Thailand. In Proceedings of the FORTROP II: Tropical Forestry Change in a Changing World, Kasetsart University, Bangkok, Thailand, 17–20 November 2008; Volume 4, pp. 67–75. [Google Scholar]
- McKinney, M.L. Urbanization, Biodiversity, and Conservation: The impacts of urbanization on native species are poorly studied, but educating a highly urbanized human population about these impacts can greatly improve species conservation in all ecosystems. BioScience 2002, 52, 883–890. [Google Scholar] [CrossRef]
- Čabanová, V.; Miterpáková, M.; Valentová, D.; Blažĕjová, H.; Rudolf, I.; Stloukal, E.; Hurníková, Z.; Dzidová, M. Urbanization impact on mosquito community and the transmission potential of filarial infection in central Europe. Parasit. Vectors 2018, 11, 261. [Google Scholar] [CrossRef] [PubMed]
- Wilke, A.B.B.; Chase, C.; Vasquez, C.; Carvajal, A.; Medina, J.; Petrie, W.D.; Beier, J.C. Urbanization creates diverse aquatic habitats for immature mosquitoes in urban areas. Sci. Rep. 2019, 9, 15335. [Google Scholar] [CrossRef] [PubMed]
- Wilke, A.B.B.; Vasquez, C.; Medina, J.; Carvajal, A.; Petrie, W.; Beier, J.C. Community composition and year-round abundance of vector species of mosquitoes make Miami Dade County, Florida a receptive gateway for arbovirus entry to the United States. Sci. Rep. 2019, 9, 8732. [Google Scholar] [CrossRef]
- Beaulieu, M.R.S.; Federico, J.L.; Reiskind, M.H. Mosquito diversity and dog heartworm prevalence in suburban areas. Parasit. Vectors 2020, 13, 12. [Google Scholar] [CrossRef]
- Kolimenakis, A.; Heinz, S.; Wilson, M.L.; Winkler, V.; Yakob, L.; Michaelakis, A.; Papachristos, D.; Richardson, C.; Horstick, O. The role of urbanisation in the spread of Aedes mosquitoes and the diseases they transmit—A systematic review. PLoS Negl. Trop. Dis. 2021, 15, e0009631. [Google Scholar] [CrossRef]
- Pernat, N.; Kampen, H.; Jeschke, J.M.; Werner, D. Buzzing homes: Using citizen science data to explore the effects of urbanization on indoor mosquito communities. Insects 2021, 12, 374. [Google Scholar] [CrossRef]
- Climatological Group, Meteorological Development Bureau, Meteorological Department. The Climate of Thailand. 2015. Available online: https://www.tmd.go.th/en/archive/thailand_climate.pdf (accessed on 9 December 2021).
- Rattanarithikul, R.; Harbach, R.E.; Harrison, B.A.; Panthusiri, P.; Jones, J.W.; Coleman, R.E. Illustrated keys to the mosquitoes of Thailand. II. Genera Culex and Lutzia. Southeast Asian J. Trop. Med. Public Health 2005, 36 (Suppl. S2), 1–97. [Google Scholar]
- Rattanarithikul, R.; Harrison, B.A.; Panthusiri, P.; Peyton, E.L.; Coleman, R.E. Illustrated keys to the mosquitoes of Thailand. III. Genera Aedeomyia, Ficalbia, Mimomyia, Hodgesia, Coquillettidia, Mansonia, and Uranotaenia. Southeast Asian J. Trop. Med. Public Health 2006, 37 (Suppl. 1), 1–85. [Google Scholar]
- Rattanarithikul, R.; Harbach, R.E.; Harrison, B.A.; Panthusiri, P.; Coleman, R.E. Illustrated keys to the mosquitoes of Thailand. V. Genera Orthopodomyia, Kimia, Malaya, Topomyia, Tripteroides, and Toxorhynchites. Southeast Asian J. Trop. Med. Public Health 2007, 37 (Suppl. S2), 1–65. [Google Scholar]
- Wilkerson, R.C.; Linton, Y.-M.; Fonseca, D.M.; Schultz, T.R.; Price, D.C.; Strickman, D.A. Making mosquito taxonomy useful: A stable classification of tribe Aedini that balances utility with current knowledge of evolutionary relationships. PLoS ONE 2015, 10, 7. [Google Scholar] [CrossRef]
- Wilkerson, R.C.; Linton, Y.-M.; Strickman, D. Mosquitoes of the World; Johns Hopkins University Press: Baltimore, MD, USA, 2021; Volumes 1 and 2, 1332p. [Google Scholar]
- Harbach, R.E. Culicipedia: Species-Group, Genus-Group and Family-Group Names in Culicidae (Diptera); CABI: Wallingford, UK, 2018; Volume xviii, 378p. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis. Palaeontol. Electron. 2001, 4, 9. [Google Scholar]
- Hsieh, T.C.; Ma, K.H.; Chao, A. iNEXT: An R package for rarefaction and extrapolation of species diversity (Hill numbers). Methods Ecol. Evol. 2016, 7, 1451–1456. [Google Scholar] [CrossRef]
- Roell, Y.E.; Phillips, J.G.; Parent, C.E. Effect of topographic complexity on species richness in the Galápagos Islands. J. Biogeogr. 2020, 48, 2645–2655. [Google Scholar] [CrossRef]
- Hutchings, R.S.G.; Sallum, M.A.M.; Hutchings, R.W. Mosquito (Diptera: Culicidae) diversity of a forest-fragment mosaic in the Amazon rain forest. J. Med. Entomol. 2011, 48, 173–187. [Google Scholar] [CrossRef]
- Ceretti-Junior, W.; Oliveira-Christe, R.; Wilk-da-Silva, R.; Mucci, L.F.; de Castro Duarte, A.M.R.; Fernandes, A.; Barrio-Nuevo, K.M.; Carvalho, M.P.; Marrelli, M.T.; Medeiros-Sousa, A.R. Diversity analysis and an updated list of mosquitoes (Diptera: Culicidae) found in Cantareira State Park, São Paulo, Brazil. Acta Trop. 2020, 212, 105669. [Google Scholar] [CrossRef]
- Vu, N.S.; Hertz, J.C.; Martin, N.J.; Tran, T.C.; Fiorenzano, J.M.; Tran, P.V.; Nguyen, H.V.; Dang, A.D.; Tran, D.N.; Motoki, M.T. Mosquitoes (Diptera: Culicidae) from villages and forest areas of rural communes in Khanh Hoa and Binh Phuoc Provinces, Vietnam. J. Med. Entomol. 2021, 58, 2264–2273. [Google Scholar] [CrossRef]
- Andrews, E.S.; Schoeler, G.B.; Gozalo, A.S.; Carbajal, F.; Lopez-Sifuentes, V.; Turell, M.J. Species diversity, seasonal, and spatial distribution of mosquitoes (Diptera: Culicidae) captured in Aotus monkey-baited traps in a forested site near Iquitos, Peru. J. Med. Entomol. 2014, 51, 1127–1135. [Google Scholar] [CrossRef]
- Andrade, D.C.; Morais, S.A.; Marteis, L.S.; Gama, R.A.; de Melo Freire, R.C.; Rekowski, B.S.; Ueno, H.M.; La Corte, R. Diversity of mosquitoes (Diptera: Culicidae) in the Caatinga biome, Brazil, from the widespread to the endemic. Insects 2020, 11, 468. [Google Scholar] [CrossRef]
- Emidi, B.; Kisinza, W.N.; Mmbando, B.P.; Malima, R.; Mosha, F.W. Effect of physicochemical parameters on Anopheles and Culex mosquito larvae abundance in different breeding sites in a rural setting of Muheza, Tanzania. Parasit. Vectors 2017, 10, 304. [Google Scholar] [CrossRef]
- Medeiros-Sousa, A.R.; de Oliveira-Christe, R.; Camargo, A.A.; Scinachi, C.A.; Milani, G.M.; Urbinatti, P.R.; Natal, D.; Ceretti-Junior, W.; Marrelli, M.T. Influence of water’s physical and chemical parameters on mosquito (Diptera: Culicidae) assemblages in larval habitats in urban parks of São Paulo, Brazil. Acta Trop. 2020, 205, 105394. [Google Scholar] [CrossRef] [PubMed]
- Multini, L.C.; Oliveira-Christe, R.; Medeiros-Sousa, A.R.; Evangelista, E.; Barrio-Nuevo, K.M.; Mucci, L.F.; Ceretti-Junior, W.; Camargo, A.A.; Wilke, A.B.B.; Marrelli, M.T. The influence of the pH and salinity of water in breeding sites on the occurrence and community composition of immature mosquitoes in the Green Belt of the city of São Paulo, Brazil. Insects 2021, 12, 797. [Google Scholar] [CrossRef]
- Mayi, M.P.A.; Foncha, D.F.; Kowo, C.; Tchuinkam, T.; Brisco, K.; Anong, D.N.; Ravinder, S.; Cornel, A.J. Impact of deforestation on the abundance, diversity, and richness of Culex mosquitoes in a southwest Cameroon tropical rainforest. J. Vector Ecol. 2019, 44, 271–281. [Google Scholar] [CrossRef]
- Bastos, A.Q.; de Mello, C.F.; Dos Santos Silva, J.; Gil-Santana, H.R.; Silva, S.O.F.; Alencar, J. Diversity of Mosquitoes (Diptera: Culicidae) in the Bom Retiro Private Natural Heritage Reserve, Rio de Janeiro State, Brazil. J. Med. Entomol. 2022, 59, 446–453. [Google Scholar] [CrossRef]
- Harbach, R.E. Culex Linnaeus, 1758. Mosquito Taxonomic Inventory. 2022. Available online: https://mosquito-taxonomic-inventory.myspecies.info/simpletaxonomy/term/6165 (accessed on 13 July 2022).
- Walter Reed Biosystematics Unit. Culex Linnaeus, 1758. 2021. Available online: http://wrbu.si.edu//vectorspecies/genera/culex (accessed on 13 July 2022).
- Almirón, W.R.; Brewer, M.E. Classification of immature stage habitats of Culicidae (Diptera) collected in Córdoba, Argentina. Mem Inst Oswaldo Cruz. 1996, 91, 1–9. [Google Scholar] [CrossRef]
- Vanwambeke, S.O.; Somboon, P.; Harbach, R.E.; Isenstadt, M.; Lambin, E.F.; Walton, C.; Butlin, R.K. Landscape and land cover factors influence the presence of Aedes and Anopheles larvae. J. Med. Entomol. 2007, 44, 133–144. [Google Scholar] [CrossRef]
- Piovezan, R.; Rosa, S.L.; Rocha, M.L.; de Azevedo, T.S.; Von Zuben, C.J. Entomological surveillance, spatial distribution, and diversity of Culicidae (Diptera) immatures in a rural area of the Atlantic Forest biome, State of São Paulo, Brazil. J. Vector Ecol. 2013, 38, 317–325. [Google Scholar] [CrossRef]
- Saleeza, S.N.R.; Norma-Rashid, Y.; Azirun, M.S. Mosquito species and outdoor breeding places in residential areas in Malaysia. Southeast Asian J. Trop. Med. Public Health 2013, 44, 963–969. [Google Scholar]
- Hendy, A.; Hernandez-Acosta, E.; Chaves, B.A.; Ferreira Fé, N.F.; Valério, D.; Mendonça, C.; de Lacerda, M.V.G.; Buenemann, M.; Vasilakis, N.; Hanley, K.A. Into the woods: Changes in mosquito community composition and presence of key vectors at increasing distances from the urban edge in urban forest parks in Manaus, Brazil. Acta Trop. 2020, 206, 105441. [Google Scholar] [CrossRef]
- Li, S.; Jiang, F.; Lu, H.; Kang, X.; Wang, Y.; Zou, Z.; Wen, D.; Zheng, A.; Liu, C.; Liu, Q.; et al. Mosquito diversity and population genetic structure of six mosquito species from Hainan Island. Front. Genet. 2020, 11, 602863. [Google Scholar] [CrossRef]
- Leisnham, P.T.; LaDeau, S.L.; Saunders, M.E.M.; Villena, O.C. Condition-specific competitive effects of the invasive mosquito Aedes albopictus on the resident Culex pipiens among different urban container habitats may explain their coexistence in the field. Insects 2021, 12, 993. [Google Scholar] [CrossRef] [PubMed]
- Somboon, P.; Thongwat, D.; Harbach, R.E. Anopheles (Cellia) rampae n. sp., alias chromosomal form K of the Oriental Maculatus Group (Diptera: Culicidae) in Southeast Asia. Zootaxa 2011, 2810, 47–55. [Google Scholar] [CrossRef]
- Hendri, J.; Astuti, E.P.; Prasetyowati, H.; Dhewantara, P.W.; Hadi, U.K. Anopheline diversity in Indonesia: An evaluation of animal-baited sampling techniques. J. Med. Entomol. 2022, 59, 710–718. [Google Scholar] [CrossRef]
- Animut, A.; Gebre-Michael, T.; Balkew, M.; Lindtjørn, B. Abundance and dynamics of anopheline larvae in a highland malarious area of south-central Ethiopia. Parasit. Vectors 2012, 5, 117. [Google Scholar] [CrossRef]
- Harbach, R.E. Anophelinae Grassi, 1900. Mosquito Taxonomic Inventory. 2022. Available online: https://mosquito-taxonomic-inventory.myspecies.info/simpletaxonomy/term/6046 (accessed on 13 July 2022).
- Walter Reed Biosystematics Unit. Anopheles Meigen, 1818. 2021. Available online: http://wrbu.si.edu//vectorspecies/genera/anopheles (accessed on 13 July 2022).
- Reid, J.A. Anopheline Mosquitoes of Malaya and Borneo; Studies from the Institute for Medical Research Malaya, No. 31; Government of Malaysia: Kuala Lumpur, Malaysia, 1968; Volume xiii, 520p.
- Hinne, I.A.; Attah, S.K.; Mensah, B.A.; Forson, A.O.; Afrane, Y.A. Larval habitat diversity and Anopheles mosquito species distribution in different ecological zones in Ghana. Parasit. Vectors 2021, 14, 193. [Google Scholar] [CrossRef]
- Abella-Medrano, C.A.; Ibáñez-Bernal, S.; MacGregor-Fors, I.; Santiago-Alarcon, D. Spatiotemporal variation of mosquito diversity (Diptera: Culicidae) at places with different land-use types within a neotropical montane cloud forest matrix. Parasit. Vectors 2015, 8, 487. [Google Scholar] [CrossRef]
- Sulesco, T.M.; Toderas, L.G.; Uspenskaia, I.G.; Toderas, I.K. Larval habitats diversity and distribution of the mosquito (Diptera: Culicidae) species in the Republic of Moldova. J. Med. Entomol. 2015, 52, 1299–1308. [Google Scholar] [CrossRef]












{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Diversity | Season | Aquatic Habitats | Community | |||||
|---|---|---|---|---|---|---|---|---|
| Hot | Rainy | Cold | Natural | Artificial | Forest | Agriculture | Village | |
| Species richness | 72 | 108 | 58 | 138 | 33 | 108 | 88 | 57 |
| Abundance | 696 | 2229 | 870 | 3431 | 364 | 2173 | 1035 | 587 |
| Shannon_H | 3.55 | 3.81 | 3.72 | 4.20 | 2.82 | 3.97 | 3.81 | 3.21 |
| Dominance_D | 0.040 | 0.039 | 0.037 | 0.023 | 0.094 | 0.028 | 0.036 | 0.072 |
| Simpson_1-D | 0.960 | 0.961 | 0.963 | 0.977 | 0.906 | 0.972 | 0.964 | 0.928 |
| Evenness_e^H/S | 0.598 | 0.419 | 0.574 | 0.483 | 0.506 | 0.488 | 0.514 | 0.436 |
| Equitability_J | 0.873 | 0.814 | 0.870 | 0.852 | 0.806 | 0.847 | 0.852 | 0.795 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Srisuka, W.; Sulin, C.; Sommitr, W.; Rattanarithikul, R.; Aupalee, K.; Saeung, A.; Harbach, R.E. Mosquito (Diptera: Culicidae) Diversity and Community Structure in Doi Inthanon National Park, Northern Thailand. Insects 2022, 13, 814. https://doi.org/10.3390/insects13090814
Srisuka W, Sulin C, Sommitr W, Rattanarithikul R, Aupalee K, Saeung A, Harbach RE. Mosquito (Diptera: Culicidae) Diversity and Community Structure in Doi Inthanon National Park, Northern Thailand. Insects. 2022; 13(9):814. https://doi.org/10.3390/insects13090814
Chicago/Turabian StyleSrisuka, Wichai, Chayanit Sulin, Wirat Sommitr, Rampa Rattanarithikul, Kittipat Aupalee, Atiporn Saeung, and Ralph E. Harbach. 2022. "Mosquito (Diptera: Culicidae) Diversity and Community Structure in Doi Inthanon National Park, Northern Thailand" Insects 13, no. 9: 814. https://doi.org/10.3390/insects13090814