
An Ecological Profile of Hydropsyche alternans (Trichoptera: Hydropsychidae) in Lake Superior, the Last Stronghold of a Once-Dominant Great Lakes Surf Zone Caddisfly

Abstract
:Simple Summary
Abstract
1. Introduction
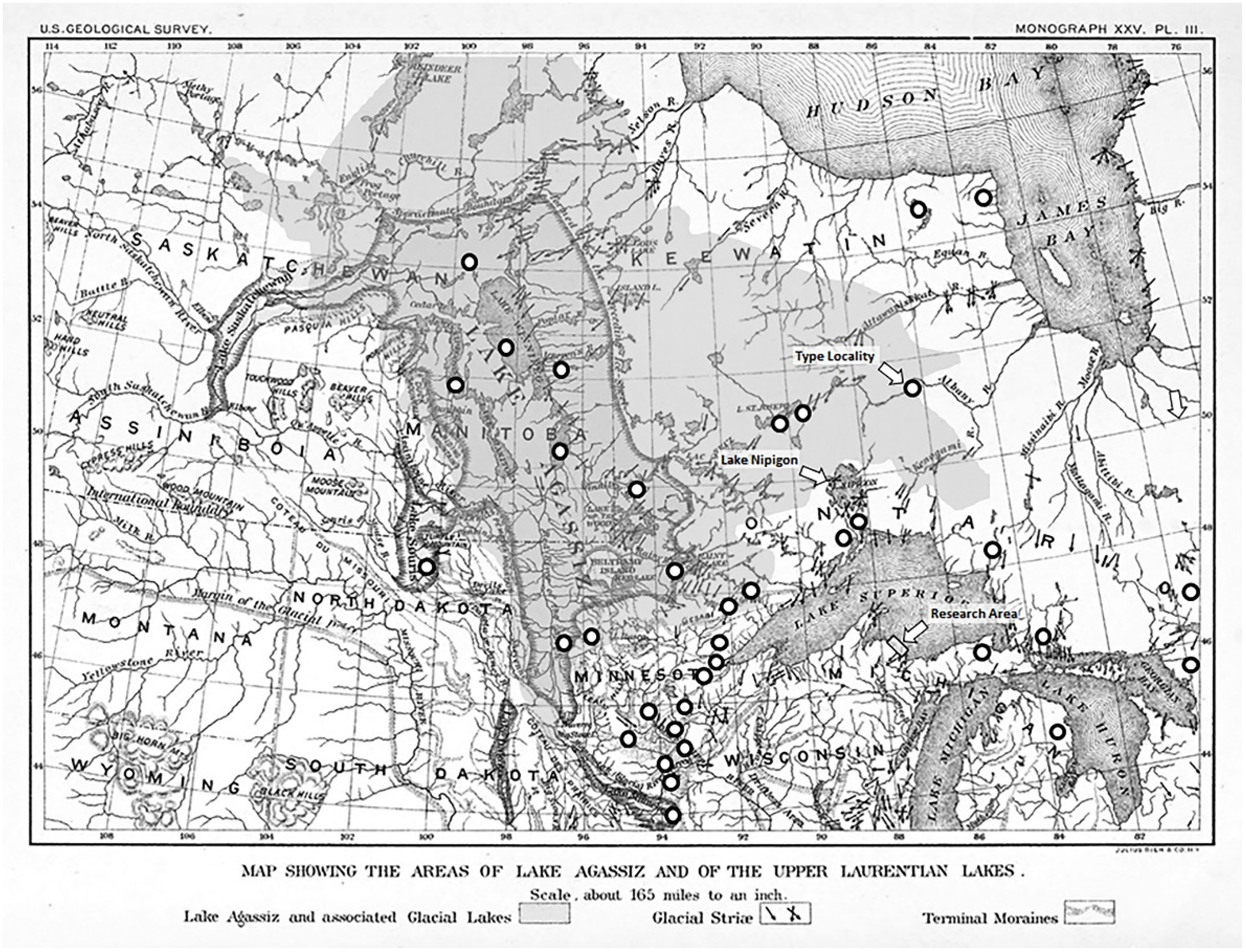
Habitat and Historical Biogeography
2. Materials and Methods
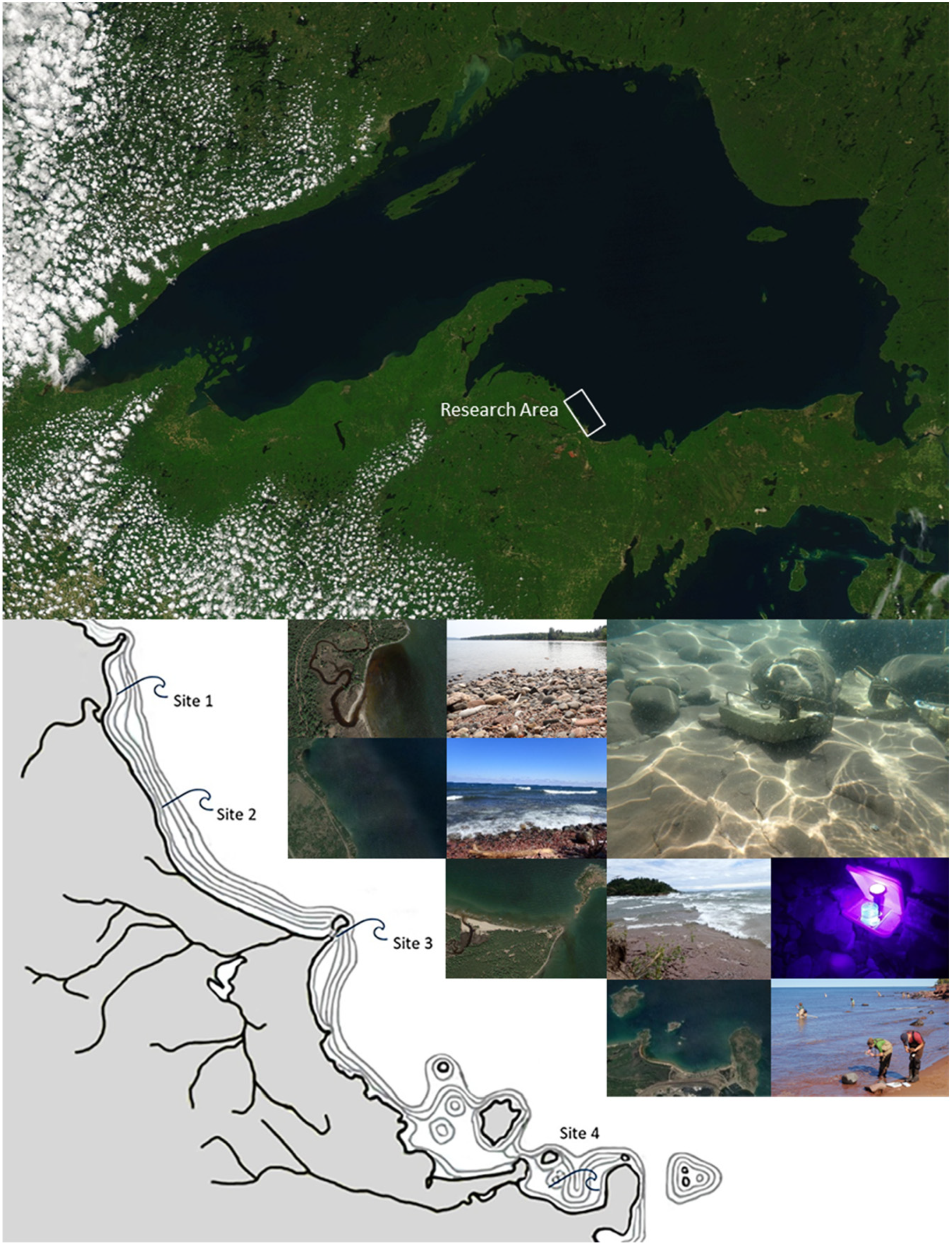
2.1. Research Area
2.2. Life History Analysis
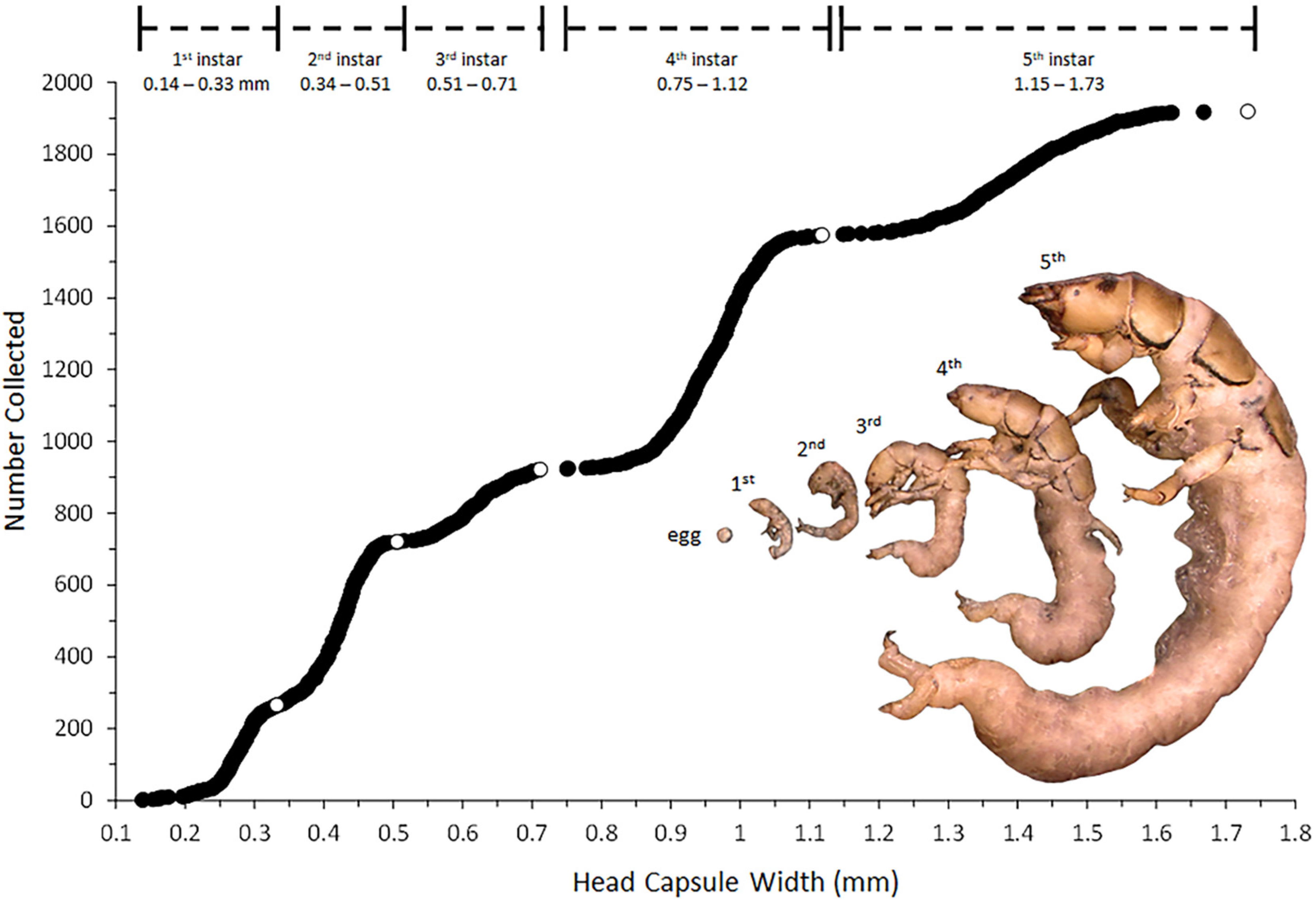
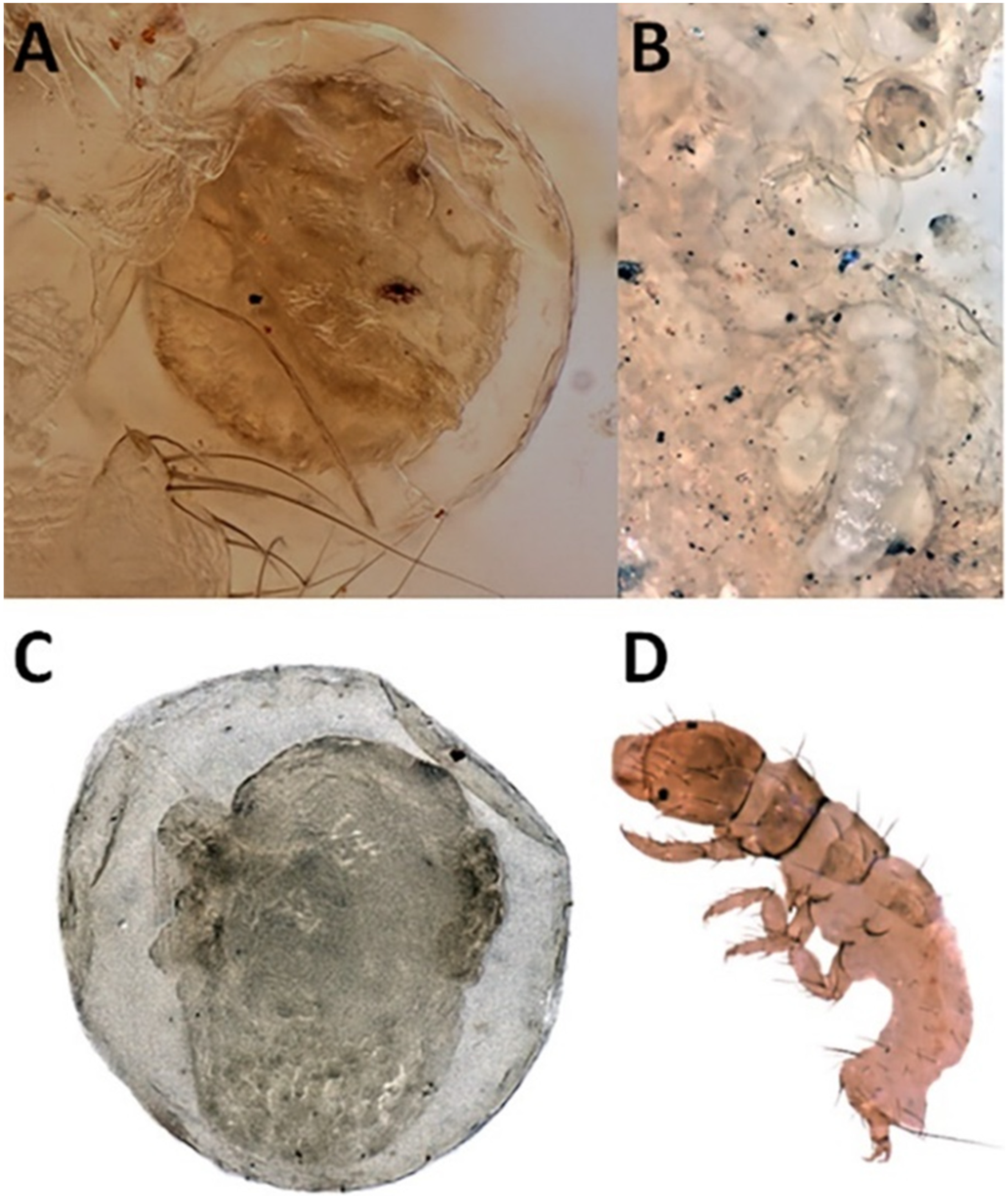
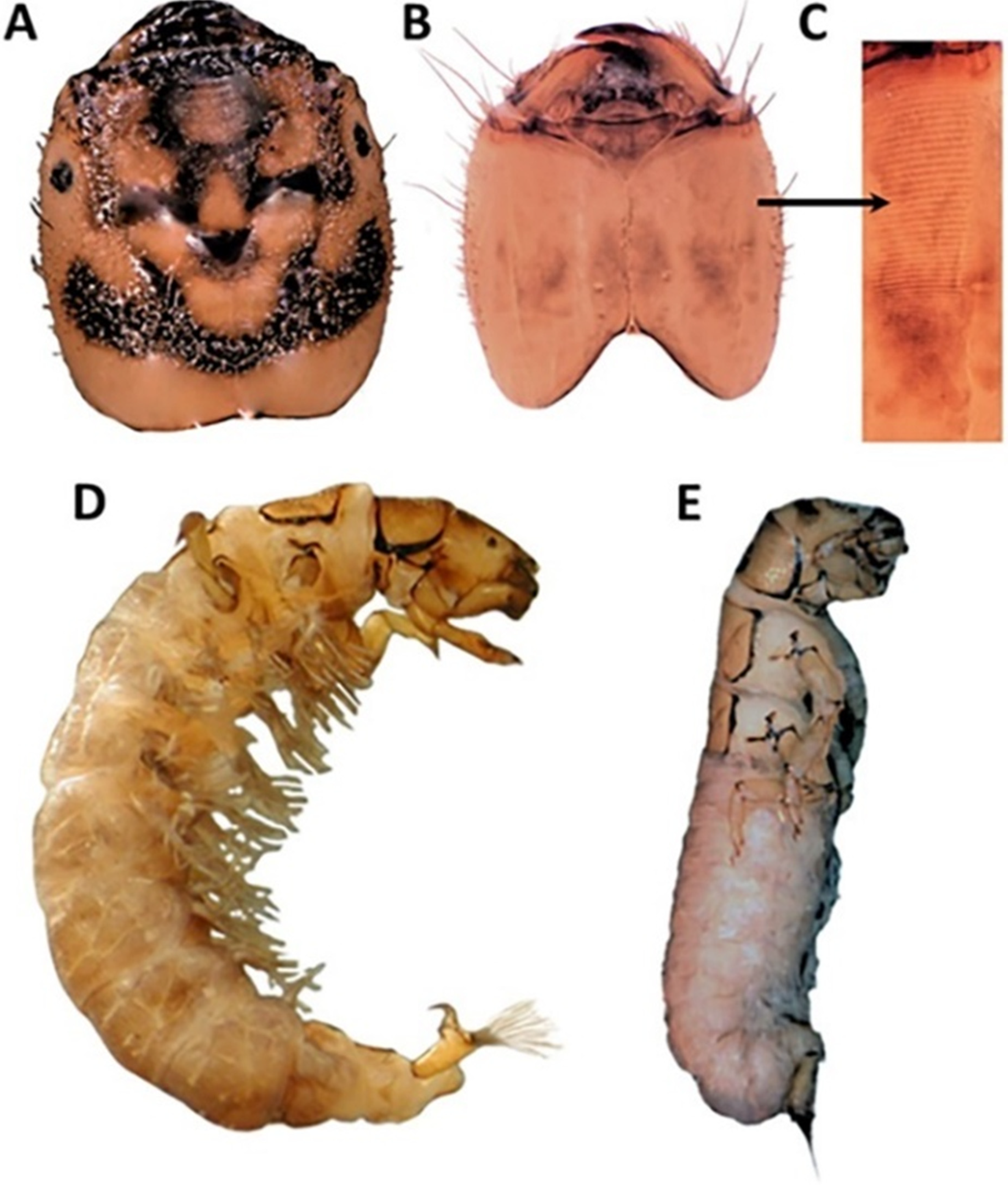
2.2.1. Larval Instar Determination
2.2.2. Adult Flight Period
2.2.3. Adult Leg Morphology
2.3. Gut Content Analyses
2.4. Stable Isotope Analyses
2.5. Data Analyses
3. Results
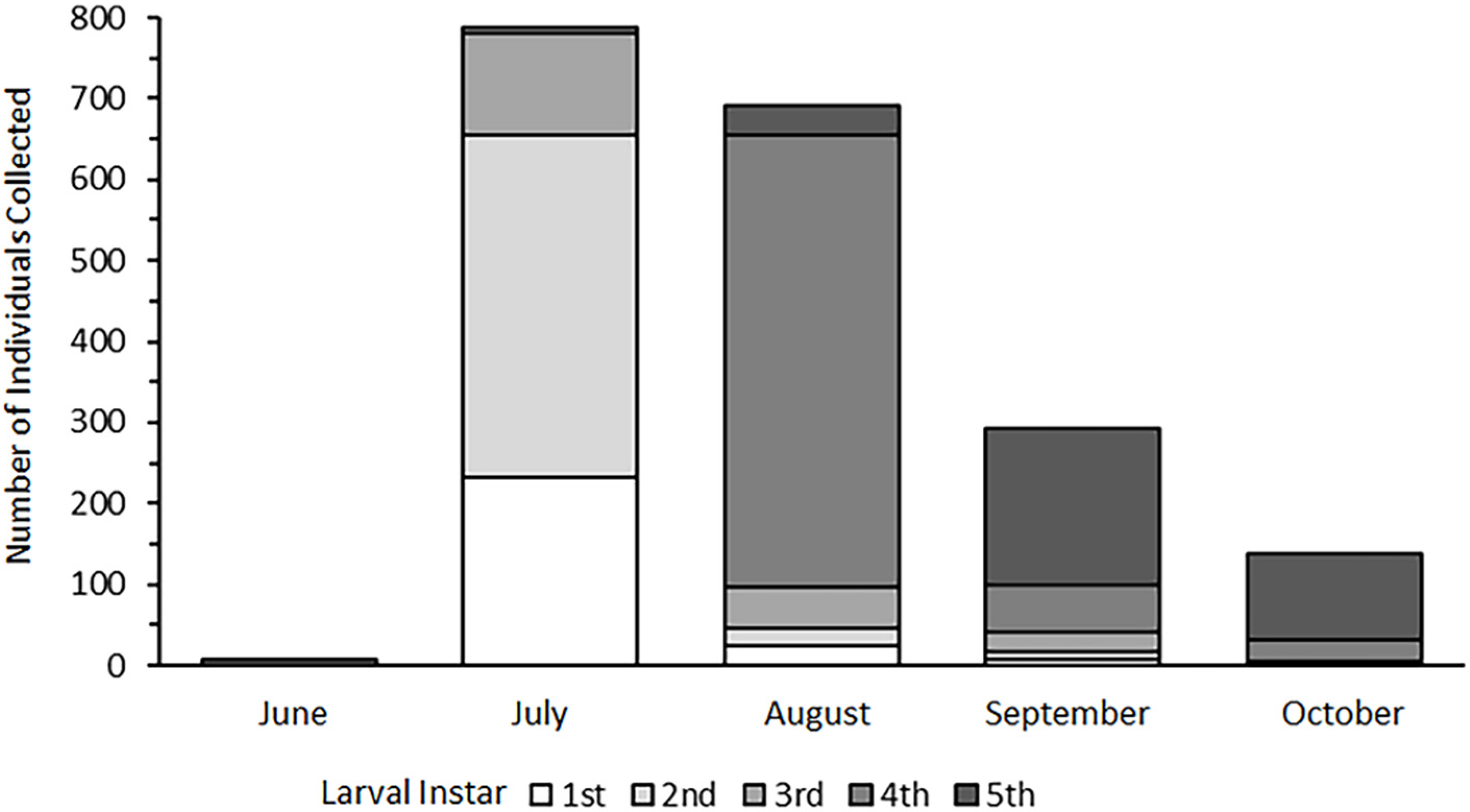
3.1. Life History Analysis
3.2. Adult Morphology
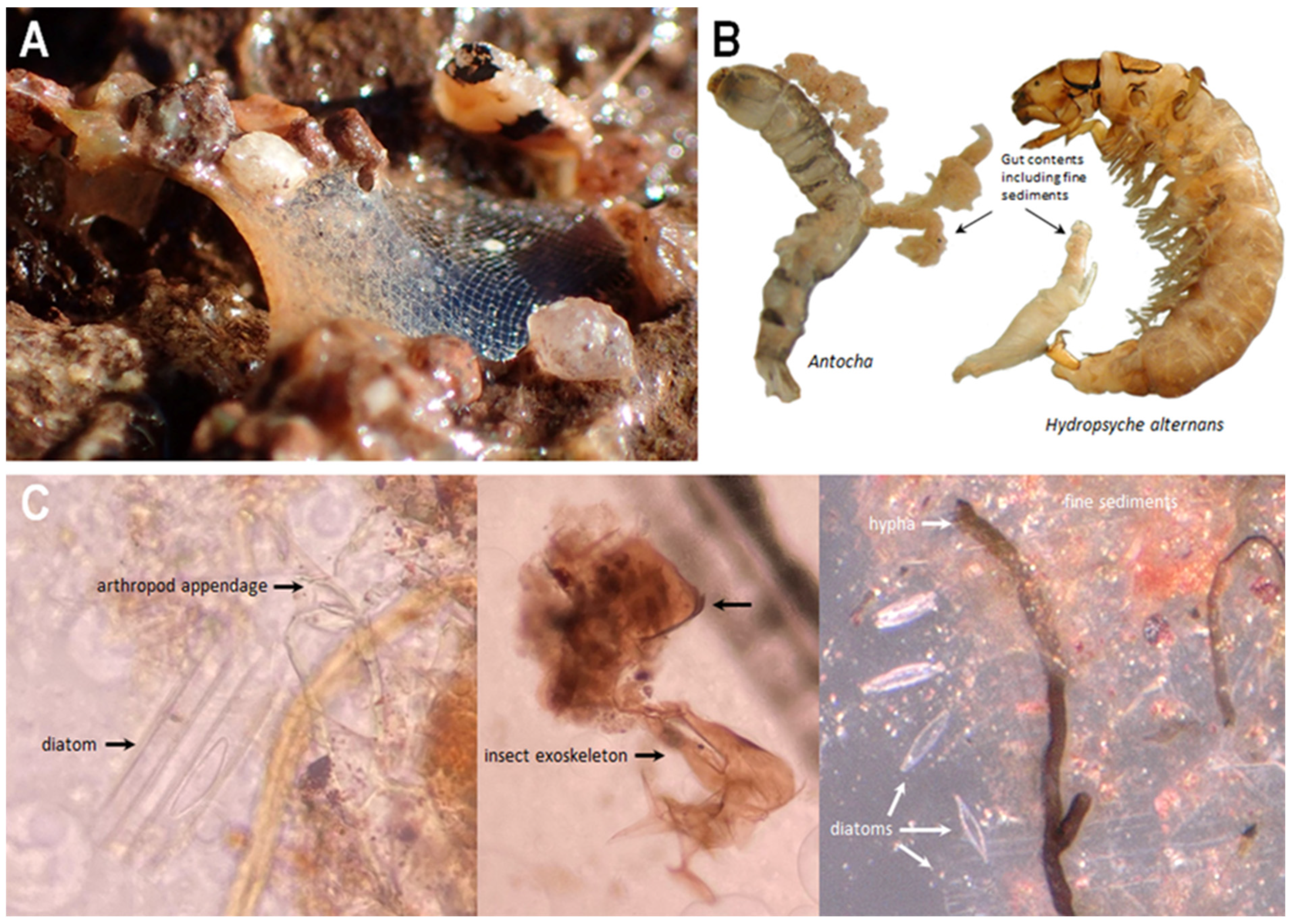
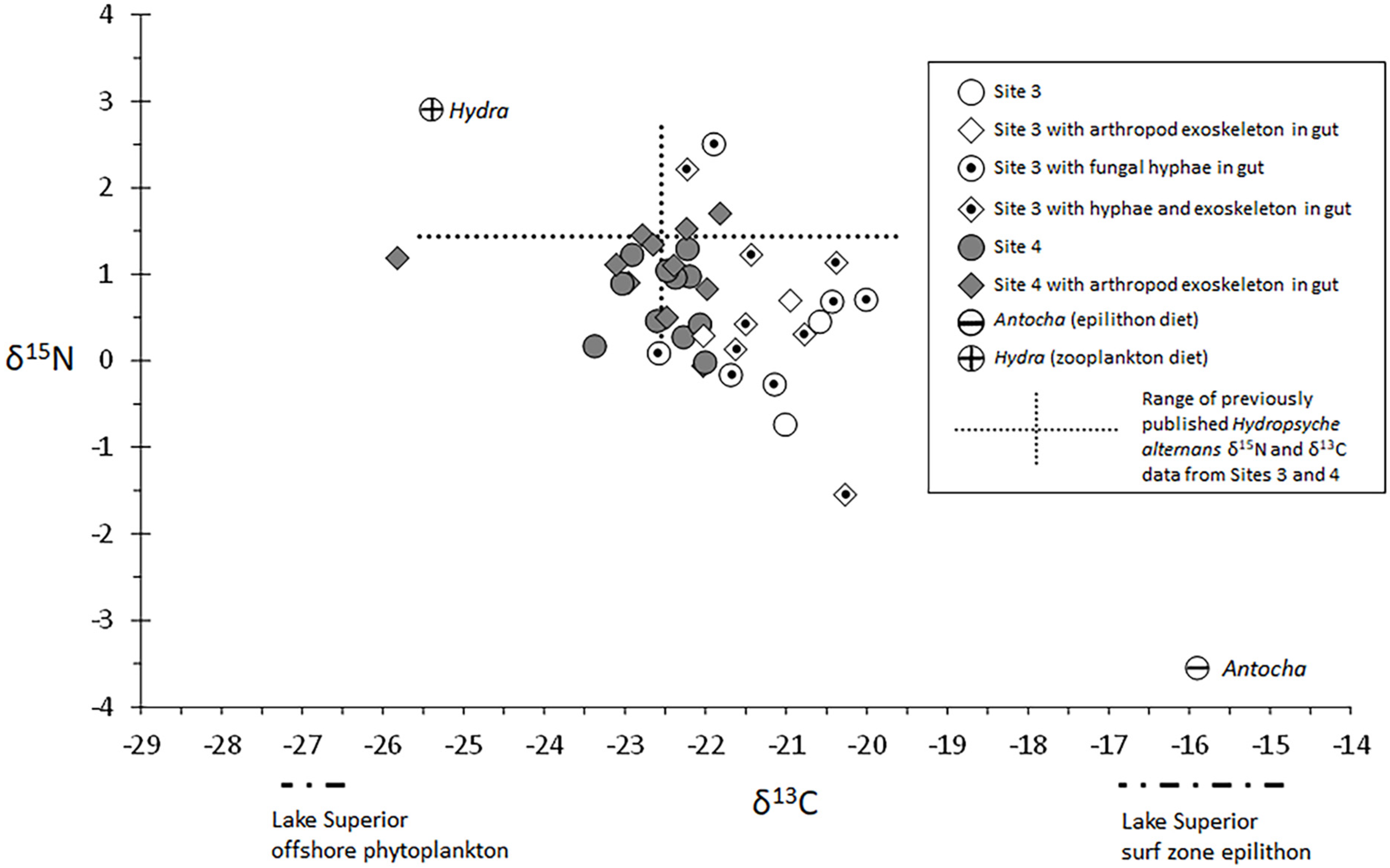
3.3. Gut Content Analyses
3.4. Stable Isotope Analyses
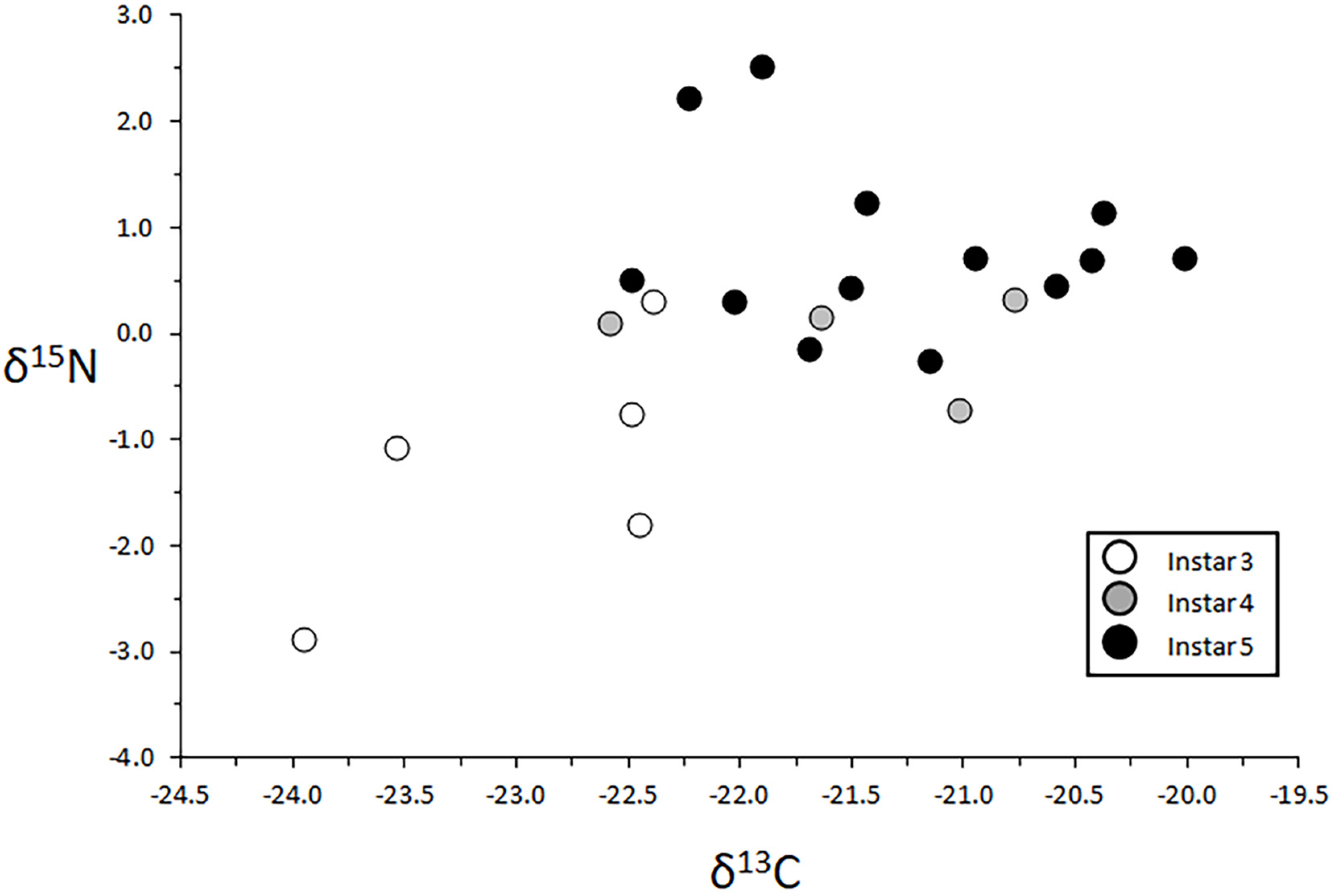
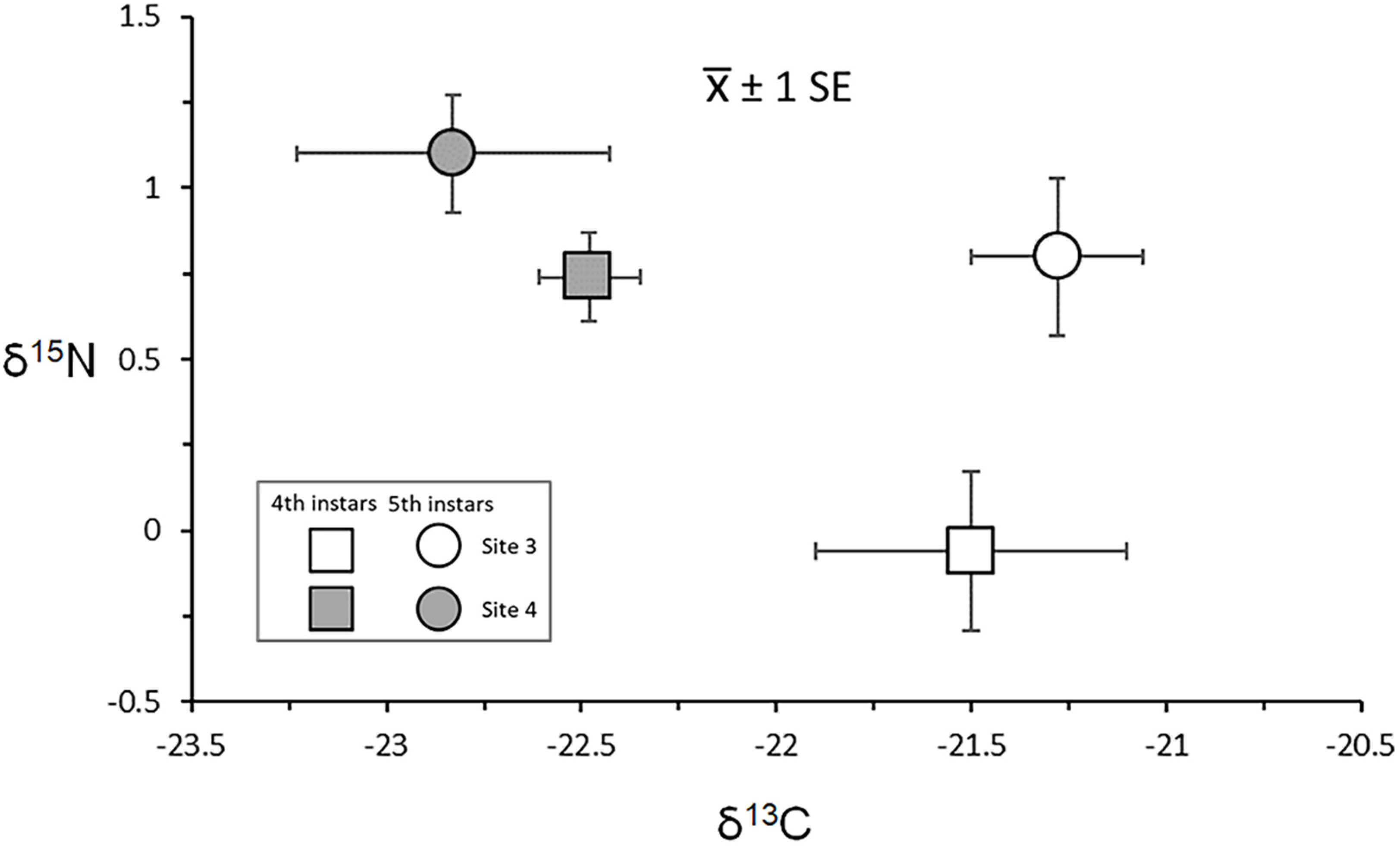
3.4.1. Larval Instar Effects
3.4.2. Intersite Variation and Gut Content Associations
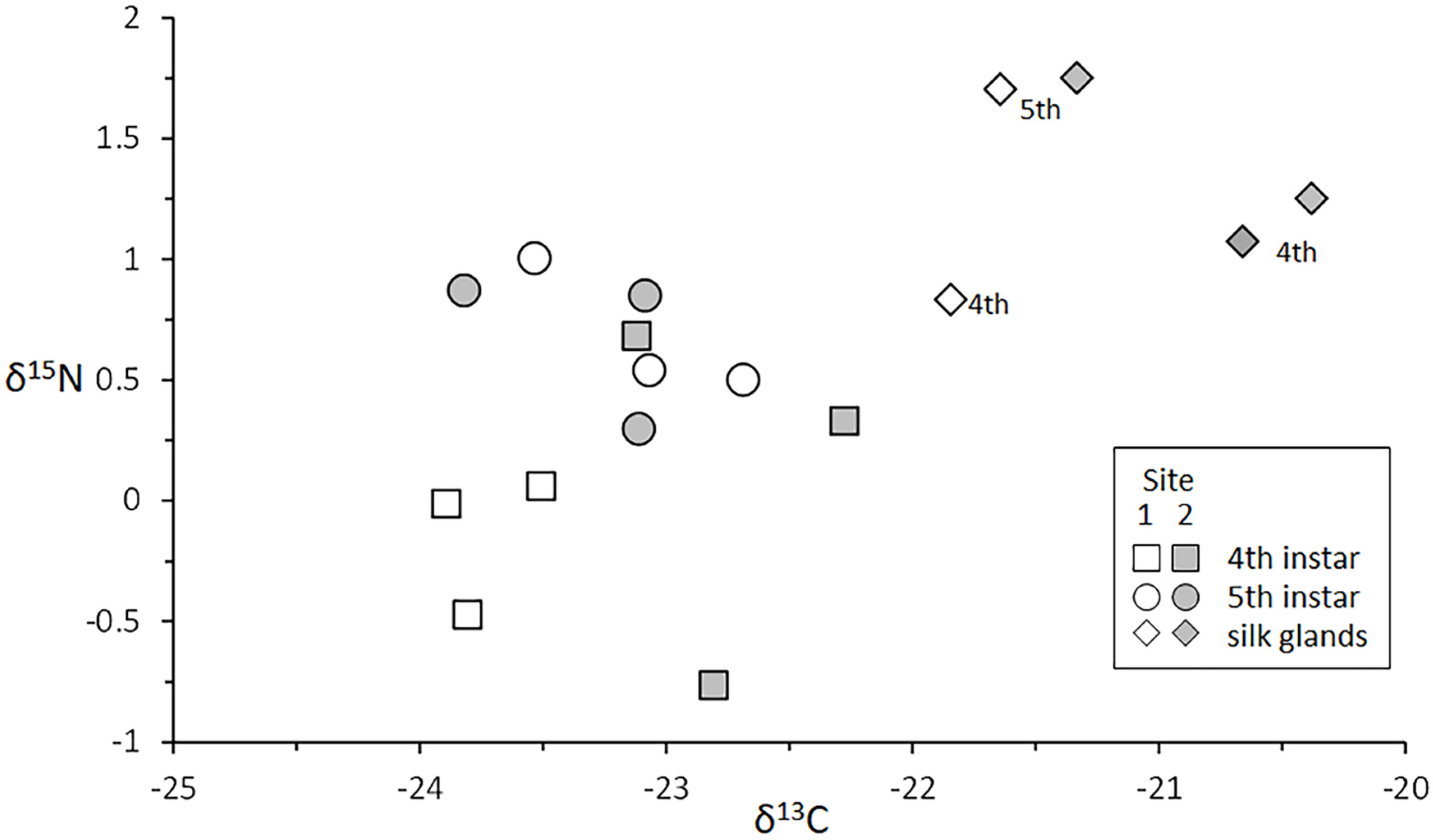
3.4.3. Silk Glands
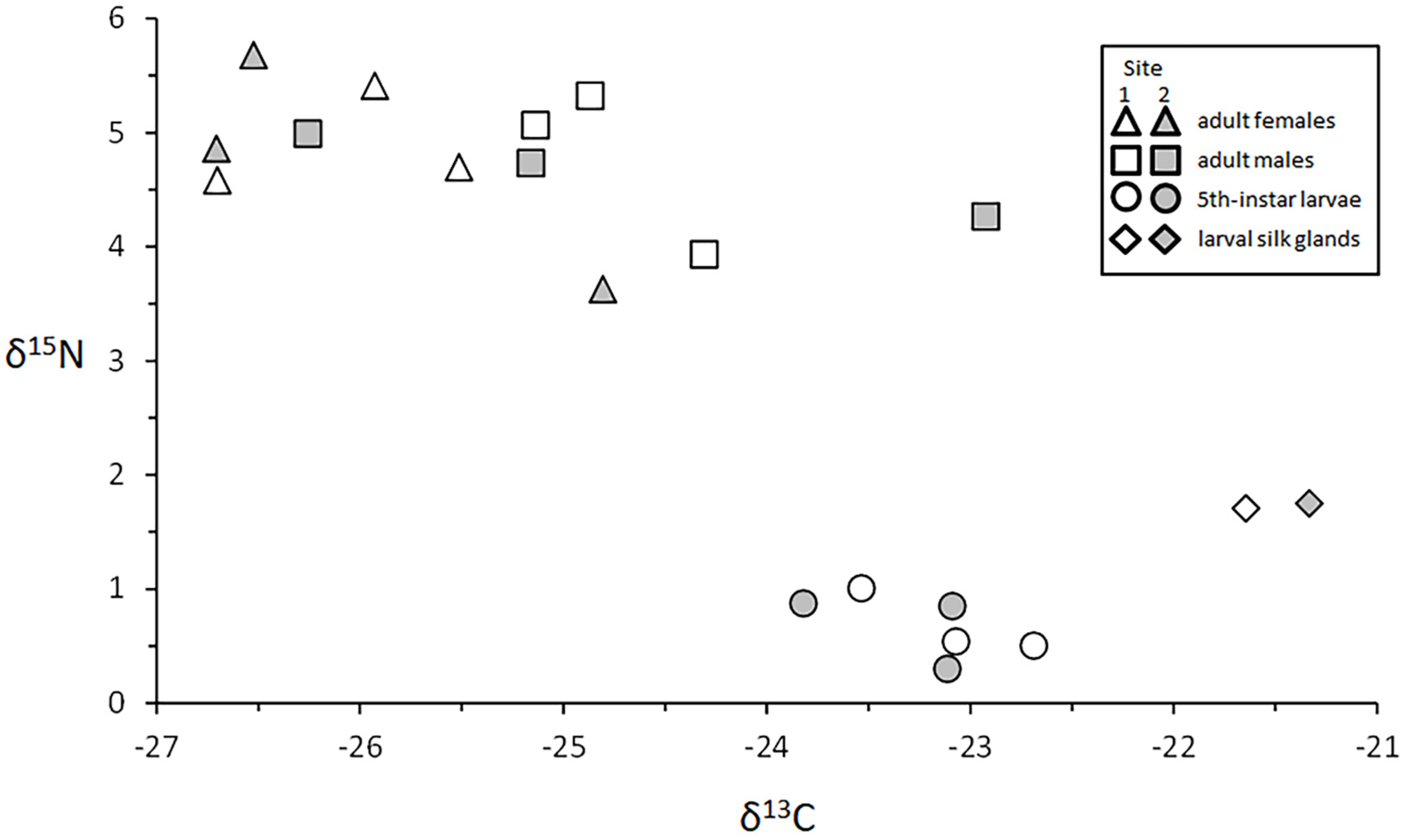
3.4.4. Adults
4. Discussion
4.1. Life History
4.2. Adult Characteristics
4.3. Gut Content Analyses
4.3.1. Carnivory
4.3.2. Mycophagy
4.4. Stable Isotope Analyses
4.4.1. δ13C
4.4.2. δ15N
4.4.3. Larval Silk Glands
4.4.4. Adults
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barton, D.R.; Hynes, H.B.N. Wave-zone Macrobenthos of the Exposed Canadian Shores of the St. Lawrence Great Lakes. J. Great Lakes Res. 1978, 4, 27–45. [Google Scholar] [CrossRef]
- Barton, D.R.; Smith, S.M. The Ecology of Aquatic Insects; Resh, V.H., Rosenberg, D.M., Eds.; Praeger: New York, NY, USA, 1984; pp. 456–483. [Google Scholar]
- Strand, M. Trophic Ecology of the Lake Superior Wave Zone: A Stable Isotope Approach. Hydrobiologia 2005, 544, 271–278. [Google Scholar] [CrossRef]
- Rao, Y.R.; Schwab, D.J. Transport and Mixing Between the Coastal and Offshore Waters in the Great Lakes: A Review. J. Great Lakes Res. 2007, 33, 202–218. [Google Scholar] [CrossRef] [Green Version]
- Herdendorf, C.E. Large Lakes of the World. J. Great Lakes Res. 1982, 8, 379–412. [Google Scholar] [CrossRef]
- Beletsky, D.; Saylor, J.H.; Schwab, D.J. Mean Circulation in the Great Lakes. J. Great Lakes Res. 1999, 25, 78–93. [Google Scholar] [CrossRef]
- Van Luven, D.M.; Huntoon, J.E.; Maclean, A.L. Determination of the Influence of Wind on the Keweenaw Current in the Lake Superior Basin as Identified by Advanced Very High Resolution Radiometer (AVHRR) Imagery. J. Great Lakes Res. 1999, 25, 625–641. [Google Scholar] [CrossRef]
- Chen, C.; Zhu, J.; Ralph, E.; Green, S.A.; Budd, J.W.; Zhang, F.Y. Prognostic Modeling Studies of the Keweenaw Current in Lake Superior. Part I: Formation and Evolution. J. Phys. Oceanogr. 2001, 31, 379–395. [Google Scholar] [CrossRef]
- Fenoglio, S.; Tierno de Figueroa, J.M.; Doretto, A.; Falasco, E.; Bona, F. Aquatic Insects and Benthic Diatoms: A History of Biotic Relationships in Freshwater Ecosystems. Water 2020, 12, 2934. [Google Scholar] [CrossRef]
- Fuller, R.J.; Mackey, R.J. Feeding Ecology of Three Species of Hydropsyche (Trichoptera: Hydropsychidae) in Southern Ontario. Can. J. Zool. 1980, 58, 2239–2251. [Google Scholar] [CrossRef]
- Sierszen, M.; Peterson, G.S.; Trebitz, A.S.; Brazner, J.C.; West, C.W. Hydrology and Nutrient Effects on Food-Web Structure in Ten Lake Superior Coastal Wetlands. Wetlands 2006, 26, 951–964. [Google Scholar] [CrossRef]
- Hynes, H.B.N. The Ecology of Running Waters; University of Toronto Press: Toronto, ON, Canada, 1970. [Google Scholar]
- Fuller, R.L.; Mackay, R.J. Effects of Food Quality on the Growth of Three Hydropsyche Species (Trichoptera: Hydropsychidae). Can. J. Zool. 1981, 59, 1133–1140. [Google Scholar] [CrossRef]
- Matczak, T.Z.; Mackay, R.J. Territoriality in filter-feeding caddisfly larvae: Laboratory experiments. J. N. Am. Benthol. Soc. 1990, 9, 26–34. [Google Scholar] [CrossRef]
- Oliveira, L.G.; Froehlich, C.G. Natural History of Three Hydropsychidae (Trichoptera, Insecta) in a “Cerrado” Stream from Northeastern Sao Paulo, Brazil. Revta Bras. Zool. 1996, 13, 755–762. [Google Scholar] [CrossRef]
- Sieglstetter, R.; Agasse, F.; Caquet, T. Ecological Segregation of Two Species of Hydropsyche (Trichoptera: Hydropsychidae) in a European Second-Order Stream (Essonne, France). J. Freshw. Ecol. 1997, 12, 269–279. [Google Scholar] [CrossRef]
- Hellmann, C.; Wissel, B.; Winkelmann, C. Omnivores as Seasonally Important Predators in a Stream Food Web. Freshw. Sci. 2013, 32, 548–562. [Google Scholar] [CrossRef]
- Peters, L.; Wetzel, M.A.; Rothaupt, K.O. Epilithic Communities in a Lake Littoral Zone: The Role of Water-Column Transport and Habitat Development for Dispersal and Colonization of Meiofauna. J. N. Am. Benthol. Soc. 2007, 26, 232–243. [Google Scholar] [CrossRef] [Green Version]
- Ferrier, M.D.; Wissing, T.E. A Scanning Electron Microscopic Study of the Mouthparts of Cheumatopsyche analis (Trichoptera: Hydropsychidae). Ohio J. Sci. 1980, 80, 38–40. [Google Scholar]
- Doucett, R.R.; Power, G.; Barton, D.R.; Drimmie, R.J.; Cunjak, R.A. Stable Isotope Analysis of Nutrient Pathways Leading to Atlantic Salmon. Can. J. Fish. Aquat. Sci. 1996, 53, 2058–2066. [Google Scholar] [CrossRef]
- Jardine, T.D.; Curry, R.A.; Heard, K.S.; Cunjak, R.A. High fidelity: Isotopic Relationship Between Stream Invertebrates and Their Gut Contents. J. N. Am. Benthol. Soc. 2005, 24, 90–299. [Google Scholar] [CrossRef]
- Ulseth, A.J.; Hershey, A.E. Natural Abundances of Stable Isotopes Trace Anthropogenic N and C in an Urban Stream. J. N. Am. Benthol. Soc. 2005, 24, 270–289. [Google Scholar] [CrossRef]
- Anderson, C.; Cabana, G. Estimating the Trophic Position of Aquatic Consumers in River Food Webs Using Stable Nitrogen Isotopes. J. N. Am. Benthol. Soc. 2007, 26, 273–285. [Google Scholar] [CrossRef]
- Ridzuan, D.S.; Salmah, C.; Rawi, M.; Hamid, S.A.; Al-Shami, S.A. Determination of Food Sources and Trophic Position in Malaysian Tropical Highland Streams Using Carbon and Nitrogen Stable Isotopes. Acta Ecol. Sin. 2017, 37, 97–104. [Google Scholar] [CrossRef]
- Benson, A.J. Quagga and Zebra Mussels: Biology, Impacts, and Control; Nalepa, T.E., Schloesser, D.W., Eds.; CRC Press: Boca Raton, FL, USA, 2013; pp. 9–31. [Google Scholar]
- Karatayev, A.Y.; Burlakova, L.E.; Pennuto, C.; Ciborowski, J.; VKaratayev, V.A.; Juette, P.; Clapsadl, M. Twenty Five Years of Changes in Dreissena spp. Populations in Lake Erie. J. Great Lakes Res. 2014, 40, 550–559. [Google Scholar] [CrossRef]
- Patterson, M.W.R.; Ciborowski, J.J.H.; Barton, D.R. The Distribution and Abundance of Dreissena Species (Dreissenidae) in Lake Erie. J. Great Lakes Res. 2005, 31, 223–237. [Google Scholar] [CrossRef]
- Nalepa, T.F.; Fanslow, D.L.; Lang, G.A.; Mabrey, K.; Rowe, M. Lake-Wide Benthic Surveys in Lake Michigan in 1994–95, 2000, 2005, and 2010: Abundances of the Amphipod Diporeia spp. and Abundances and Biomass of the Mussels Dreissena polymorpha and Dreissena rostriformis bugensis; NOAA Technical Memorandum GLERL-164; NOAA: Washington, DC, USA, 2014.
- Ratti, C.; Barton, D.R. Decline in the diversity of benthic invertebrates in the wave-zone of eastern Lake Erie, 1974–2001. J. Great Lakes Res. 2003, 29, 608–615. [Google Scholar] [CrossRef]
- Barton, D.R. Differences in Wave-Zone Benthic Invertebrate Communities in Lake Huron and Georgian Bay, 1974–2003. J. Great Lakes Res. 2004, 30, 508–518. [Google Scholar] [CrossRef]
- Ives, J.T.; McMeans, B.C.; McCann, K.S.; Fisk, A.T.; Johnson, T.B.; Bunnell, D.B.; Frank, K.T.; Muir, A.M. Food-Web Structure and Ecosystem Function in the Laurentian Great Lakes—Toward a Conceptual Model. Freshw. Biol. 2019, 64, 1–23. [Google Scholar] [CrossRef]
- Ross, H.H. The Caddis Flies, or Trichoptera, of Illinois. Bull. Ill. Nat. Hist. Surv. Div. 1944, 23, 1–326. [Google Scholar] [CrossRef]
- Schmude, K.L.; Hilsenhoff, W.L. Biology, Ecology, Larval Taxonomy, and Distribution of Hydropsychidae (Trichoptera) in Wisconsin. Great Lakes Entomol. 1986, 19, 123–145. [Google Scholar]
- Sykora, J.L.; Swegman, B.G.; Weaver, J.S. Occurrence of the Genus Hydropsyche in the North American Great Lakes. In Proceedings of the 3rd International Symposium on Trichoptera, Series Entomologica 20; Moretti, G.P., Ed.; Junk: The Hague, The Netherlands, 1981; Chapter 45; pp. 337–346. [Google Scholar]
- Williams, N.E.; Eyles, N. Sedimentary and Paleoclimatic Controls on Caddisfly (Insecta: Trichoptera) Assemblages During the Last Interglacial-to-Glacial Transition in Southern Ontario. Quat. Res. 1995, 43, 90–105. [Google Scholar] [CrossRef]
- Past Interglacials Working Group of PAGES. Interglacials of the last 800,000 years. Rev. Geophys. 2016, 54, 162–219. [Google Scholar] [CrossRef] [Green Version]
- Mann, J.D.; Leverington, D.W.; Rayburn, J.; Teller, J.T. The Volume and Paleobathymetry of Glacial Lake Agassiz. J. Paleolimnol. 1999, 22, 71–80. [Google Scholar] [CrossRef]
- Teller, J.T.; Leverington, D.W.; Mann, J.D. Freshwater Outbursts to the Oceans and Glacial Lake Agassiz and Their Role in Climate Change During the Last Deglaciation. Quat. Sci. Rev. 2002, 21, 879–887. [Google Scholar] [CrossRef]
- Teller, J.T.; Thorleifson, L.H. The Lake Agassiz-Lake Superior Connection. Geol. Assoc. Can. Spec. Pap. 1983, 26, 261–290. [Google Scholar]
- Leverington, D.W.; Teller, J.T. Paleotopographic Reconstructions of the Eastern Outlets of Glacial Lake Agassiz. Can. J. Earth Sci. 2003, 40, 1259–1278. [Google Scholar] [CrossRef] [Green Version]
- Patalas, K. Lake Winnipeg, Remnant of Glacial Lake Agassiz, an Efficient Dispersal Route of Planktonic Crustaceans through Central and Northern Canada. Aquat. Ecosyst. Health Manag. 2006, 9, 285–306. [Google Scholar] [CrossRef]
- Neave, F. A Contribution to the Aquatic Insect Fauna of Lake Winnipeg. Int. Rev. Gesamten Hydrobiol. Hydrogr. 1934, 31, 157–170. [Google Scholar] [CrossRef]
- Connallon, C.B.; Schaetzl, R.J. Geomorphology of the Chippewa River Delta of Glacial Lake Saginaw, Central Lower Michigan, USA. Geomorphology 2017, 290, 128–141. [Google Scholar] [CrossRef]
- Myers, L.W.; Kondratieff, B.C.; Mihuc, T.B.; Ruiter, D.E. The Mayflies (Ephemeroptera), Stoneflies (Plecoptera), and Caddisflies (Trichoptera) of the Adirondack Park (New York State). Trans. Am. Entomol. Soc. 2011, 137, 63–140. [Google Scholar] [CrossRef]
- Upham, W. The Glacial Lake Agassiz. Monographs of the United States Geological Survey 1895. Volume 25. 658p. Available online: https://commons.wikimedia.org/wiki/File:Agassiz.jpg (accessed on 8 October 2021).
- Leverington, D.W.; Mann, J.D.; Teller, J.T. Changes in the Bathymetry and Volume of Glacial Lake Agassiz between 11,000 and 9300 14C yr B.P. Quat. Res. 2000, 54, 174–181. [Google Scholar] [CrossRef] [Green Version]
- Houghton, D.C. Biological Diversity of Minnesota Caddisflies (Insecta: Trichoptera). Zoo Keys 2012, 189, 1–389. [Google Scholar] [CrossRef] [PubMed]
- Leonard, J.; Leonard, F. An Annotated List of Michigan Trichoptera; Occasional Papers of the Museum of Zoology; University of Michigan: Ann Arbor, MI, USA, 1949; 522p. [Google Scholar]
- Schefter, P.W.; Wiggins, G.B. A Systematic Study of the Hydropsyche Morosa Group (Trichoptera: Hydropsychidae); Royal Ontario Museum, Life Sciences Miscellaneous Publications: Toronto, ON, Canada, 1986.
- Sebastion, R.J.; Rosenberg, D.M.; Wiens, A.P. A Method for Subsampling Unsorted Benthic Macroinvertebrates by Weight. Hydrobiologia 1988, 157, 69–75. [Google Scholar] [CrossRef]
- Houghton, D.C.; DeWalt, R.E.; Pytel, A.J.; Brandin, C.M.; Rogers, S.E.; Ruiter, D.E.; Bright, E.; Hudson, P.L.; Armitage, B.J. Updated Checklist of the Michigan (USA) Caddisflies, with Regional and Habitat Affinities. Zoo Keys 2018, 730, 57–74. [Google Scholar] [CrossRef] [Green Version]
- NOAA Coastwatch Great Lakes. Available online: coastwatch.glerl.noaa.gov/modis (accessed on 8 October 2021).
- Google Earth. Available online: https://earth.google.com/web/ (accessed on 8 October 2021).
- Hester, F.E.; Dendy, J.S. A Multiple-Plate Sampler for Aquatic Macroinvertebrates. Trans. Am. Fish. Soc. 1962, 91, 420–421. [Google Scholar] [CrossRef] [Green Version]
- Ohio Environmental Protection Agency. Biological Criteria for the Protection of Aquatic Life: Vol III. Standardized Biological Field Sampling and Laboratory Methods for Assessing Fish and Macroinvertebrate Communities; Division of Water Quality Planning and Assessment, Ecological Assessment Section: Columbus, OH, USA, 1989. [Google Scholar]
- Schefter, P.W.; Wiggins, G.B.; Unzicker, J.D. A Proposal for Assignment of Ceratopsyche as a Subgenus of Hydropsyche, with New Synonyms and a New Species (Trichoptera: Hydropsychidae). J. N. Am. Benthol. Soc. 1986, 1, 67–84. [Google Scholar] [CrossRef]
- Dyar, H.G. The Number of Moults of Lepidopterous Larvae. Psyche 1890, 5, 420–422. [Google Scholar] [CrossRef]
- Mackay, R.J. Larval Identification and Instar Association in Some Species of Hydropsyche and Cheumatopsyche (Trichoptera: Hydropsychidae). Ann. Entomol. Soc. Am. 1978, 71, 499–509. [Google Scholar] [CrossRef]
- Waringer, J. Tools for Instar Determination of European Caddisfly Larvae (Insecta: Trichoptera). Zootaxa 2021, 4908, 85–101. [Google Scholar] [CrossRef]
- Deutsch, W.G. Swimming Modifications of Adult Female Hydropsychidae Compared with Other Trichoptera. Freshw. Invertebr. Biol. 1985, 4, 35–40. [Google Scholar] [CrossRef]
- Metcalfe, A.N.; Muehlbauer, J.D.; Kennedy, T.A.; Yackulic, C.B.; Dibble, K.L.; Marks, J.C. Net-Spinning Caddisfly Distribution in Large Regulated Rivers. Freshw. Biol. 2021, 66, 89–101. [Google Scholar] [CrossRef]
- Cummins, K.W. Trophic Relations of Aquatic Insects. Annu. Rev. Entomol. 1973, 18, 183–206. [Google Scholar] [CrossRef]
- Ashton, N.N.; Taggart, D.S.; Stewart, R.J. Silk Tape Nanostructure and Silk Gland Anatomy of Trichoptera. Biopolymers 2012, 97, 432–445. [Google Scholar] [CrossRef] [PubMed]
- Löfstedt, C.; Hansson, B.S.; Petersson, E.; Valeur, P.; Richards, A. Pheromonal Secretions from Glands on the 5th Abdominal Sternite of Hydropsychid and Rhyacophilid Caddisflies (Trichoptera). J. Chem. Ecol. 1994, 20, 153–170. [Google Scholar] [CrossRef]
- Willis, L.D.; Hendricks, A.C. Life History, Growth, Survivorship, and Production of Hydropsyche slossonae in Mill Creek, Virginia. J. N. Am. Benthol. Soc. 1992, 11, 290–303. [Google Scholar] [CrossRef]
- Keough, J.R.; Sierszen, M.E.; Hagley, C.A. Analysis of a Lake Superior coastal food wed with stable isotope techniques. Limnol. Oceanogr. 1996, 41, 136–146. [Google Scholar] [CrossRef]
- Rutherford, J.E.; Mackay, R.J. Variability in the Life-History Patterns of Four Species of Hydropsyche (Trichoptera: Hydropsychidae) in Southern Ontario Streams. Holarct. Ecol. 1986, 9, 149–163. [Google Scholar] [CrossRef]
- Rhame, R.E.; Stewart, K.W. Life Cycles and Food Habits of Three Hydropsychidae (Trichoptera) Species in the Brazos River, Texas. Trans. Am. Entomol. Soc. 1976, 102, 65–99. [Google Scholar]
- Huryn, A.D.; Wallace, J.B. Habitat, Life History, Secondary Production, and Behavioral Adaptations of Aquatic Insects. In An Introduction to the Aquatic Insects of North America, 4th ed.; Merritt, R.W., Cummins, K.W., Berg, M.B., Eds.; Kendall Hunt: Dubuque, IA, USA, 2019; pp. 65–116. [Google Scholar]
- Waters, T.F. Diurnal Periodicity in the Drift of Stream Invertebrates. Ecology 1962, 43, 316–320. [Google Scholar] [CrossRef]
- Elliot, J.M. The Life Histories and Drifting of Trichoptera in a Dartmoor Stream. J. Anim. Ecol. 1968, 37, 615–625. [Google Scholar] [CrossRef]
- Statzner, J.; Elouard, M.; Dejoux, C. Field Experiments on the Relationship Between Drift and Benthic Densities of Aquatic Insects in Tropical Streams (Ivory Coast). II. Cheumatopsyche falcifera (Trichoptera: Hydropsychidae). J. Anim. Ecol. 1986, 55, 93–110. [Google Scholar] [CrossRef]
- Tachet, H.; Pierrot, J.P.; Chantel, R.; Bournaud, M. Net-Building Behaviour of Six Hydropsyche Species (Trichoptera) in Relation to Current Velocity and Distribution along the Rhône River. J. N. Am. Benthol. Soc. 1992, 11, 350–365. [Google Scholar] [CrossRef]
- Ozersky, T.; Bramburger, A.J.; Elgin, A.K.; Vanderploeg, H.A.; Wang, J.; Austin, J.A. The Changing Face of Winter: Lessons and Questions from the Laurentian Great Lakes. J. Geophys. Res. Biogeosci. 2021, 126, 1–25. [Google Scholar] [CrossRef]
- Assel, R.J. Fall and Winter Thermal Structure of Lake Superior. J. Great Lakes Res. 1986, 12, 251–262. [Google Scholar] [CrossRef]
- Titze, D.J.; Austin, J.A. Winter Thermal Structure of Lake Superior. Limnol. Oceanogr. 2014, 59, 1336–1348. [Google Scholar] [CrossRef]
- Kovalenko, K.E.; Reavie, E.D.; Bramburger, A.J.; Cotter, A.; Sierszen, M.E. Nearshore-offshore trends in Lake Superior phytoplankton. J. Great Lakes Res. 2019, 45, 1197–1204. [Google Scholar] [CrossRef]
- Vanderploeg, H.A.; Cavaletto, J.F.; Liebig, J.F.; Gardner, W.S. Limnocalanus macrurus (Copepoda: Calanoida) Retains a Marine Arctic Lipid and Life Cycle Strategy in Lake Michigan. J. Plankton Res. 1998, 20, 1581–1597. [Google Scholar] [CrossRef] [Green Version]
- Badcock, R.M. Observation of Oviposition Under Water of the Aerial Insect Hydropsyche angustipennis (Curtis) (Trichoptera). Hydrobiologia 1953, 5, 222–225. [Google Scholar] [CrossRef]
- Kjær, C.; Sørensen, P.B.; Wiberg-Larsen, P.; Bak, J.; Bruus, M.; Strandberg, B.; Larsen, S.E.; Rasmussen, J.J.; Strandberg, M. Vulnerability of Aquatic Insect Species to Insecticides, Depending on their Flight Period and Adult Life Span. Environ. Toxicol. Chem. 2021, 40, 1778–1787. [Google Scholar] [CrossRef]
- Kovats, Z.; Ciborowski, J.H.; Corkum, L.D. Inland Dispersal of Adult Aquatic Insects. Freshw. Biol. 1996, 36, 265–276. [Google Scholar] [CrossRef] [Green Version]
- Sode, A.; Wiberg-Larsen, P. Dispersal of Adult Trichoptera at a Danish Forest Brook. Freshw. Biol. 1993, 30, 439–446. [Google Scholar] [CrossRef]
- Fremling, C.R. Biology and Possible Control of Nuisance Caddisflies of the Upper Mississippi River. Iowa Agric. Home Econ. Exp. Stn. Res. Bull. 1960, 33, 856–879. [Google Scholar]
- Wallace, J.B. The Larval Retreat and Food of Arctopsyche; with Phylogenetic Notes on Feeding Adaptations in Hydropsychidae Larvae (Trichoptera). Ann. Entomol. Soc. Am. 1975, 68, 167–173. [Google Scholar] [CrossRef]
- Hoagland, K.D.; Zlotsky, A.; Peterson, C.G. Algal Biofouling; Evans, L.V., Hoagland, K.D., Eds.; Elsevier Science Publishing: Amsterdam, The Netherlands, 1986; pp. 21–39. [Google Scholar]
- Barton, D.R.; Carter, J.C.H. Shallow-Water Epilithic Invertebrate Communities of Eastern Georgian Bay, Ontario, in Relation to Exposure to Wave Action. Can. J. Zool. 1982, 60, 984–993. [Google Scholar] [CrossRef]
- Petersen, L.B.M. Food Preferences in Three Species of Hydropsyche (Trichoptera). Verh. Int. Ver. Limnol. 1985, 22, 3270–3274. [Google Scholar] [CrossRef]
- Benke, A.C.; Wallace, J.B. Trophic Basis of Production among Net-Spinning Caddisflies in a Southern Appalachian Stream. Ecology 1980, 61, 108–118. [Google Scholar] [CrossRef]
- Benke, A.C.; Wallace, J.B. Trophic Basis of Production among Riverine Caddisflies: Implications for Food Web Analysis. Ecology 1997, 78, 1132–1145. [Google Scholar] [CrossRef]
- Benke, A.C.; Wallace, J.B.; Harrison, J.W.; Koebel, J.W. Food Web Quantification using Secondary Production Analysis: Predaceous Invertebrates of the Snag Habitat in a Subtropical River. Freshw. Biol. 2001, 46, 329–346. [Google Scholar] [CrossRef]
- Koske, R.E.; Polson, W.R. Are VA Mycorrhizae Required for Sand Dune Stabilization? Bioscience 1984, 34, 420–424. [Google Scholar] [CrossRef]
- Koske, R.E.; Gemma, J.N. Mycorrhizae and Succession in Plantings of Beach Grass in Sand Dunes. Am. J. Bot. 1997, 84, 118–130. [Google Scholar] [CrossRef]
- Bago, B.; Zipfel, W.; Williams, R.M.; Jun, J.; Arreola, R.; Lammers, P.J.; Pfeffer, P.E.; Shachar-Hill, Y. Translocation and Utilization of Fungal Storage Lipid in the Arbuscular Mycorrhizal Symbiosis. Plant Physiol. 2002, 128, 108–124. [Google Scholar] [CrossRef]
- Gleixner, G.; Danier, H.J.; Werner, R.A.; Schmidt, H.L. Correlations Between the 13C Content of Primary and Secondary Plant Products in Different Cell Compartments and that in Decomposing Basidiomycetes. Plant Physiol. 1993, 102, 1287–1290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trudell, S.A.; Rygiewicz, P.T.; Edmonds, R.L. Patterns of Nitrogen and Carbon Stable Isotope Ratios in Macrofungi, Plants and Soils in Two Old-Growth Conifer Forests. New Phytol. 2004, 164, 317–335. [Google Scholar] [CrossRef] [PubMed]
- Klink, S.; Giesemann, P.; Hubmann, T.; Pausch, J. Stable C and N Isotope Natural Abundances of Intraradical Hyphae of Arbuscular Mycorrhizal Fungi. Mycorrhiza 2020, 30, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Colombini, I.; Brilli, M.; Fallaci, M.; Gagnarli, E.; Chelazzi, L. Food Webs of a Sandy Beach Macroinvertebrate Community using Stable Isotopes Analysis. Acta Oecol. 2011, 37, 422–432. [Google Scholar] [CrossRef]
- Harris, A.L.; Zinnert, J.C.; Young, D.R. Differential Response of Barrier Island Dune Grasses to Species Interactions and Burial. Plant Ecol. 2017, 218, 609–619. [Google Scholar] [CrossRef] [Green Version]
- Powers, M.D.; Webster, C.R.; Pregitzer, K.S.; Palik, B.J. Spatial Dynamics of Radial Growth and Growth Efficiency in Residual Pinus resinosa Following Aggregated Retention Harvesting. Can. J. For. Res. 2009, 39, 109–117. [Google Scholar] [CrossRef] [Green Version]
- Melillo, J.M.; Aber, J.D.; Linkins, A.E.; Ricca, A.; Fry, B.; Nadelhoffer, K.J. Carbon and Nitrogen Dynamics along the Decay Continuum: Plant litter to Soil Organic Matter. Plant Soil 1989, 115, 189–198. [Google Scholar] [CrossRef]
- Sutherland, T.D.; Young, J.H.; Weisman, S.; Hayashi, C.Y.; Merritt, D.J. Insect Silk: One Name Many Materials. Annu. Rev. Entomol. 2010, 55, 171–188. [Google Scholar] [CrossRef]
- Wallace, J.B. Silk Spinning as an Escape Mechanism in Hydropsyche orris Larvae Following Removal from Water (Trichoptera: Hydropsychidae). Ann. Entomol. Soc. Am. 1975, 68, 549–550. [Google Scholar] [CrossRef]
- Sehnal, F.; Sutherland, T. Silks Produced by Insect Labial Glands. Prion 2008, 2, 145–153. [Google Scholar] [CrossRef] [Green Version]
- Steffan, S.A.; Chikaraishi, Y.; Horton, D.R.; Ohkouchi, N.; Singleton, M.E. Trophic Hierarchies Illuminated via Amino Acid Isotopic Analysis. PLoS ONE 2013, 8, e76152. [Google Scholar] [CrossRef] [PubMed]
- Takizawa, Y.; Takano, Y.; Choi, B.; Dharampa, P.S.; Steffan, S.A.; Ogawa, N.O.; Ohkouchi, N.; Chikaraishi, Y. A New Insight into Isotopic Fractionation Associated with Decarboxylation in Organisms: Implications for Amino Acid Isotope Approaches in Biogeoscience. Prog. Earth Planet. Sci. 2020, 7, 50. [Google Scholar] [CrossRef]
- Arrese, E.L.; Canavoso, L.E.; Jouni, Z.E.; Pennington, J.E.; Tsuchida, K.; Wells, M.A. Lipid Storage and Mobilization in Insects: Current Status and Future Directions. Insect Biochem. Mol. Biol. 2001, 31, 7–17. [Google Scholar] [CrossRef]
- Bartrons, M.; Grimalt, J.O.; Catalan, J. Concentration Changes of Organochlorine Compounds and Polybromodiphenyl Ethers during Metamorphosis of Aquatic Insects. Environ. Sci. Technol. 2007, 41, 6137–6141. [Google Scholar] [CrossRef]
- Tibbets, T.M.; Wheeless, L.A.; Martínez del Rio, C. Isotopic Enrichment Without Change in Diet: An Ontogenetic Shift in δ15N During Insect Metamorphosis. Funct. Ecol. 2008, 22, 109–113. [Google Scholar] [CrossRef]
- Post, D.M.; Layman, C.A.; Arrington, D.A.; Takimoto, G.; Quattrochi, J.; Montana, C.G. Getting to the Fat of the Matter: Models, Methods and Assumptions for Dealing with Lipids in Stable Isotope Analyses. Oecologia 2007, 152, 179–189. [Google Scholar] [CrossRef]
- Smith, S.I. Sketch of the Invertebrate Fauna of Lake Superior; Report of Commissioner of Fish and Fisheries XXVII; U.S. Commission on Fish and Fisheries: Washington, DC, USA, 1873.
- Wiedmer, M.; Montgomery, D.R.; Gillespie, A.R.; Greenberg, H. Late Quaternary Megafloods from Glacial Lake Atna, Southcentral Alaska, U.S.A. Quat. Res. 2010, 73, 413–424. [Google Scholar] [CrossRef]
- Woolway, R.L.; Kraemer, B.M.; Lenters, J.D.; Merchant, C.J.; O’Reilly, C.M.; Sharma, S. Global Lake Responses to Climate Change. Nat. Rev. Earth Environ. 2020, 1, 388–403. [Google Scholar] [CrossRef]
- Austin, J.A.; Colman, S.M. Lake Superior Summer Water Temperatures are Increasing More Rapidly than Regional Air Temperatures: A Positive Ice-Albedo Feedback. Geophys. Res. Lett. 2007, 34, 1–5. [Google Scholar] [CrossRef] [Green Version]
- Ye, X.; Anderson, E.J.; Chu, P.Y.; Huang, C.; Xue, P. Impact of Water Mixing and Ice Formation on the Warming of Lake Superior: A Model-Guided Mechanism Study. Limnol. Oceanogr. 2019, 64, 558–574. [Google Scholar] [CrossRef]
- Sterner, R.S.; Rein, K.; Lafrancois, B.M.; Brovold, S.; Miller, T.R. A First Assessment of Cyanobacterial Blooms in Oligotrophic Lake Superior. Limnol. Oceanogr. 2020, 65, 2984–2998. [Google Scholar] [CrossRef]
- Trebitz, A.T.; Hatzenbuhler, C.L.; Hoffman, J.C.; Meredith, C.S.; Peterson, G.S.; Pilgrim, E.M.; Barge, J.T.; Cotter, A.M.; Wick, M.J. Dreissena veligers in western Lake Superior—Inference from New Low-Density Detection. J. Great Lakes Res. 2019, 45, 691–699. [Google Scholar] [CrossRef]
- Grigorovich, I.A.; Korniushin, A.V.; Gray, D.K.; Duggan, I.C.; Colutti, R.I.; MacIsaac, H.J. Lake Superior: An Invasion Coldspot? Hydrobiologia 2003, 499, 191–210. [Google Scholar] [CrossRef]
- Grigorovich, I.A.; Kelly, J.R.; Darling, J.A.; West, C.W. The Quagga Mussel Invades Lake Superior. J. Great Lakes Res. 2008, 34, 342–350. [Google Scholar] [CrossRef]
- Handfield, L.; Handfield, D. The Curious, and Incorrect, Case of “St. Martin’s Falls”, a Type Locality for Many Insect Species Described by Francis Walker. Zootaxa 2020, 4786, 437–443. [Google Scholar] [CrossRef]
- Banks, N. American Trichoptera—Notes and Descriptions. Can. Entomol. 1914, 46, 149–156. [Google Scholar] [CrossRef]
- Betten, C. The Caddis Flies or Trichoptera of New York State. N. Y. State Mus. Bull. 1934, 292. [Google Scholar]
- Ross, H.H. Lectotypes of North American Caddis Flies in the Museum of Comparative Zoology. Psyche 1938, 45, 1–61. [Google Scholar] [CrossRef] [Green Version]
- Banks, N. Descriptions of New Nearctic Neuropteroid Insects. Trans. Am. Entomol. Soc. 1905, 32, 1–20. [Google Scholar]
- Nimmo, A.P. Francis Walker Types of, and New Synonymies for, North American Hydropsyche Species. Psyche 1981, 88, 259–263. [Google Scholar] [CrossRef] [Green Version]
- Vorhies, C.T. Studies on the Trichoptera of Wisconsin. Trans. Wis. Acad. Sci. Arts Lett. 1909, 6, 647–738. [Google Scholar]
- Schuster, G.A.; Etnier, D.A. A Manual for the Identification of the Larvae of the Caddisfly Genera Hydropsyche Pictet and Symphitopsyche Ulmer in Eastern and Central North America (Trichoptera: Hydropsychidae); 600/4-78-060; United States Environmental Protection Agency: Washington, DC, USA, 1978; 141p.
- Nielson, A. On the evolution of the phallus and other male terminalia in the Hydropsychidae with a proposal for a new generic name. In Proceedings of the 3rd International Symposium on Trichoptera, Series Entomologica 20; Moretti, G.P., Ed.; Junk: The Hague, The Netherlands, 1981; Chapter 34; pp. 273–278. [Google Scholar]
- Morse, J.C. A Checklist of the Trichoptera of North America, Including Greenland and Mexico. Trans. Am. Entomol. Soc. 1993, 119, 47–93. [Google Scholar]
- Wiggins, G.B. Trichoptera families. In An Introduction to the Aquatic Insects of North America, 3rd ed.; Merritt, R.W., Cummins, K.W., Eds.; Kendall Hunt: Dubuque, IA, USA, 1996; pp. 309–349. [Google Scholar]
- Morse, J.C.; Holzenthal, R.W. Trichoptera genera. In An Introduction to the Aquatic Insects of North America, 3rd ed.; Merritt, R.W., Cummins, K.W., Eds.; Kendall Hunt: Dubuque, IA, USA, 1996; pp. 350–386. [Google Scholar]
- Morse, J.C.; Holzenthal, R.W.; Robertson, D.R.; Rasmussen, A.K.; Currie, D.C. Trichoptera. In An Introduction to the Aquatic Insects of North America, 4th ed.; Merritt, R.W., Cummins, K.W., Berg, M.B., Eds.; Kendall Hunt: Dubuque, IA, USA, 2019; pp. 481–552. [Google Scholar]
- Wiggins, G.B. Larvae of the North American Caddisfly Genera (Trichoptera), 2nd ed.; University of Toronto Press: Toronto, ON, Canada, 2000. [Google Scholar]
- Geraci, C.J.; Zhou, X.; Morse, J.C.; Kjer, K.J. Defining the genus Hydropsyche (Trichoptera: Hydropsychidae) Based on DNA and Morphological Evidence. J. N. Am. Benthol. Soc. 2010, 29, 918–933. [Google Scholar] [CrossRef] [Green Version]
- Cohen, J.G.; Kost, M.A.; Slaughter, B.S.; Albert, D.A.; Lincoln, J.M.; Kortenhoven, A.P.; Wilton, C.M.; Enander, H.D.; Korroch, K.M. Michigan Natural Community Classification [Web Application]; Michigan Natural Features Inventory, Michigan State University Extension: Lansing, MI, USA, 2020; Available online: https://mnfi.anr.msu.edu/communities/classification (accessed on 8 October 2021).
- Anderton, J.B. The Jewel in the Crown: An Environmental History of Presque Isle Park, Marquette, Michigan; Center for Upper Peninsula Studies, Northern Michigan University: Marquette, MI, USA, 2009; 40p. [Google Scholar]
- Ross, H.H. The Quaternary of the United States; Wright, H.E., Jr., Frey, D.G., Eds.; Princeton University Press: Princeton, NJ, USA, 1965; pp. 583–596. [Google Scholar]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Algae (Diatoms and Green Algae) | Animal (Arthropod Exoskeleton Fragments) | Fungi (Hyphae) | Sediment (Fine Particles) | |
|---|---|---|---|---|
| Site 3 | 94 | 50 | 78 | 72 |
| Site 4 | 95 | 48 | 0 | 57 |
| Sample Type | Site | n | Mean δ13C ± 1SE | Low δ13C | High δ13C | Mean δ15N ± 1SE | Low δ15N | High δ15N |
|---|---|---|---|---|---|---|---|---|
| 3rd instar | 3 | 5 | −22.95 ± 0.33 | −23.94 | −22.38 | −1.26 ± 0.53 | −2.90 | 0.29 |
| 4th instar | 3 | 4 | −21.50 ± 0.40 | −22.58 | −20.77 | −0.06 ± 0.23 | −0.74 | 0.30 |
| 4th instar | 4 | 12 | −22.48 ± 0.13 | −23.37 | −22.00 | 0.74 ± 0.13 | −0.02 | 1.29 |
| 5th instar | 3 | 13 | –21.28 ± 0.22 | −22.48 | −20.01 | 0.80 ± 0.23 | −0.28 | 2.50 |
| 5th instar | 4 | 9 | −22.83 ± 0.40 | −25.82 | −21.82 | 0.23 vs. 1.10 | −0.06 | 1.70 |
| 4th instar | 1 | 3 | −23.74 ± 0.12 | −23.89 | −23.51 | −0.14 ± 0.17 | −0.47 | 0.06 |
| 4th instar silk glands | 1 | 1 (n = 3) | −21.85 | 0.83 | ||||
| 4th instar | 2 | 3 | −22.73 ± 0.25 | −23.12 | −22.28 | 0.08 ± 0.44 | −0.77 | 0.68 |
| 4th instar silk glands | 2 | 1 (n = 3) | −20.38 | 1.25 | ||||
| 4th instar silk glands | 2 | 1 (n = 3) | −20.66 | 1.07 | ||||
| 5th instar | 1 | 3 | −23.10 ± 0.25 | −23.54 | −22.69 | 0.68 ± 0.16 | 0.50 | 1.01 |
| 5th instar silk glands | 1 | 1 (n = 3) | −21.64 | 1.70 | ||||
| 5th instar | 2 | 3 | −23.34 ± 0.24 | −23.82 | −23.11 | 0.67 ± 0.19 | 0.30 | 0.87 |
| 5th instar silk glands | 2 | 1 (n = 3) | −21.33 | 1.75 | ||||
| adult female | 1 | 3 | −26.05 ± 0.35 | −26.70 | −25.51 | 4.89 ± 0.26 | 4.54 | 5.41 |
| adult female | 2 | 3 | −26.01 ± 0.61 | −26.52 | −24.80 | 4.72 ± 0.60 | 3.63 | 5.68 |
| adult male | 1 | 3 | −24.77 ± 0.24 | −25.14 | −24.31 | 4.77 ± 0.43 | 3.93 | 5.32 |
| adult male | 2 | 3 | −24.78 ± 0.98 | −26.26 | −22.92 | 4.66 ± 0.21 | 4.26 | 4.99 |
| Contrast | p-Value δ13C | p-Value δ15N |
|---|---|---|
| Instar 3 × Instar 4 | 0.0271 | 0.1235 |
| Instar 3 × Instar 5 | 0.0027 | 0.0007 |
| Instar 4 × Instar 5 | 0.9482 | 0.2251 |
| Contrast | p-Value δ13C | p-Value δ15N |
|---|---|---|
| Site 3 Instar 4 × Site 3 Instar 5 | 0.9683 | 0.0958 |
| Site 3 Instar 4 × Site 4 Instar 4 | 0.1817 | 0.1384 |
| Site 3 Instar 4 × Site 4 Instar 5 | 0.0486 | 0.0189 |
| Site 3 Instar 5 × Site 4 Instar 4 | 0.0047 | 0.9956 |
| Site 3 Instar 5 × Site 4 Instar 5 | 0.0006 | 0.6686 |
| Site 4 Instar 4 × Site 4 Instar 5 | 0.7615 | 0.5513 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Miess, S.; Chrisekos, A.; Strand, M. An Ecological Profile of Hydropsyche alternans (Trichoptera: Hydropsychidae) in Lake Superior, the Last Stronghold of a Once-Dominant Great Lakes Surf Zone Caddisfly. Insects 2022, 13, 659. https://doi.org/10.3390/insects13070659
Miess S, Chrisekos A, Strand M. An Ecological Profile of Hydropsyche alternans (Trichoptera: Hydropsychidae) in Lake Superior, the Last Stronghold of a Once-Dominant Great Lakes Surf Zone Caddisfly. Insects. 2022; 13(7):659. https://doi.org/10.3390/insects13070659
Chicago/Turabian StyleMiess, Sam, Alissa Chrisekos, and Mac Strand. 2022. "An Ecological Profile of Hydropsyche alternans (Trichoptera: Hydropsychidae) in Lake Superior, the Last Stronghold of a Once-Dominant Great Lakes Surf Zone Caddisfly" Insects 13, no. 7: 659. https://doi.org/10.3390/insects13070659