
Mosquitocidal Activity of the Methanolic Extract of Annickia chlorantha and Its Isolated Compounds against Culex pipiens, and Their Impact on the Non-Target Organism Zebrafish, Danio rerio

 , , , , , , and
, , , , , , and 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Sample
2.2. Mosquito Colony
2.3. Larvicidal Activity
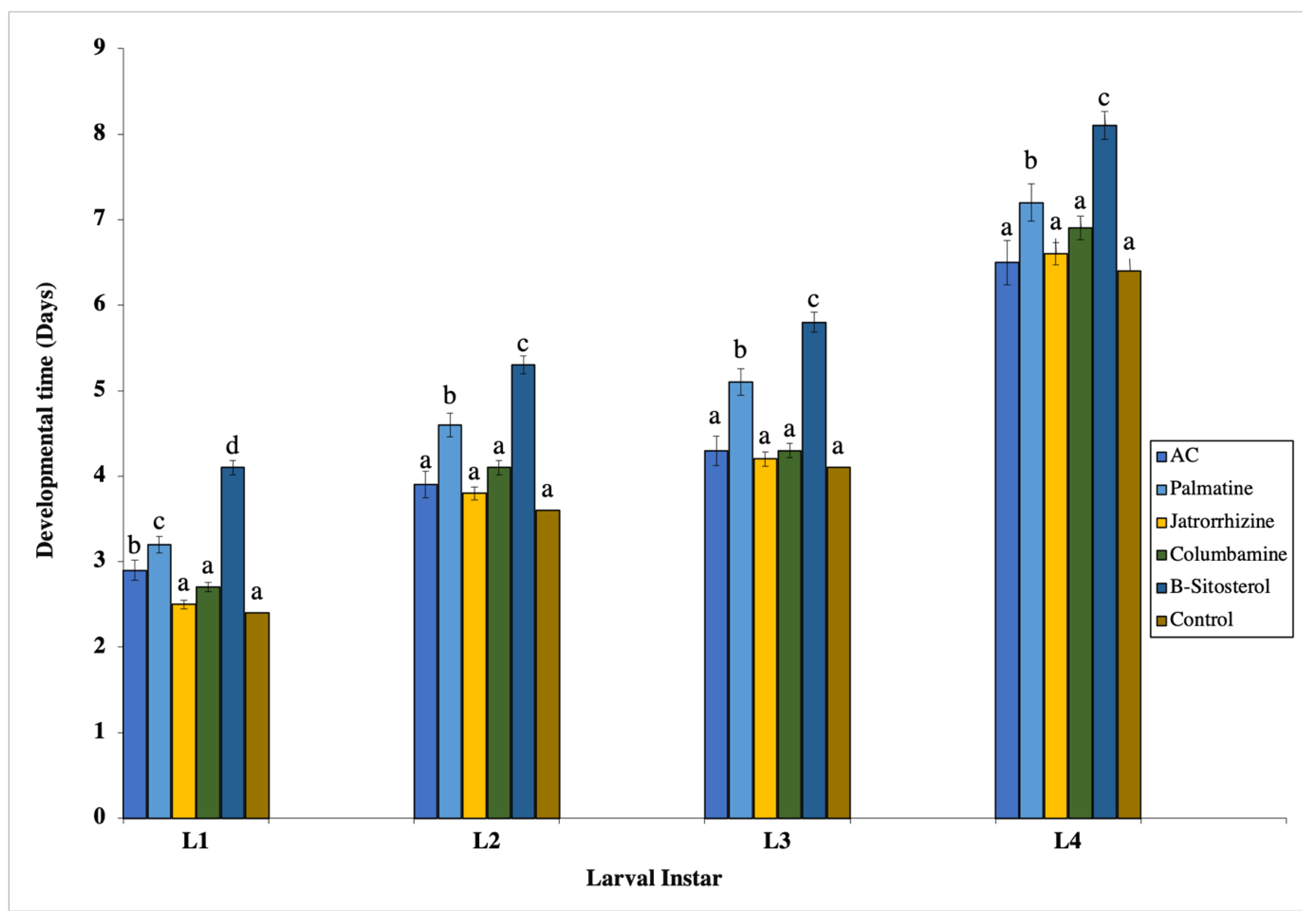
2.4. Developmental Durations
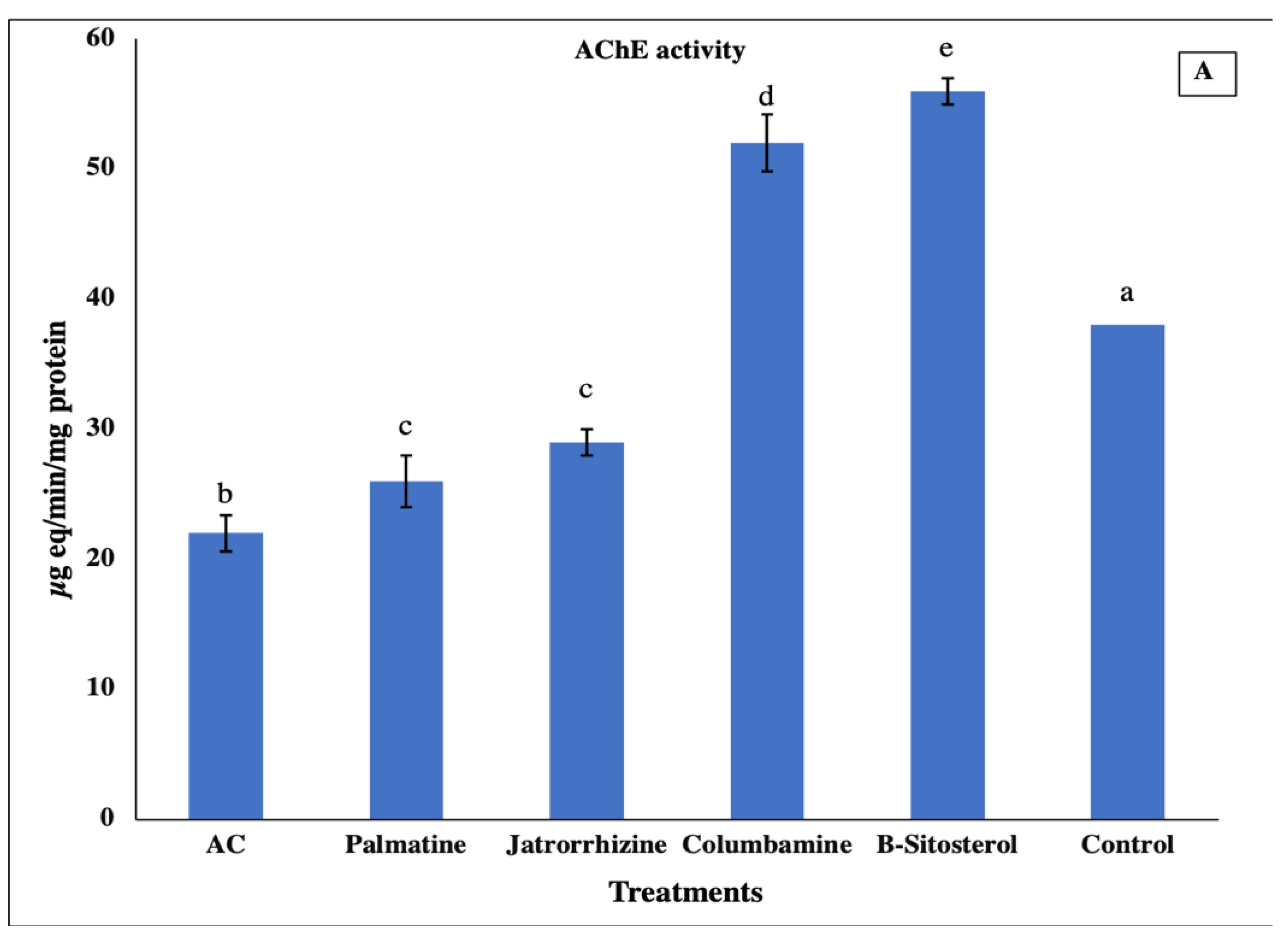
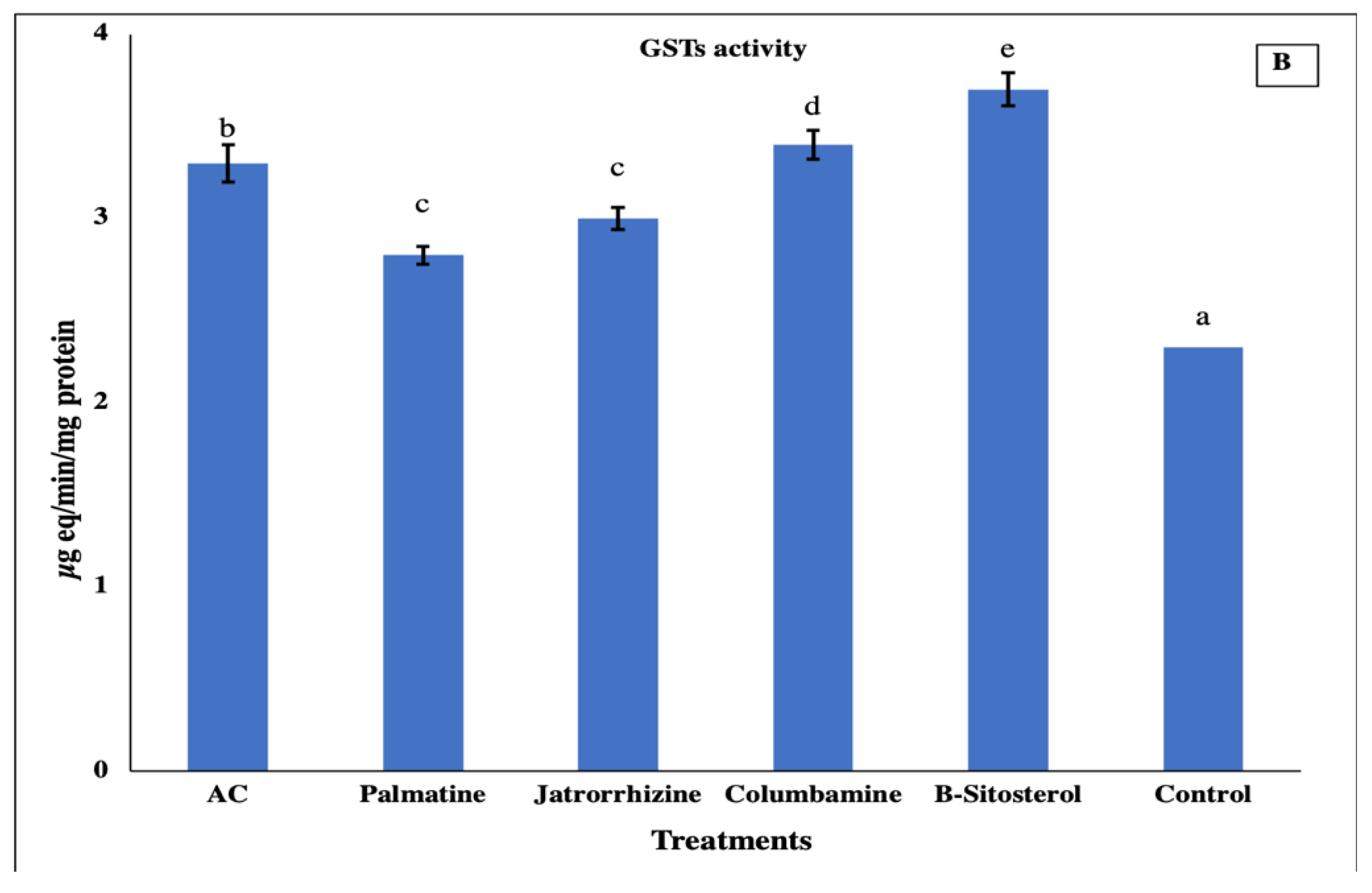
2.5. Biochemical Assay
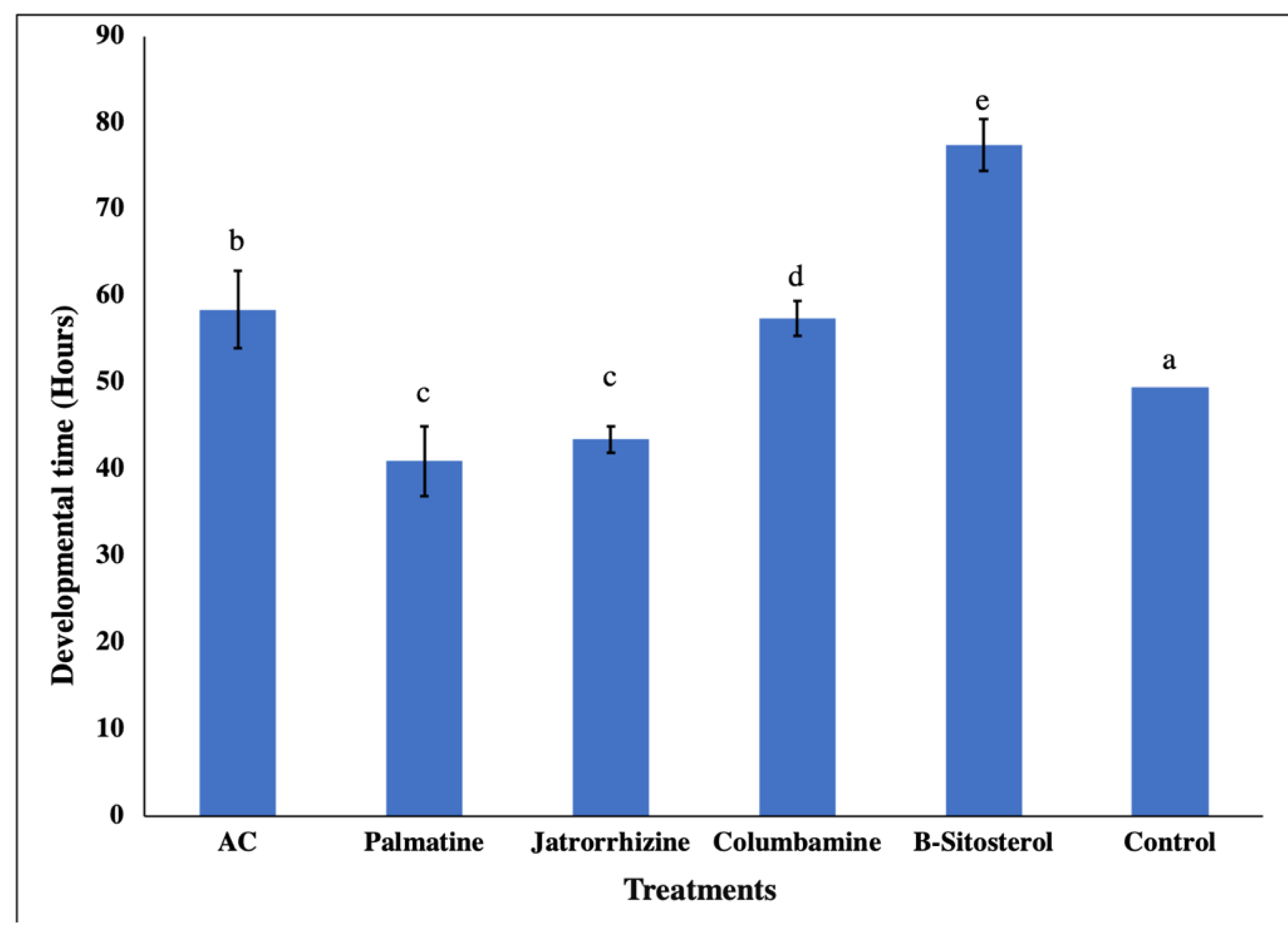
2.6. Adult Emergence and Sex Ratio
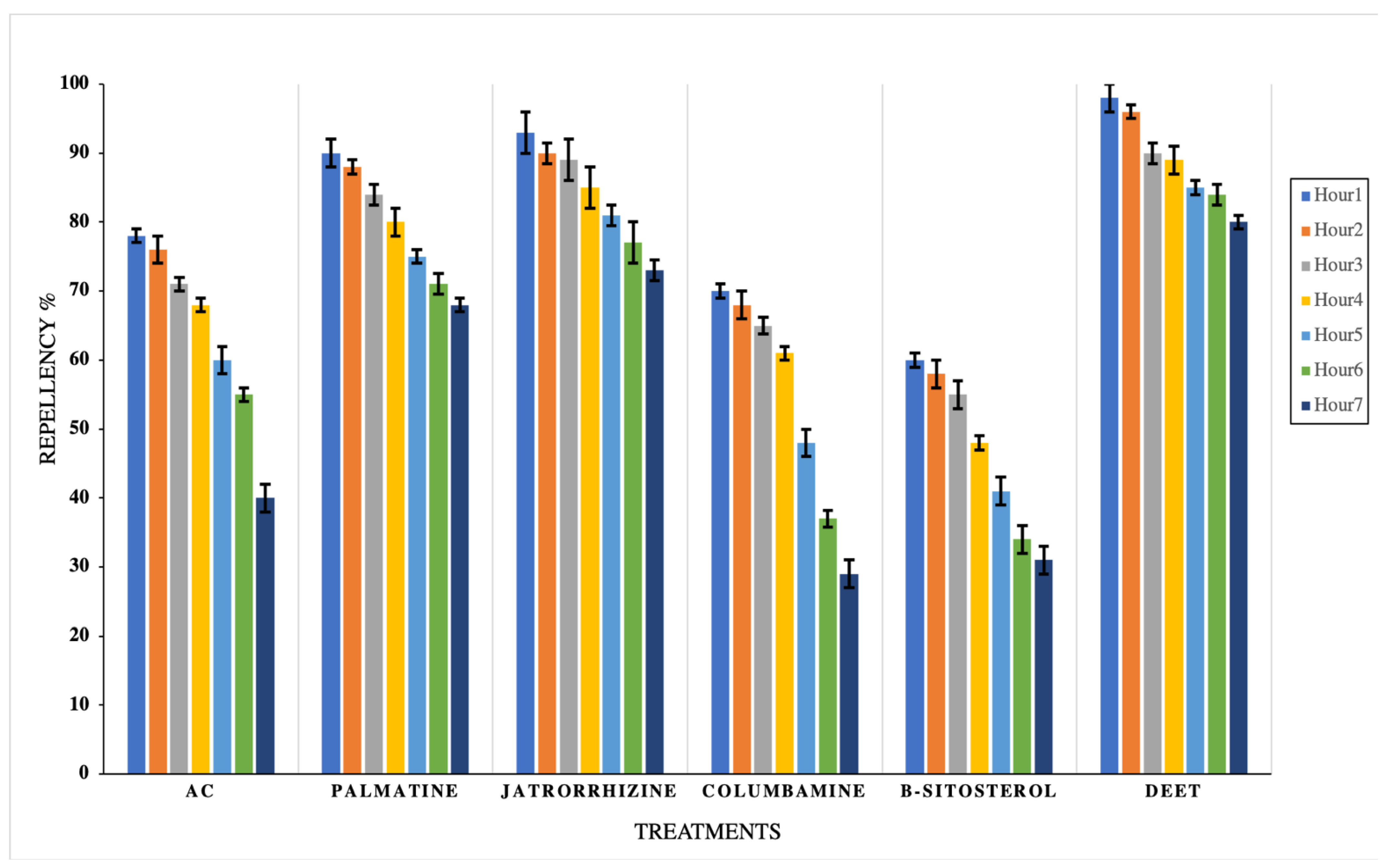
2.7. Repellency Test
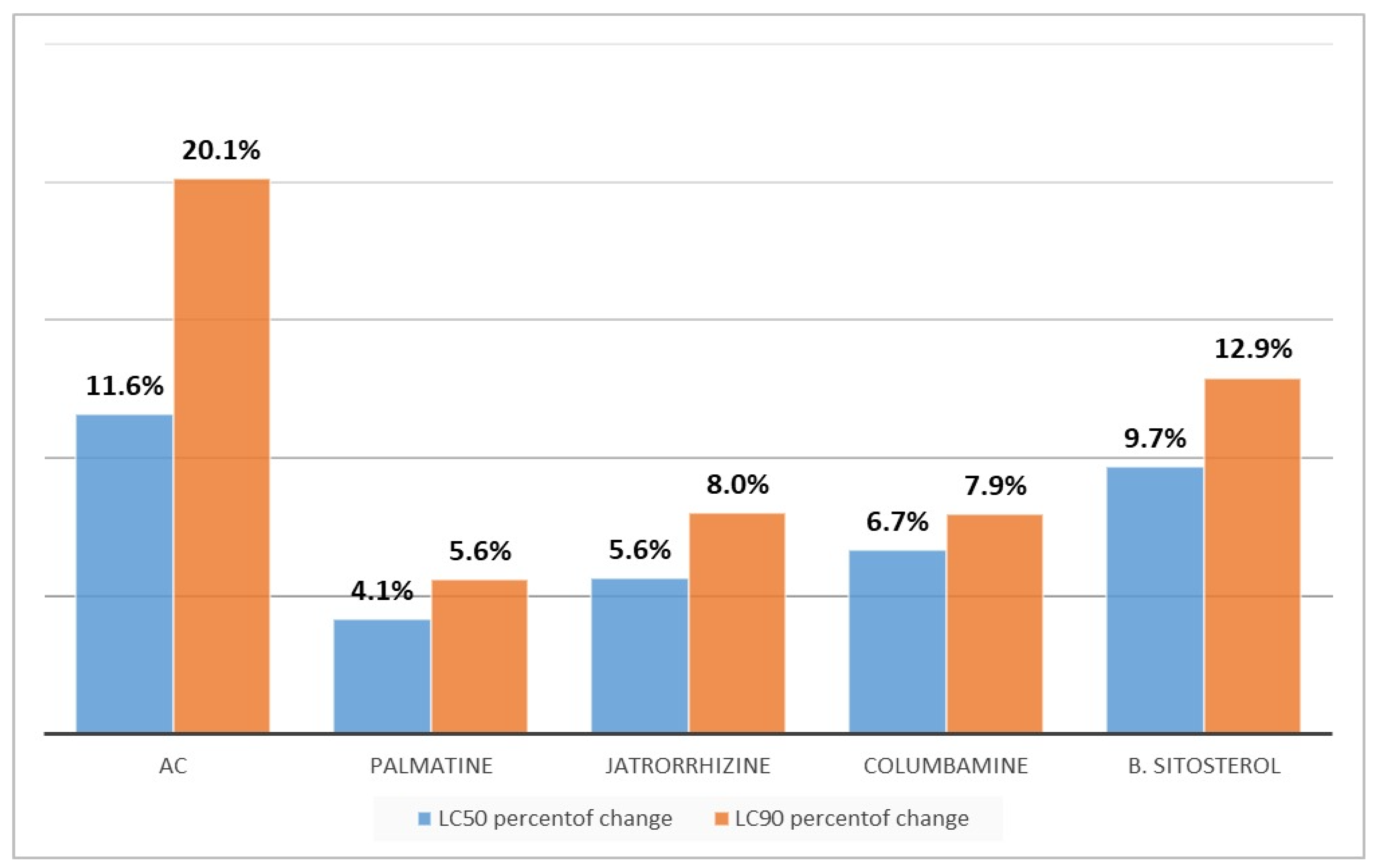
2.8. Toxicity to the Non-Target Organism
2.9. Statistical Analysis
2.10. Molecular Modeling
2.10.1. Preparation of Small Molecule
2.10.2. Selection of Proteins Structures
2.10.3. Binding Site Analysis
2.10.4. MOE Stepwise Docking Method
2.10.5. Computational Methods
3. Results
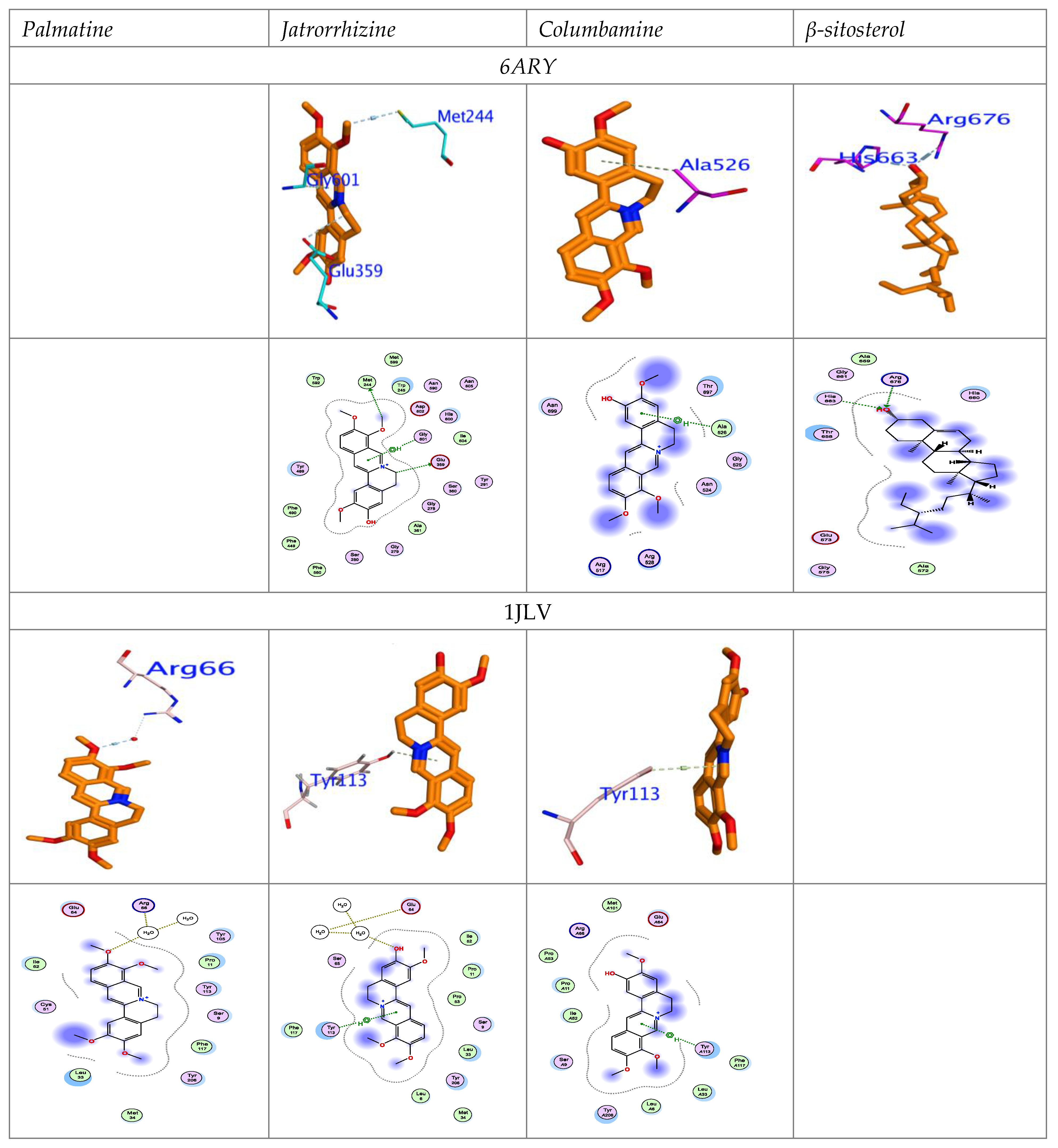
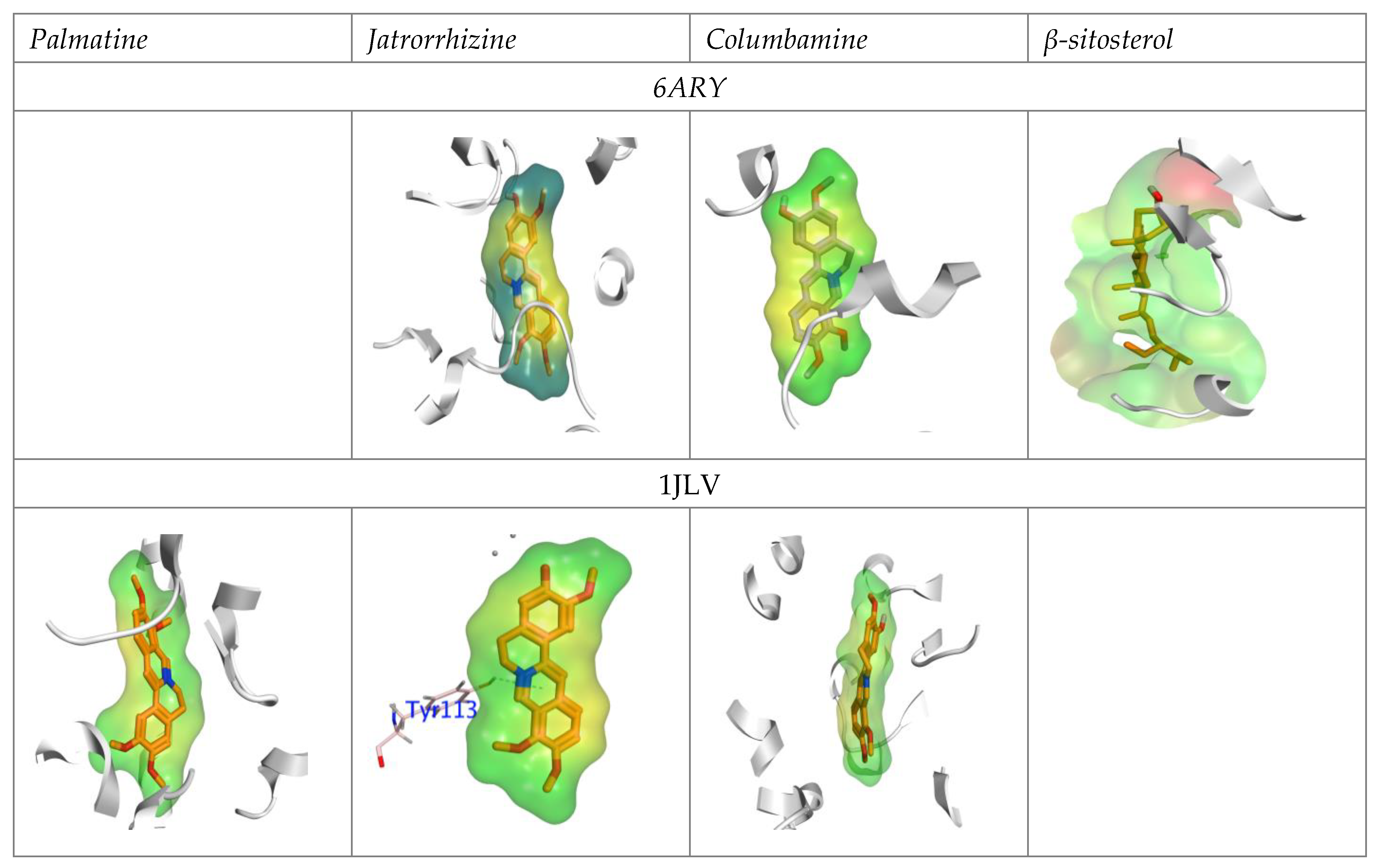
Binding Efficacy for Molecular Docking
4. Discussion
4.1. Chemical Interaction with AChE and GST Domain Based on Molecular Docking Studies
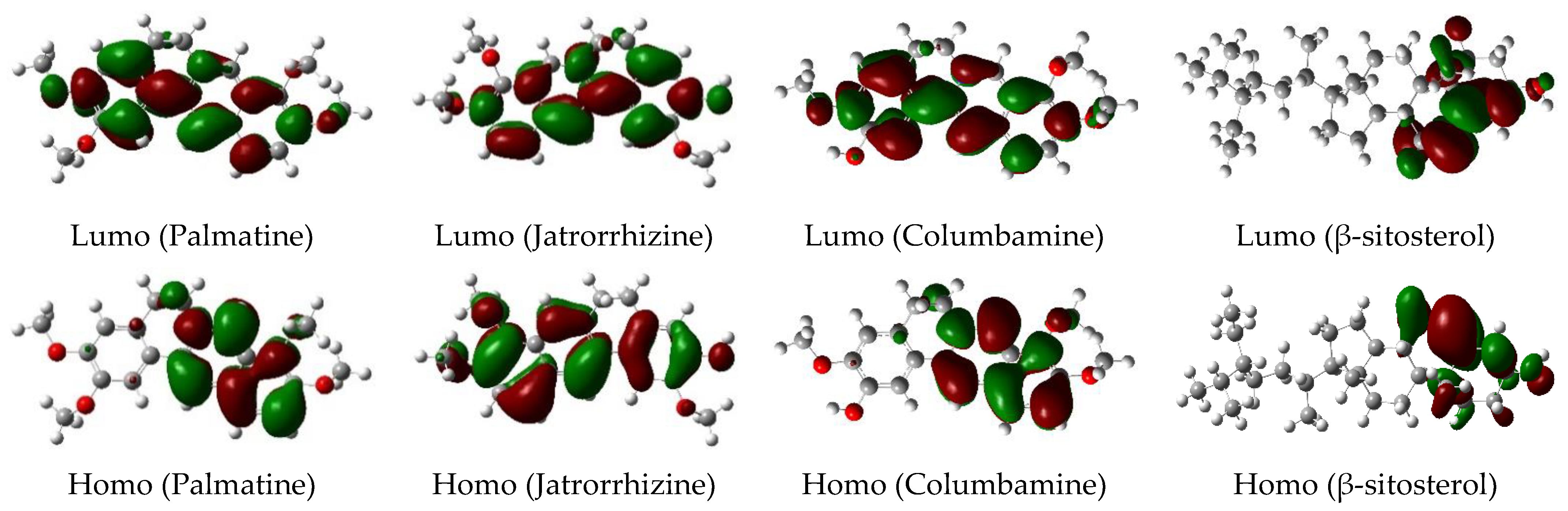
4.2. DFT Calculation
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Hasaballah, A.I.; El-Naggar, H.A. Antimicrobial activities of some marine sponges, and its biological, repellent effects against Culex pipiens (Diptera: Culicidae). Annu. Res. Rev. Biol. 2017, 12, 1–14. [Google Scholar] [CrossRef]
- Gabarty, A.; Selim, T.A.; Hasaballah, A.I. Effect of gamma irradiation on protease and nuclease enzymes activity and egg oviposition of Culex pipiens mosquito engorged with Hepatitis C Virus (HCV). J. Radiat. Res. Appl. Sci. 2022, 15, 1–6. [Google Scholar] [CrossRef]
- Hasaballah, A.I.; El-Naggar, H.A.; Abdelbary, S.; Bashar, M.A.E.; Selim, T.A. Eco-friendly Synthesis of Zinc Oxide Nanoparticles by Marine Sponge, Spongia officinalis: Antimicrobial and Insecticidal Activities Against the Mosquito Vectors, Culex pipiens and Anopheles pharoensis. BioNanoScience 2021, 12, 89–104. [Google Scholar] [CrossRef]
- Hasaballah, A.I. Toxicity of some plant extracts against vector of lymphatic filariasis. Culex Pipiens. J. Egypt. Soc. Parasitol. 2015, 45, 185–193. [Google Scholar]
- Hasaballah, A.; Selim, T.; Tanani, M.; Nasr, E. Lethality and Vitality Efficiency of Different Extracts of Salix safsaf Leaves against the House Fly, Musca domestica L. (Diptera: Muscidae). Afr. Entomol. 2021, 29, 479–490. [Google Scholar] [CrossRef]
- Shehabeldine, A.M.; Elbahnasawy, M.A.; Hasaballah, A.I. Green Phytosynthesis of Silver Nanoparticles Using Echinochloa stagnina Extract with Reference to their Antibacterial, Cytotoxic, and Larvicidal Activities. BioNanoScience 2021, 11, 526–538. [Google Scholar] [CrossRef]
- Benelli, G. Research in mosquito control: Current challenges for a brighter future. Parasitol. Res. 2015, 114, 2801–2805. [Google Scholar] [CrossRef] [PubMed]
- Pavela, R.; Maggi, F.; Lupidi, G.; Mbuntcha, H.; Woguem, V.; Womeni, H.M.; Barboni, L.; Tapondjou, L.A.; Benelli, G. Clausena anisata and Dysphania ambrosioides essential oils: From ethno-medicine to modern uses as effective insecticides. Environ. Sci. Pollut. Res. 2017, 25, 10493–10503. [Google Scholar] [CrossRef]
- Malik, A.; Singh, N.; Satya, S. House fly (Musca domestica): A review of control strategies for a challenging pest. J. Environ. Sci. Health Part B 2007, 42, 453–469. [Google Scholar] [CrossRef] [PubMed]
- Isman, M.B. Botanical insecticides, deterrents and repellents in modern agriculture and an in increasingly regulated world. Annu. Rev. Entomol. 2006, 51, 45–56. [Google Scholar] [CrossRef]
- Zibaee, A.; Bandani, A.R.; Tork, M. Effect of the entomopathogenic fungus, Beauveria bassiana, and its secondary metabolite on detoxifying enzyme activities and acetylcholinesterase (AChE) of the Sunn pest, Eurygaster integriceps (Heteroptera: Scutellaridae). Biocontrol Sci. Technol. 2009, 19, 485–498. [Google Scholar] [CrossRef]
- Enayati, A.A.; Ranson, H.; Hemingway, J. Insect glutathione transferases and insecticide resistance. Insect Mol. Biol. 2005, 14, 3–8. [Google Scholar] [CrossRef] [PubMed]
- Adesokan, A.; Yakubu, M.; Owoyele, B.; Akanji, M.; Soladoye, A.; Lawal, O. Effect of administration of aqueous and ethanolic extracts of Enantia chlorantha stem bark on brewer’s yeast-induced pyresis in rats. Afr. J. Biochem. Res. 2008, 2, 165–169. [Google Scholar]
- Vennerstrom, J.L.; Klayman, D.L. Protoberberine alkaloids as antimalarials. J. Med. Chem. 1988, 31, 1084–1087. [Google Scholar] [CrossRef] [PubMed]
- Moody, J.; Ogundipe, O.; Akang, E.; Agbedana, E. Toxicological studies on the purified protoberberine alkaloidal fraction of Enantia chlorantha Oliv (Annonaceae). Afr. J. Med. Med. Sci. 2007, 36, 317–323. [Google Scholar]
- Nyasse, B.; Nkwengoua, E.; Sondengam, B.; Denier, C.; Willson, M. Modified berberine and protoberberines from Enantia chlorantha as potential inhibitors of Trypanosoma brucei. Die Pharmazie 2002, 57, 358–361. [Google Scholar] [CrossRef]
- Imieje, V.; Falodun, A.; Zaki, A.; Ali, Z.; Khan, I.; Tekwani, B.; Khan, S.; Egiebor, N. Antiprotozoal and Cytotoxicity Studies of Fractions and Compounds from Enantia chlorantha. Trop. J. Nat. Prod. Res. 2017, 1, 89–94. [Google Scholar] [CrossRef]
- Abdallah, F.I.; Rady, M.H.; Merdan, B.A.; Shaarawi, F.A.; Mohammed, A.F.; Alshammery, K.A.; Dahab, A.A. Effects of blood sources and artificial blood feeding membranes on the biological parameters and hepatitis C virus infectivity of Culex pipiens (Diptera: Culicidae). Afr. Entomol. 2021, 29, 262–273. [Google Scholar] [CrossRef]
- World Health Organization. Guidelines for Laboratory and Field Testing of Mosquito Larvicides; World Health Organization: Geneva, Switzerland, 2005; pp. 1–39. [Google Scholar]
- Kauffman, E.; Payne, A.; Franke, M.A.; Schmid, M.A.; Harris, E.; Kramer, L.D. Rearing of Culex spp. and Aedes spp. mosquitoes. Bio-Protocol 2017, 7, e2542. [Google Scholar] [CrossRef] [PubMed]
- Hasaballah, A.I. Impact of gamma irradiation on the development and reproduction of Culex pipiens (Diptera; Culicidae). Int. J. Radiat. Biol. 2018, 94, 844–849. [Google Scholar] [CrossRef]
- Simpson, D.R.; Bulland, D.L.; Linquist, D.A. A semimicro-technique for estimation of cholinesterase activity in boll weevils. Ann. Entomol. Soc. Am. 1964, 57, 367–371. [Google Scholar] [CrossRef]
- Kao, C.-H.; Hung, C.-F.; Sun, C.-N. Parathion and Methyl Parathion Resistance in Diamondback Moth (Lepidoptera: Plutellidae) Larvae. J. Econ. Entomol. 1989, 82, 1299–1304. [Google Scholar] [CrossRef]
- Khazanie, R. Elementary Statistics in a World of Applications; Good Year Publishing Co.: Santa Monica, CA, USA, 1979. [Google Scholar]
- Shetty, V.; Shetty, N.; Harini, B.; Ananthanarayana, S.; Jha, S.; Chaubey, R. Effect of gamma radiation on life history traits of Aedes aegypti (L.). Parasite Epidemiol. Control 2016, 1, 26–35. [Google Scholar] [CrossRef]
- Abbott, W.S. A Method of Computing the Effectiveness of an Insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Mount, D.I. Methods for Aquatic Toxicity Identification Evaluations: Phase I: Toxicity Characterization Procedures; US Environmental Protection Agency, Environmental Research Laboratory, National Effluent Toxicity Assessment Center: Washington, DC, USA, 1988; Volume 88.
- Deo, P.G.; Hasan, S.B.; Majumdar, S.K. Toxicity and suitability of some insecticides for household use. Int. Pest Control 1988, 30, 118–129. [Google Scholar]
- Molecular Operating Environment (MOE). Chemical Computing Group ULC, 1010 Sherbooke St. West, Suite #910; Molecular Operating Environment (MOE): Montreal, QC, Canada, 2019. [Google Scholar]
- Cheung, J.; Mahmood, A.; Kalathur, R.; Liu, L.; Carlier, P.R. Structure of the G119S Mutant Acetylcholinesterase of the Malaria Vector Anopheles gambiae Reveals Basis of Insecticide Resistance. Structure 2017, 26, 130–136.e2. [Google Scholar] [CrossRef]
- Oakley, A.J.; Harnnoi, T.; Udomsinprasert, R.; Jirajaroenrat, K.; Ketterman, A.J.; Wilce, M.C. The crystal structures of glutathione S-transferases isozymes 1-3 and 1-4 from Anopheles dirus species B. Protein Sci. 2009, 10, 2176–2185. [Google Scholar] [CrossRef]
- Elumalai, K.; Dhanasekaran, S.; Krishnappa, K. Larvicidal activity of saponin isolated from Gymnema sylvestre R. Br. (Asclepiadaceae) against Japanese encephalitis vector, Culex tritaeniorhynchus Giles (Diptera: Culicidae). Eur. Rev. Med. Pharmacol. Sci. 2013, 17, 1404–1410. [Google Scholar]
- Al-Khazraji, A.; Mustafa, M.A. Effect of some plant extracts on the Culex pipiens molestus Forskal larvae. Iraqi J. Veter Sci. 2008, 22, 9–12. [Google Scholar] [CrossRef]
- Abutaha, N.; Al-Mekhlafi, F.A.; Al-Keridis, L.A.; Farooq, M.; Nasr, F.A.; Al-Wadaan, M. Larvicidal potency of selected xerophytic plant extracts onCulex pipiens (Diptera: Culicidae). Entomol. Res. 2018, 48, 362–371. [Google Scholar] [CrossRef]
- Talukder, F.A. Plant products as potential stored-product insect stored management agents—A mini review. Emir. J. Food Agric. 2006, 18, 17–32. [Google Scholar] [CrossRef]
- Koul, O.; Waliai, S.; Dhaliwal, G.S. Essential oils as green pesticides: Potential and constraints. Biopestic. Int. 2008, 4, 63–84. [Google Scholar]
- Shaalan, E.A.S.; Canyon, D.; Younes, M.W.F.; Abdel-Wahab, H.; Mansour, A.H. A review of botanical phytochemicals with mosquitocidal potential. Environ. Int. 2005, 31, 1149–1166. [Google Scholar] [CrossRef]
- Al-Solami, H.M. Larvicidal activity of plant extracts by inhibition of detoxification enzymes in Culex pipiens. J. King Saud Univ. Sci. 2021, 33, 101371. [Google Scholar] [CrossRef]
- Elimam, A.M.; Elmalik, K.H.; Ali, F.S. Efficacy of leaves extract of Calotropis procera Ait. (Asclepiadaceae) in controlling Anopheles arabiensis and Culex quinquefasciatus mosquitoes. Saudi J. Biol. Sci. 2009, 16, 95–100. [Google Scholar] [CrossRef]
- Singh, R.K.; Dhiman, R.C.; Mittal, P.K. Studies on mosquito larvicidal properties of Eucalyptus citriodora Hook (family-Myrtaceae). J. Commun. Dis. 2007, 39, 233–236. [Google Scholar]
- Bara, J.J.; Montgomery, A.; Muturi, E.J. Sublethal effects of atrazine and glyphosate on life history traits of Aedes aegypti and Aedes albopictus (Diptera: Culicidae). Parasitol. Res. 2014, 113, 2879–2886. [Google Scholar] [CrossRef]
- Steinwascher, K. Competition among Aedes aegypti larvae. PLoS ONE 2018, 13, e0202455. [Google Scholar] [CrossRef]
- Das, N.G.; Nath, D.R.; Baruah, I.; Talukdar, P.K.; Das, S.C. Field evaluation of herbal mosquito repellents. J. Commun. Dis. 1999, 31, 241–245. [Google Scholar]
- Rajkumar, S.; Jebanesan, A. Oviposition deterrent and skin repellent activities of Solanum trilobatum leaf extract against the malarial vector Anopheles stephensi. J. Insect Sci. 2005, 5, 15. [Google Scholar]
- Mullai, K.; Jebanesan, A.; Pushpanathan, T. Effect of bioactive fractions of Citrullus vulgaris Schrad. leaf extract against Anopheles stephensi and Aedes aegypti. Parasitol. Res. 2008, 102, 951–955. [Google Scholar] [CrossRef]
- Junkum, A.; Maleewong, W.; Saeung, A.; Champakaew, D.; Chansang, A.; Amornlerdpison, D.; Pitasawat, B. Ligusticum sinense nanoemulsion gel as potential repellent against Aedes aegypti, Anopheles minimus, and Culex quinquefasciatus (Diptera: Culicidae). Insects 2021, 12, 596. [Google Scholar] [CrossRef] [PubMed]
- Foldbjerg, R.; Jiang, X.; Miclăus, T.; Chen, C.; Autrup, H.; Beer, C. Silver nanoparticles—Wolves in sheep’s clothing? Toxicol. Res. 2015, 4, 563–575. [Google Scholar] [CrossRef]
- Patil, C.D.; Borase, H.P.; Patil, S.V.; Salunkhe, R.B.; Salunke, B.K. Larvicidal activity of silver nanoparticles synthesized using Pergularia daemia plant latex against Aedes aegypti and Anopheles stephensi and nontarget fish Poecillia reticulata. Parasitol. Res. 2012, 111, 555–562. [Google Scholar] [CrossRef]
- Haldar, K.M.; Haldar, B.; Chandra, G. Fabrication, characterization and mosquito larvicidal bioassay of silver nanoparticles synthesized from aqueous fruit extract of putranjiva, Drypetes roxburghii (Wall.). Parasitol. Res. 2013, 112, 1451–1459. [Google Scholar] [CrossRef]
- Benelli, G. Plant-mediated biosynthesis of nanoparticles as an emerging tool against mosquitoes of medical and veterinary importance: A review. Parasitol. Res. 2016, 115, 23–34. [Google Scholar] [CrossRef]
- Sussman, J.L.; Harel, M.; Frolow, F.; Oefner, C.; Goldman, A.; Toker, L.; Silman, I. Atomic structure of acetylcholinesterase from Torpedo californica: A prototypic acetylcholine-binding protein. Science 1991, 253, 872–879. [Google Scholar] [CrossRef] [PubMed]
- Kryger, G.; Silman, I.; Sussman, J.L. Structure of acetylcholinesterase complexed with E2020 (Aricept®): Implications for the design of new anti-Alzheimer drugs. Structure 1999, 7, 297–307. [Google Scholar] [CrossRef]
- Ma, Y.-L.; Zhou, R.-J.; Zeng, X.-Y.; An, Y.-X.; Qiu, S.-S.; Nie, L.-J. Synthesis, DFT and antimicrobial activity assays in vitro for novel cis/trans-but-2-enedioic acid esters. J. Mol. Struct. 2014, 1063, 226–234. [Google Scholar] [CrossRef]
- Al-Amiery, A.A.; Al-Bayati, R.I.; Saed, F.M.; Ali, W.B.; Kadhum, A.A.H.; Mohamad, A.B. Novel Pyranopyrazoles: Synthesis and Theoretical Studies. Molecules 2012, 17, 10377–10389. [Google Scholar] [CrossRef]
- Mansour, A.M. Coordination behavior of sulfamethazine drug towards Ru (III) and Pt (II) ions: Synthesis, spectral, DFT, magnetic, electrochemical and biological activity studies. Inorg. Chim. Acta 2013, 394, 436–445. [Google Scholar] [CrossRef]
- Wang, Q.; Wang, H.; Wei, L.; Yang, S.-W.; Chen, Y. Reactive Sites for Chiral Selective Growth of Single-Walled Carbon Nanotubes: A DFT Study of Ni55–Cn Complexes. J. Phys. Chem. A 2012, 116, 11709–11717. [Google Scholar] [CrossRef] [PubMed]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds (μg/mL) | A. chlorantha (AC) | Palmatine | Jatrorrhizine | Columbamine | β-Sitosterol |
|---|---|---|---|---|---|
| LC50 (LCL–UCL) (μg/mL) | 162.630 (144.472–182.347) | 33.392 (27.366–40.343) | 91.343 (74.861–110.929) | 61.440 (53.260–70.980) | 123.236 (52.365–210.466) |
| LC90 (LCL–UCL) (μg/mL) | 433.95 (365.123–546.16) | 81.522 (63.405–122.239) | 228.135 (174.962–354.875) | 119.542 (97.470–171.995) | 254.709 (166.189–4711.963) |
| Regression equation | Y = 0.2499X − 1.0163 | Y = 1.3293X − 5.1626 | Y = 0.5161X − 6.7236 | Y = 1.1267X − 23.867 | Y = 0.456X − 14.267 |
| Statistic summary | d. f. = 5, p < 0.001 | d. f. = 5, p < 0.001 | d. f. = 5, p < 0.001 | d. f. = 5, p < 0.001 | d. f. = 5, p < 0.001 |
| χ2 | 19.459 | 7.121 | 5.140 | 6.860 | 8.006 |
| Treatments | Adult Emergence (%) | Sex Ratio (%) (Mean ± SE) | |||
|---|---|---|---|---|---|
| n | Mean ± SE | n | Males | Females | |
| Control | 75 | 100.0 ± 0.0 a | 75 | 46.67 ± 1.33 a | 53.33 ± 1.33 a |
| A. chlorantha (AC) | 38 | 80.13 ± 1.58 b | 31 | 40.72 ± 1.17 b | 59.28 ± 1.87 b |
| Palmatine | 35 | 68.68 ± 2.02 c | 24 | 45.83 ± 4.17 a | 54.17 ± 4.17 a |
| Jatrorrhizine | 35 | 72.34 ± 4.54 d | 25 | 43.52 ± 6.02 a | 56.48 ± 6.02 a |
| Columbamine | 36 | 83.08 ± 1.53 b | 30 | 46.97 ± 1.52 a | 53.03 ± 1.5 a |
| β-Sitosterol | 36 | 83.33 ± 4.81 b | 30 | 46.63 ± 1.71 a | 53.37 ± 1.71 a |
| Statistic summary | d. f. = 5 p < 0.05 | d. f. = 5 p > 0.05 | d. f. = 5 p > 0.05 | ||
| Compounds (μg/mL) | A. chlorantha | Palmatine | Jatrorrhizine | Columbamine | β-Sitosterol |
|---|---|---|---|---|---|
| LC50 ± SE (LCL–UCL) | 1406.6 ± 66.4 (984.7–4196.9) | 805.5 ± 20.1 (641.6–1153.4) | 1622.3 ± 108.8 (998.2–16177.5) | 922.3 ± 22.4 (743.0–1324.1) | 1273.1 ± 50.5 (931.9–2792.6) |
| LC90 ± SE (LCL–UCL) | 2157.1 ± 117.6 (1428.7–7232.3) | 1456.6 ± 43.9 (1121.4–2270.5) | 2850.1 ± 212.8 (1653.3–31863.3) | 1504.9 ± 44.4 (1167.7–2348.0) | 1974.8 ± 94.0 (1371.7–4830.2) |
| d. f. | 4 | 4 | 4 | 4 | 4 |
| χ2 | 1.93 | 8.87 | 3.72 | 1.87 | 3.12 |
| ΔG | rmsd | E.vdw | E.Int | E.H.B | Eele | |
|---|---|---|---|---|---|---|
| 6ARY | ||||||
| Palmatine | −4.781 | 1.991 | 8.934 | −5.667 | −7.385 | −24.345 |
| Jatrorrhizine | −4.725 | 1.012 | 28.914 | −14.683 | −8.725 | −18.975 |
| Columbamine | −4.560 | 1.731 | 2.230 | −5.482 | −7.739 | −21.639 |
| β-sitosterol | −4.785 | 1.450 | 46.606 | −4.659 | −5.647 | −21.316 |
| Difluoromethyl | −5.01 | 1.12 | 33.26 | −11.259 | −5.78 | −28.168 |
| 1JLV | ||||||
| Palmatine | −6.489 | 1.761 | 16.228 | −17.429 | −11.193 | −30.506 |
| Jatrorrhizine | −6.256 | 1.971 | 6.333 | −14.474 | −18.195 | −25.802 |
| Columbamine | −6.051 | 1.774 | 25.000 | −11.036 | −14.145 | −21.342 |
| β-sitosterol | −6.798 | 1.168 | 8.083 | −16.394 | −13.908 | −31.302 |
| Glutathione | −5.387 | 1.89 | 6.45 | −10.67 | −11.61 | −32.544 |
| Compound No. | EHOMO (eV) | ELUMO (eV) | HL Gaps (eV) |
|---|---|---|---|
| Palmatine | −0.1074 | −0.0352 | 0.0722 |
| Jatrorrhizine | −0.1891 | −0.0362 | 0.1529 |
| Columbamine | −0.1072 | −0.0356 | 0.0716 |
| β-sitosterol | −0.2279 | −0.0279 | 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Selim, T.A.; Abd-El Rahman, I.E.; Mahran, H.A.; Adam, H.A.M.; Imieje, V.; Zaki, A.A.; Bashar, M.A.E.; Hwihy, H.; Hamed, A.; Elhenawy, A.A.; et al. Mosquitocidal Activity of the Methanolic Extract of Annickia chlorantha and Its Isolated Compounds against Culex pipiens, and Their Impact on the Non-Target Organism Zebrafish, Danio rerio. Insects 2022, 13, 676. https://doi.org/10.3390/insects13080676
Selim TA, Abd-El Rahman IE, Mahran HA, Adam HAM, Imieje V, Zaki AA, Bashar MAE, Hwihy H, Hamed A, Elhenawy AA, et al. Mosquitocidal Activity of the Methanolic Extract of Annickia chlorantha and Its Isolated Compounds against Culex pipiens, and Their Impact on the Non-Target Organism Zebrafish, Danio rerio. Insects. 2022; 13(8):676. https://doi.org/10.3390/insects13080676
Chicago/Turabian StyleSelim, Tharwat A., Ibrahim E. Abd-El Rahman, Hesham A. Mahran, Hamza A. M. Adam, Vincent Imieje, Ahmed A. Zaki, Mansour A. E. Bashar, Hossam Hwihy, Abdelaaty Hamed, Ahmed A. Elhenawy, and et al. 2022. "Mosquitocidal Activity of the Methanolic Extract of Annickia chlorantha and Its Isolated Compounds against Culex pipiens, and Their Impact on the Non-Target Organism Zebrafish, Danio rerio" Insects 13, no. 8: 676. https://doi.org/10.3390/insects13080676