
Characterization of Chitin Synthase A cDNA from Diaphorina citri (Hemiptera: Liviidae) and Its Response to Diflubenzuron

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect
2.2. Leaf-Dip Bioassay
2.3. Total RNA Extraction and cDNA Synthesis
2.4. Cloning and Sequencing of DcCHSA
2.5. Real-Time Quantitative PCR (RT-qPCR)
2.6. Preparation of dsRNA and Feeding
2.7. Bioinformatic and Phylogenetic Analyses and Statistical Analysis
2.8. Statistical Analysis
3. Results
3.1. Cloning and Sequence Analysis of DcCHSA
3.2. Phylogenetic Analysis of DcCHSA
3.3. Gene Expression Profiles of DcCHSA
3.4. Effect of DFB on D. citri Survival and DcCHSA Expression Level
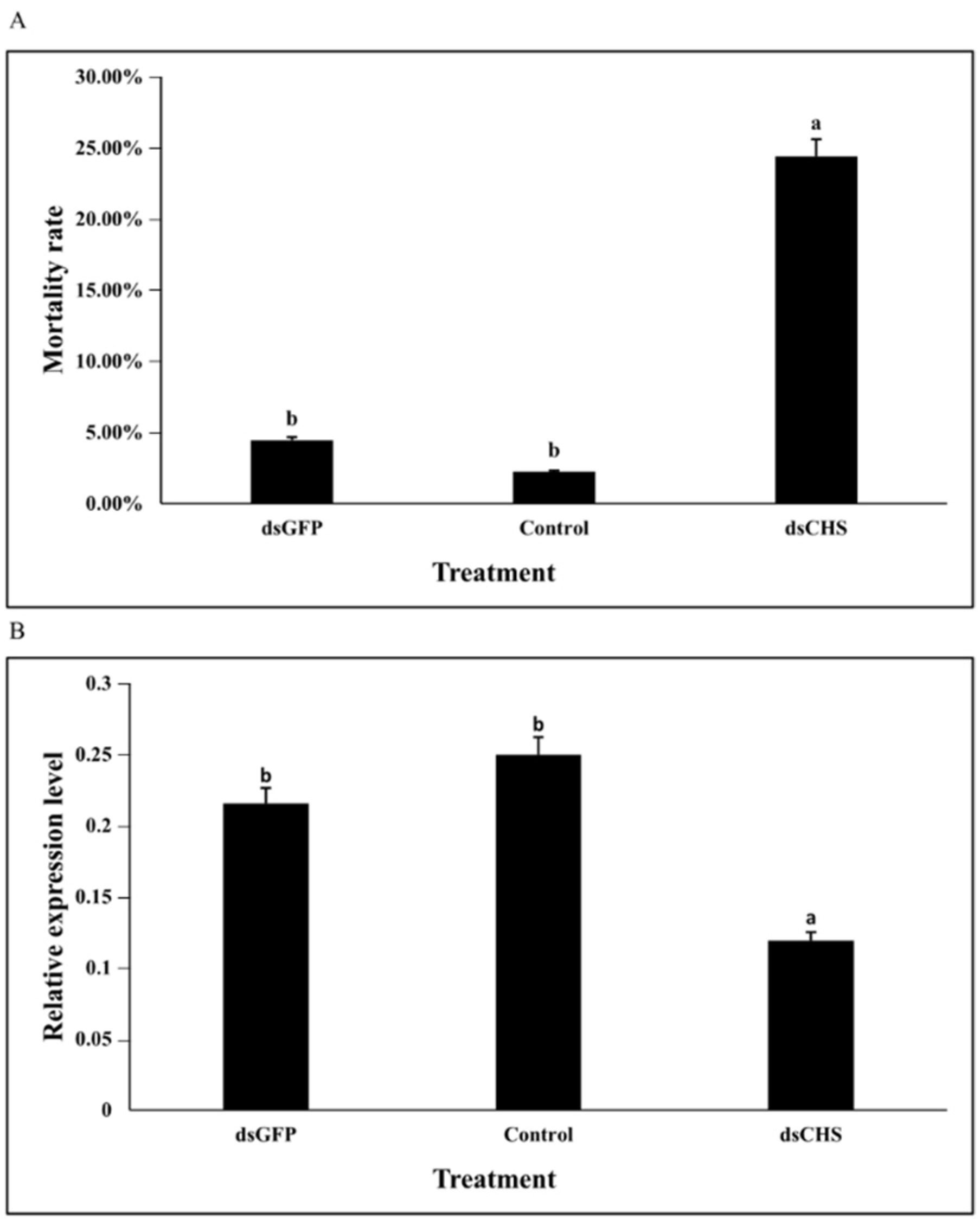
3.5. DcCHSA RNAi Analysis
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dong, M.; Wen, G.; Tang, H.; Wang, T.; Zhao, Z.; Song, W.; Wang, W.; Zhao, L. Dissipation and safety evaluation of novaluron, pyriproxyfen, thiacloprid and tolfenpyrad residues in the citrus-field ecosystem. Food Chem. 2018, 269, 136–141. [Google Scholar] [CrossRef]
- Ruiz, M.J.; Juárez, M.L.; Alzogaray, R.A.; Arrighi, F.; Arroyo, L.; Gastaminza, G.; Willink, E.; Bardón, A.; Vera, M.T. Oviposition behaviour and larval development of Anastrepha fraterculus from Argentina in citrus. Entomol. Exp. Appl. 2015, 157, 198–213. [Google Scholar] [CrossRef]
- Sun, D.; Zhu, Y.; Pang, J.; Zhou, Z.; Jiao, B. Residue level, persistence and safety of spirodiclofen-pyridaben mixture in citrus fruits. Food Chem. 2016, 194, 805–810. [Google Scholar] [CrossRef]
- Chu, C.C.; Hoffmann, M.; Braswell, W.E.; Pelz Stelinski, K.S. Genetic variation and potential coinfection of Wolbachia among widespread Asian citrus psyllid (Diaphorina citri Kuwayama) populations. Insect Sci. 2019, 26, 671–682. [Google Scholar] [CrossRef] [PubMed]
- Halbert, S.; Manjunath, K. Asian citrus psyllids (Sternorrhyncha: Psyllidae) and greening disease of citrus: A literature review and assessment of risk in Florida. Fla. Entomol. 2004, 3, 330–353. [Google Scholar] [CrossRef]
- Yang, Y.; Huang, M.; Beattie, G.A.C.; Xia, Y.; Ouyang, G.; Xiong, J. Distribution, biology, ecology and control of the psyllid Diaphorina citri Kuwayama, a major pest of citrus: A status report for China. Int. J. Pest Manag. 2006, 52, 343–352. [Google Scholar] [CrossRef]
- Zhou, C. The status of citrus Huanglongbing in China. Trop. Plant Pathol. 2020, 45, 279–284. [Google Scholar] [CrossRef]
- Wang, R.; Yang, H.; Luo, W.; Wang, M.; Lu, X.; Huang, T.; Zhao, J.; Li, Q. Predicting the potential distribution of the Asian citrus psyllid, Diaphorina citri (Kuwayama), in China using the MaxEnt model. PeerJ 2019, 7, e7323. [Google Scholar] [CrossRef]
- Merzendorfer, H.; Zimoch, L. Chitin metabolism in insects: Structure, function and regulation of chitin synthases and chitinases. J. Exp. Biol. 2003, 206, 4393–4412. [Google Scholar] [CrossRef]
- Merzendorfer, H. Insect chitin synthases: A review. J. Comp. Physiol. B 2006, 176, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Bansal, R.; Mian, M.A.R.; Mittapalli, O.; Michel, A.P. Characterization of a Chitin Synthase Encoding Gene and Effect of Diflubenzuron in Soybean Aphid, Aphis glycines. Int. J. Biol. Sci. 2012, 8, 1323–1334. [Google Scholar] [CrossRef]
- Zhuo, W.; Fang, Y.; Kong, L.; Li, X.; Sima, Y.; Xu, S. Chitin synthase A: A novel epidermal development regulation gene in the larvae of Bombyx mori. Mol. Biol. Rep. 2014, 41, 4177–4186. [Google Scholar] [CrossRef]
- Toprak, U.; Erlandson, M.; Baldwin, D.; Karcz, S.; Wan, L.; Coutu, C.; Gillott, C.; Hegedus, D.D. Identification of the Mamestra configurata (Lepidoptera: Noctuidae) peritrophic matrix proteins and enzymes involved in peritrophic matrix chitin metabolism. Insect Sci. 2016, 23, 656–674. [Google Scholar] [CrossRef] [PubMed]
- Arakane, Y.; Muthukrishnan, S.; Kramer, K.J.; Specht, C.A.; Tomoyasu, Y.; Lorenzen, M.D.; Kanost, M.; Beeman, R.W. The Tribolium chitin synthase genes TcCHS1 and TcCHS2 are specialized for synthesis of epidermal cuticle and midgut peritrophic matrix. Insect Mol. Biol. 2005, 14, 453–463. [Google Scholar] [CrossRef]
- Moussian, B.; Schwarz, H.; Bartoszewski, S.; Nüsslein-Volhard, C. Involvement of chitin in exoskeleton morphogenesis in Drosophila melanogaster. J. Morphol. 2005, 264, 117–130. [Google Scholar] [CrossRef]
- Wang, Y.; Fan, H.; Huang, H.; Xue, J.; Wu, W.; Bao, Y.; Xu, H.; Zhu, Z.; Cheng, J.; Zhang, C. Chitin synthase 1 gene and its two alternative splicing variants from two sap-sucking insects, Nilaparvata lugens and Laodelphax striatellus (Hemiptera: Delphacidae). Insect Biochem. Mol. Biol. 2012, 42, 637–646. [Google Scholar] [CrossRef]
- Miller, S.; Shippy, T.D.; Tamayo, B.; Hosmani, P.S.; Flores-gonzalez, M.; Mueller, L.A.; Hunter, W.B.; Brown, S.J.; Elia, T.D.; Surya, S. Annotation of chitin biosynthesis genes in Diaphorina citri, the Asian citrus psyllid. Gigabyte 2021, 2, 1–12. [Google Scholar] [CrossRef]
- Grosscurt, A.C.; Jongsma, B. Mode of Action and Insecticidal Properties of Diflubenzuron; Springer: Dordrecht, The Netherlands, 1987; pp. 75–99. [Google Scholar]
- Cohen, E. Chitin synthesis and inhibition: A revisit. Pest Manag. Sci. 2001, 57, 946–950. [Google Scholar] [CrossRef]
- Merzendorfer, H. Chitin synthesis inhibitors: Old molecules and new developments. Insect Sci. 2013, 20, 121–138. [Google Scholar] [CrossRef]
- Douris, V.; Steinbach, D.; Panteleri, R.; Livadaras, I.; Pickett, J.A.; Van Leeuwen, T.; Nauen, R.; Vontas, J. Resistance mutation conserved between insects and mites unravels the benzoylurea insecticide mode of action on chitin biosynthesis. Proc. Natl. Acad. Sci. USA 2016, 113, 14692–14697. [Google Scholar] [CrossRef]
- Rösner, J.; Merzendorfer, H. Transcriptional plasticity of different ABC transporter genes from Tribolium castaneum contributes to diflubenzuron resistance. Insect Biochem. Mol. Biol. 2020, 116, 103282. [Google Scholar] [CrossRef]
- Tiwari, S.; Clayson, P.J.; Kuhns, E.H.; Stelinski, L.L. Effects of buprofezin and diflubenzuron on various developmental stages of Asian citrus psyllid, Diaphorina citri. Pest Manag. Sci. 2012, 68, 1405–1412. [Google Scholar] [CrossRef]
- Huvenne, H.; Smagghe, G. Mechanisms of dsRNA uptake in insects and potential of RNAi for pest control: A review. J. Insect Physiol. 2010, 56, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Dias, N.P.; Cagliari, D.; Dos Santos, E.A.; Smagghe, G.; Jurat-Fuentes, J.L.; Mishra, S.; Nava, D.E.; Zotti, M.J. Insecticidal Gene Silencing by RNAi in the Neotropical Region. Neotrop. Entomol. 2020, 49, 1–11. [Google Scholar] [CrossRef]
- Tijsterman, M.; Plasterk, R.H.A. Dicers at RISC: The mechanism of RNAi. Cell 2004, 117, 1–3. [Google Scholar] [CrossRef]
- Kishk, A.; Anber, H.A.I.; AbdEl Raof, T.K.; El Sherbeni, A.H.D.; Hamed, S.; Gowda, S.; Killiny, N. RNA interference of carboxyesterases causes nymph mortality in the Asian citrus psyllid, Diaphorina citri. Arch. Insect Biochem. 2017, 94, e21377. [Google Scholar] [CrossRef]
- Andrade, E.C.; Hunter, W.B. RNAi feeding bioassay: Development of a non-transgenic approach to control Asian citrus psyllid and other hemipterans. Entomol. Exp. Appl. 2017, 162, 389–396. [Google Scholar] [CrossRef]
- Yu, X.; Gowda, S.; Killiny, N. Double-stranded RNA delivery through soaking mediates silencing of the muscle protein 20 and increases mortality to the Asian citrus psyllid, Diaphorina citri. Pest Manag. Sci. 2017, 73, 1846–1853. [Google Scholar] [CrossRef] [PubMed]
- Chen, J.; Peng, Y.; Zhang, H.; Wang, K.; Tang, Y.; Gao, J.; Zhao, C.; Zhu, G.; Palli, S.R.; Han, Z. Transcript level is a key factor affecting RNAi efficiency. Pestic. Biochem. Phys. 2021, 176, 104872. [Google Scholar] [CrossRef]
- Galdeano, D.M.; Breton, M.C.; Lopes, J.R.S.; Falk, B.W.; Machado, M.A. Oral delivery of double-stranded RNAs induces mortality in nymphs and adults of the Asian citrus psyllid, Diaphorina citri. PLoS ONE 2017, 12, e171847. [Google Scholar] [CrossRef] [PubMed]
- Lu, Z.; Huang, Y.; Yu, H.; Li, N.; Xie, Y.; Zhang, Q.; Zeng, X.; Hu, H.; Huang, A.; Yi, L.; et al. Silencing of the Chitin Synthase Gene Is Lethal to the Asian Citrus Psyllid, Diaphorina citri. Int. J. Mol. Sci. 2019, 20, 3734. [Google Scholar] [CrossRef] [PubMed]
- Xia, W.; Ding, T.; Niu, J.; Liao, C.; Zhong, R.; Yang, W.; Liu, B.; Dou, W.; Wang, J. Exposure to Diflubenzuron Results in an Up-Regulation of a Chitin Synthase 1 Gene in Citrus Red Mite, Panonychus citri (Acari: Tetranychidae). Int. J. Mol. Sci. 2014, 15, 3711–3728. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of Relative Gene Expression Data Using Real-Time Quantitative PCR and the 2−ΔΔCT Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular Evolutionary Genetics Analysis Version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef]
- Shang, F.; Xiong, Y.; Xia, W.K.; Wei, D.D.; Wei, D.; Wang, J.J. Identification, characterization and functional analysis of a chitin synthase gene in the brown citrus aphid, Toxoptera citricida (Hemiptera, Aphididae). Insect Mol. Biol. 2016, 25, 422–430. [Google Scholar] [CrossRef]
- Merzendorfer, H. The cellular basis of chitin synthesis in fungi and insects: Common principles and differences. Eur. J. Cell Biol. 2011, 90, 759–769. [Google Scholar] [CrossRef] [PubMed]
- Hegedus, D.; Erlandson, M.; Gillott, C.; Toprak, U. New insights into peritrophic matrix synthesis, architecture, and function. Annu. Rev. Entomol. 2009, 54, 285–302. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.; Yang, H.; Zhou, C.; Yang, W.; Jin, D.; Long, G. Molecular cloning, expression, and functional analysis of the chitin synthase 1 gene and its two alternative splicing variants in the white-backed planthopper, Sogatella furcifera (Hemiptera: Delphacidae). Sci. Rep. 2019, 9, 1087. [Google Scholar] [CrossRef]
- Fotakis, E.A.; Mastrantonio, V.; Grigoraki, L.; Porretta, D.; Puggioli, A.; Chaskopoulou, A.; Osório, H.; Weill, M.; Bellini, R.; Urbanelli, S.; et al. Identification and detection of a novel point mutation in the Chitin Synthase gene of Culex pipiens associated with diflubenzuron resistance. PLoS Negl. Trop. Dis. 2020, 14, e8284. [Google Scholar] [CrossRef] [PubMed]
- Ye, C.; Jiang, Y.; An, X.; Yang, L.; Shang, F.; Niu, J.; Wang, J. Effects of RNAi-based silencing of chitin synthase gene on moulting and fecundity in pea aphids (Acyrthosiphon pisum). Sci. Rep. 2019, 9, 3694. [Google Scholar] [CrossRef]
- Zhang, J.; Liu, X.; Zhang, J.; Li, D.; Sun, Y.; Guo, Y.; Ma, E.; Zhu, K.Y. Silencing of two alternative splicing-derived mRNA variants of chitin synthase 1 gene by RNAi is lethal to the oriental migratory locust, Locusta migratoria manilensis (Meyen). Insect Biochem. Mol. Biol. 2010, 40, 824–833. [Google Scholar] [CrossRef] [PubMed]
- Boina, D.R.; Rogers, M.E.; Wang, N.; Stelinski, L.L. Effect of pyriproxyfen, a juvenile hormone mimic, on egg hatch, nymph development, adult emergence and reproduction of the Asian citrus psyllid, Diaphorina citri Kuwayama. Pest Manag. Sci. 2010, 66, 349–357. [Google Scholar] [CrossRef] [PubMed]
- Jiang, L.; Mu, L.; Jin, L.; Anjum, A.A.; Li, G. RNAi for chitin synthase 1 rather than 2 causes growth delay and molting defect in Henosepilachna vigintioctopunctata. Pestic. Biochem. Phys. 2021, 178, 104934. [Google Scholar] [CrossRef] [PubMed]
- Guedes, R.N.C.; Beins, K.; Navarro Costa, D.; Coelho, G.E.; Bezerra, H.S.D.S. Patterns of insecticide resistance in Aedes aegypti: Meta-analyses of surveys in Latin America and the Caribbean. Pest Manag. Sci. 2020, 76, 2144–2157. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Chen, J.; Fan, X.; Chen, W.; Zhang, W. MicroRNA and dsRNA targeting chitin synthase A reveal a great potential for pest management of the hemipteran insect Nilaparvata lugens. Pest Manag. Sci. 2017, 73, 1529–1537. [Google Scholar] [CrossRef] [PubMed]
- Mulder, R.; Gijswijt, M.J. The laboratory evaluation of two promising new insecticides which interfere with cuticle deposition. Pestic. Sci. 1973, 4, 737–745. [Google Scholar] [CrossRef]
- Zhang, J.; Zhu, K.Y. Characterization of a chitin synthase cDNA and its increased mRNA level associated with decreased chitin synthesis in Anopheles quadrimaculatus exposed to diflubenzuron. Insect Biochem. Mol. Biol. 2006, 36, 712–725. [Google Scholar] [CrossRef] [PubMed]
- Gong, C.; Wang, X.; Huang, Q.; Zhang, J.; Zhang, Y.; Zhan, X.; Zhang, S.; Hasnain, A.; Ruan, Y.; Shen, L. The fitness advantages of bistrifluron resistance related to chitin synthase A in Spodoptera litura (Fab.) (Noctuidae: Lepidoptera). Pest Manag. Sci. 2021, 77, 3458–3468. [Google Scholar] [CrossRef]
- Zhu, K.Y.; Palli, S.R. Mechanisms, Applications, and Challenges of Insect RNA Interference. Annu. Rev. Entomol. 2020, 65, 293–311. [Google Scholar] [CrossRef] [PubMed]
- Shao, Z.; Li, Y.; Ding, J.; Liu, Z.; Zhang, X.; Wang, J.; Sheng, S.; Wu, F. Identification, Characterization, and Functional Analysis of Chitin Synthase Genes in Glyphodes pyloalis Walker (Lepidoptera: Pyralidae). Int. J. Mol. Sci. 2020, 21, 4656. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experiments | Name | Sequences |
|---|---|---|
| PCR | DcCHSA5F | TWCTGCAATAAAGAYYTA |
| DcCHSA5R | AKATAWRTGAARTAWCGWGT | |
| DcCHSA3F | ATGATGGAVATGWTNARSTSKAT | |
| DcCHSA3R | CATAAGRAATATHGTKCCHGGDC | |
| DcCHSAZF | ACTACAACGAGCAAACATTCA | |
| DcCHSAZR | GAGAATGGGCGTAATGTTGT | |
| 3′-RACE | DcCHSAF1 | GACAAGCAAGCGGTGGACATA |
| DcCHSAF2 | AGGTCCGCGTCAACTCTTCTTCCATC | |
| 5′-RACE | DcCHSAR1 | AGGCGGGAGGTCTCGGAACA |
| DcCHSAR2 | ATGGAGGTGACGAAGATAAA | |
| 3′/5′-RACE | NUP | AAGCAGTGGTATCAACGCAGAGT |
| UPM (mix) | TAATACGACTCACTATAGGGCAAGCAGTGGTATCAACGCAGAGT | |
| CTAATACGACTCACTATAGGGC | ||
| RT-qPCR | DcActinF | CCATCTTGGCTTCTCTGTCTAC |
| DcActinR | CATTTGCGGTGAACGATTCC | |
| DcTubulinF | TTTCCAACACCACCGCTAT | |
| DcTubulinR | AGGTCTTCCCTCGCCTCTG | |
| DcCHSAqF | CCACGACTCCTTCTCACAG | |
| DcCHSAqR | TCATGGCGAAATTATCCTC | |
| RNAi | GFPiF | GCCAACACTTGTCACTACTT |
| GFPiR | GGAGTATTTTGTTGATAATGGTCTG | |
| GFPdsF | taatacgactcactatagggGCCAACACTTGTCACTACTT | |
| GFPdsR | taatacgactcactatagggGGAGTATTTTGTTGATAATGGTCTG | |
| DcCHSAiF | CGAGTGGTAGACCCGAACAT | |
| DcCHSAiR | ATGAAAAAGGCGAACACGAC | |
| DcCHSAdsF | taatacgactcactatagggCGAGTGGTAGACCCGAACAT | |
| DcCHSAdsR | taatacgactcactatagggATGAAAAAGGCGAACACGAC |
| Gene Name | GenBank Accession Number | Species |
|---|---|---|
| DcCHSA | this study | Diaphorina citri |
| SeCHSA | AAZ03545.1 | Spodoptera exigua |
| HaCHSA | AKJ54482.1 | Helicoverpa armigera |
| EoCHS1 | ACA50098.1 | Ectropis obliqua |
| MsCHSA | AAL38051.2 | Manduca sexta |
| PxCHS1 | API61827.1 | Plutella xylostella |
| DtCHS1 | XP_017009970.1 | Drosophila takahashii |
| CqCHSA | XP_001866798.1 | Culex quinquefasciatus |
| AqCHS1 | ABD74441.1 | Anopheles quadrimaculatus |
| NvCHS1 | XP_001602290 | Nasonia vitripennis |
| BtCHSA | XP_00339885 | Bombus terrestris |
| AmCHS1 | XP_395677 | Apis mellifera |
| AtCHS1 | AFM38193.1 | Anasa tristis |
| LhCHSA | JAQ09912.1 | Lygus hesperus |
| AcCHSA | KR611528.1 | Aphis citricidus |
| AgCHSA | AFJ00066.1 | Aphis glycines |
| NlCHSA | JQ040014 | Nilaparvata lugens |
| SfCHSA | KY987034 | Sogatella furcifera |
| LsCHSA | JQ040012 | Laodelphax striatellus |
| PcCHS1 | AJQ20794.1 | Panonychus citri |
| CmCHS2 | AJF93428.1 | Mamestra configurata |
| MysCHSB | ASF79498.1 | Mythimna separate |
| SeCHSB | ABI96087.1 | Spodoptera exigua |
| HaCHSB | AKZ08595.1 | Helicoverpa armigera |
| DmCHSB | CAC83725.1 | Drosophila melanogaster |
| Toxic Regression Equation | 95% Confidence Interval | Correlation Coefficient | Chi-Square Value | Half Lethal Concentration (mg/L) |
|---|---|---|---|---|
| y = −4.012 + 1.867x | 110.295–174.718 | 0.903 | 1.861 | 140.992 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, C.; Hu, W.; Yu, Z.; Liu, X.; Wang, J.; Xin, T.; Zou, Z.; Xia, B. Characterization of Chitin Synthase A cDNA from Diaphorina citri (Hemiptera: Liviidae) and Its Response to Diflubenzuron. Insects 2022, 13, 728. https://doi.org/10.3390/insects13080728
Zhang C, Hu W, Yu Z, Liu X, Wang J, Xin T, Zou Z, Xia B. Characterization of Chitin Synthase A cDNA from Diaphorina citri (Hemiptera: Liviidae) and Its Response to Diflubenzuron. Insects. 2022; 13(8):728. https://doi.org/10.3390/insects13080728
Chicago/Turabian StyleZhang, Cong, Wenfeng Hu, Zhuo Yu, Xian Liu, Jing Wang, Tianrong Xin, Zhiwen Zou, and Bin Xia. 2022. "Characterization of Chitin Synthase A cDNA from Diaphorina citri (Hemiptera: Liviidae) and Its Response to Diflubenzuron" Insects 13, no. 8: 728. https://doi.org/10.3390/insects13080728