Phylogeny of the Chinese Subgenera of the Genus Homoneura (Diptera, Lauxaniidae, Homoneurinae) Based on Morphological Characters


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Morphological Study and Terminology
2.2. Specimens Examined and Morphological Characters

2.3. List of Characters Used in the Cladistic Analysis
- Frons: (0) flat; (1) uplifted; (2) sunken.
- Spot on anterior margin of frons: (0) absent; (1) present.
- Width of ventral margin of face/height of gena: (0) ≥ 3X; (1) < 3X.
- Length of gena/length of eye: (0) ≤ 1; (1) > 1.
- Color of proboscis: (0) black; (1) brown to yellow (Figure 2N).

- 29.
- 30.
- 31.
- 32.
- 33.

- 34.
- 35.
- 36.
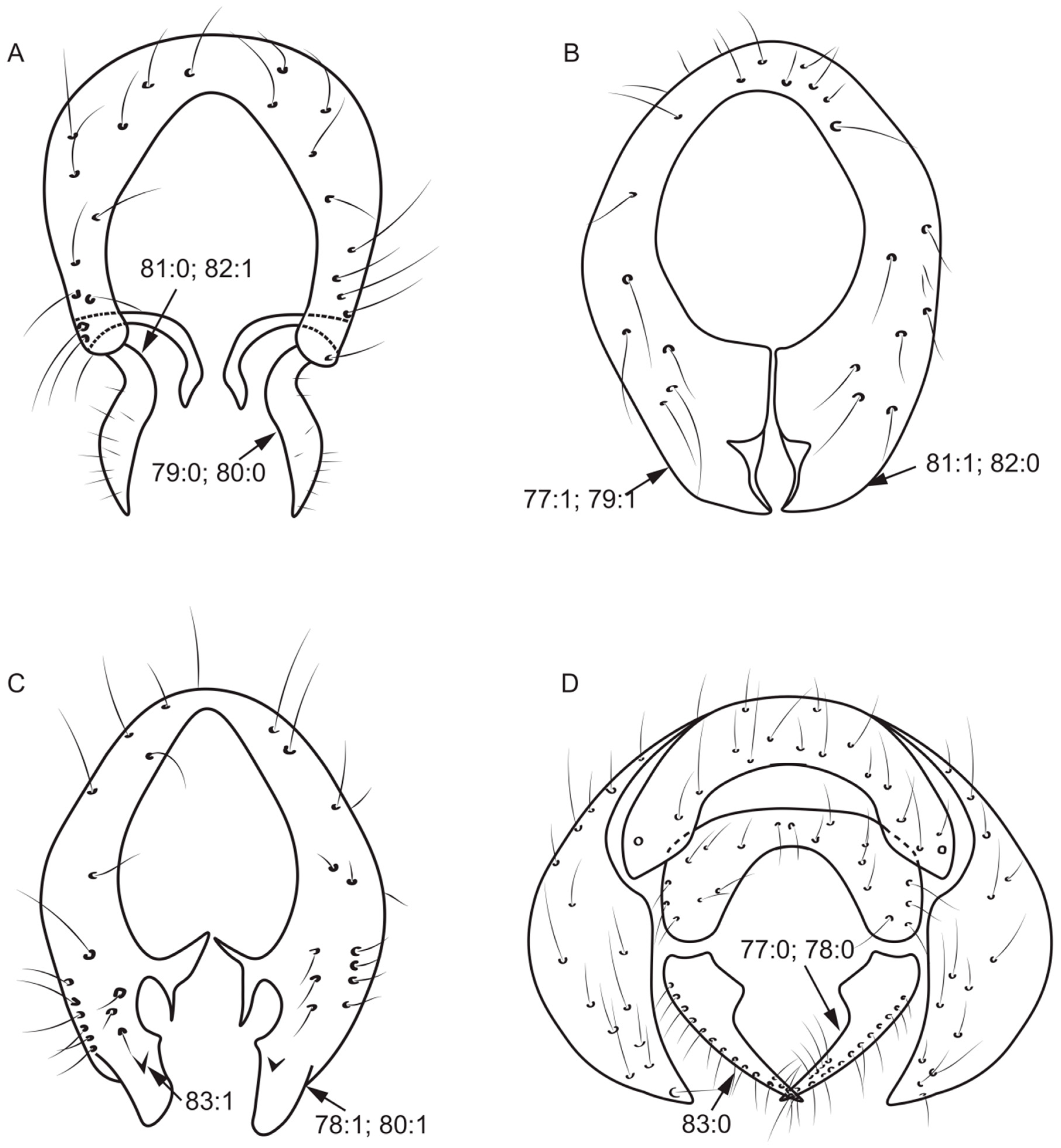
- Prescutellar acrostichal seta: (0) present (Figure 3A); (1) absent.
- 37.
- Intra-alar seta: (0) present; (1) absent.
- 38.
- 39.
- Length of leg/length of body: (0) ≤ 1; (1) > 1.
- 40.
- 41.
- Ctenidium short seta in fore femur: (0) absent; (1) no more than ten; (2) more than ten (Figure 3G).
- 42.
- Anterior seta in mid femur: (0) more than six; (1) no more than six (Figure 3I).
- 43.
- Posterior ventral seta in mid femur: (0) absent; (1) present.
- 44.
- 45.
- 46.
- Preapical anterior dorsal seta in hind femur; (0) present (Figure 3H); (1) absent.
- 47.
- Anteroventral seta in hind femur: (0) absent; (1) present (Figure 3H).
- 48.
- 49.
- Area of transparent area or light-yellow area of wing/area of wing spot area: (0) > 1; (1) ≤ 1.
- 50.
- Area of transparent or light-yellow area above wing R4+5/area of wing spot above R4+5: (0) > 1; (1) ≤ 1.
- 51.
- 52.
- 53.
- 54.
- 55.
- 56.
- 57.
- 58.
- 59.
- 60.
- 2nd (between R1 and R2+3) section/3rd (between R2+3 and R4+5) section: (0) ≥ 3X; (1) < 3X.
- 61.
- 3rd (between R2+3 and R4+5) section/4th (between R4+5 and M1) section: (0) ≥ 1.5X; (1) < 1.5X.
- 62.
- Length of the ultimate section of CuA1/length of the penultimate section of CuA1; (0) < 1/5; (1) ≥ 1/5.
- 63.
- 64.
- Color of knob part of haltere: (0) black; (1) brown or yellow.

- 65.
- Color of tergites 3–6: (0) black; (1) brown or yellow.
- 66.
- 67.
- 68.
- 69.
- 70.
- 71.

- 72.
- 73.
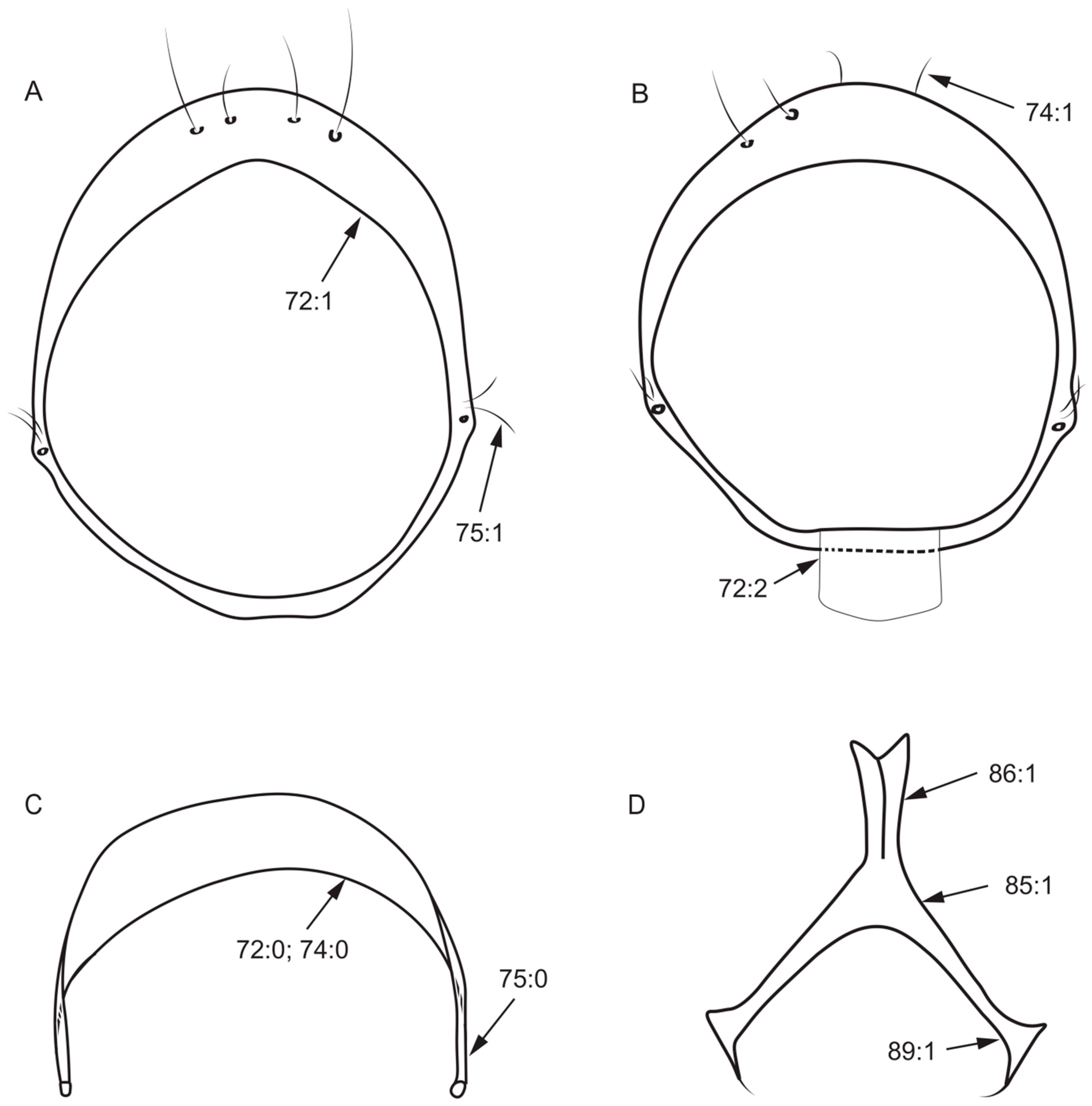
- Length of dorsal margin of syntergosternite/length of posterior margin of syntergosternite: (0) < 1/2; (1) ≥ 1/2.
- 74.
- 75.
- 76.
- Length of dorsal margin of epandrium/length of ventral margin of epandrium: (0) > 1/2; (1) ≤ 1/2.

- 77.
- 78.
- 79.
- 80.
- 81.

- 82.
- 83.
- 84.
- Surstylus: (0) without seta; (1) with seta.
- 85.
- 86.
- 87.
- Both sides anterior margin of hypandrium: (0) without inner processes; (1) with inner processes (Figure 8A).
- 88.

- 89.
- Both sides posterior margin of hypandrium: (0) without ventral process; (1) with ventral process (Figure 6D).
- 90.
- 91.
- 92.
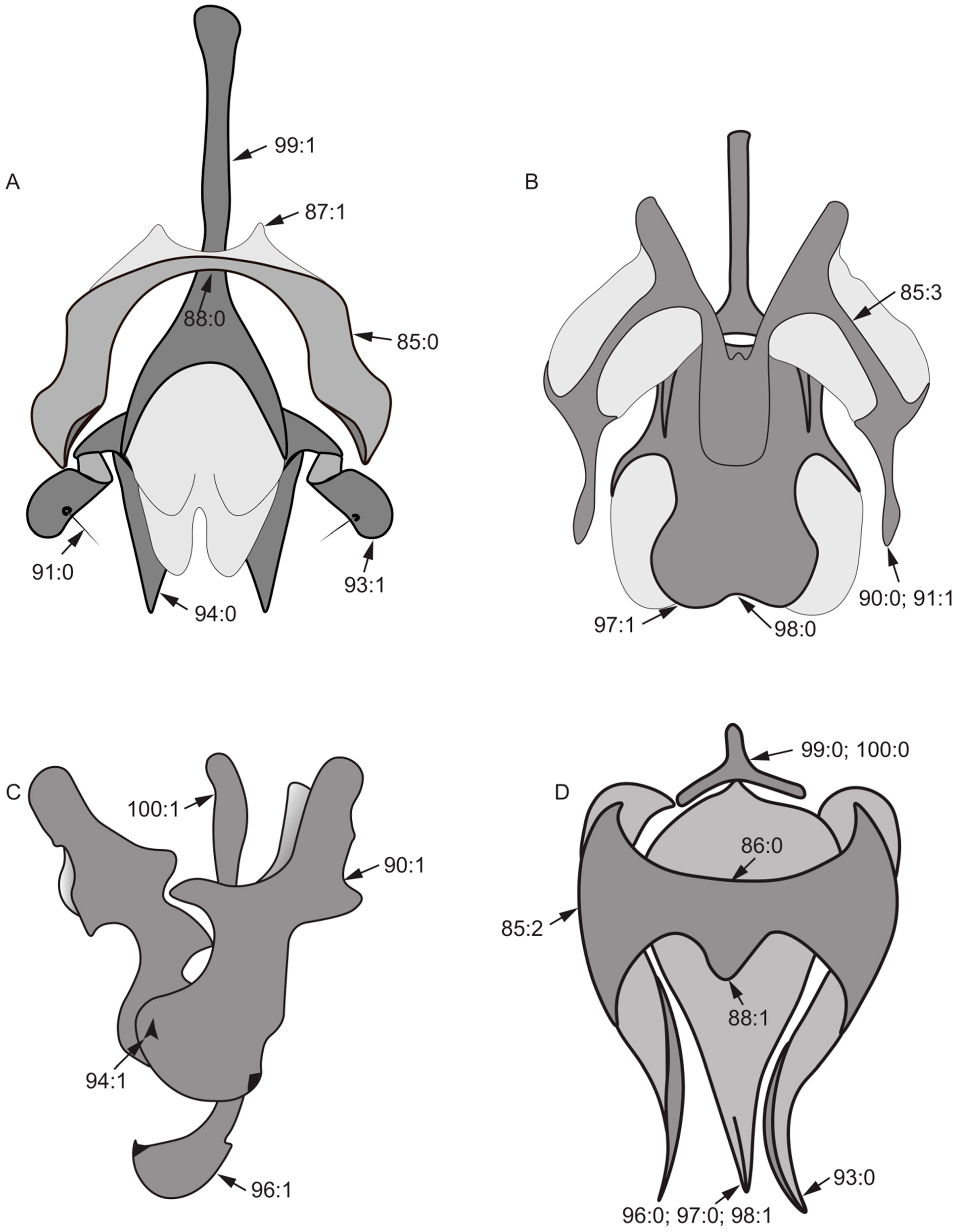
- Length of gonite/length of phallus: (0) ≥ 1/2; (1) < 1/2.
- 93.
- 94.
- 95.
- Lateral view of the top of phallus: (0) bent; (1) straight.
- 96.
- 97.
- 98.
- 99.
- 100.
- 101.
- 102.
- 103.
- 104.
- Both sides of posterior margin of sternite 8: (0) with process; (1) without process.
- 105.
- Posterior margin of tergite 9: (0) uplifted; (1) sunken.

2.4. Cladistic Analysis
3. Results
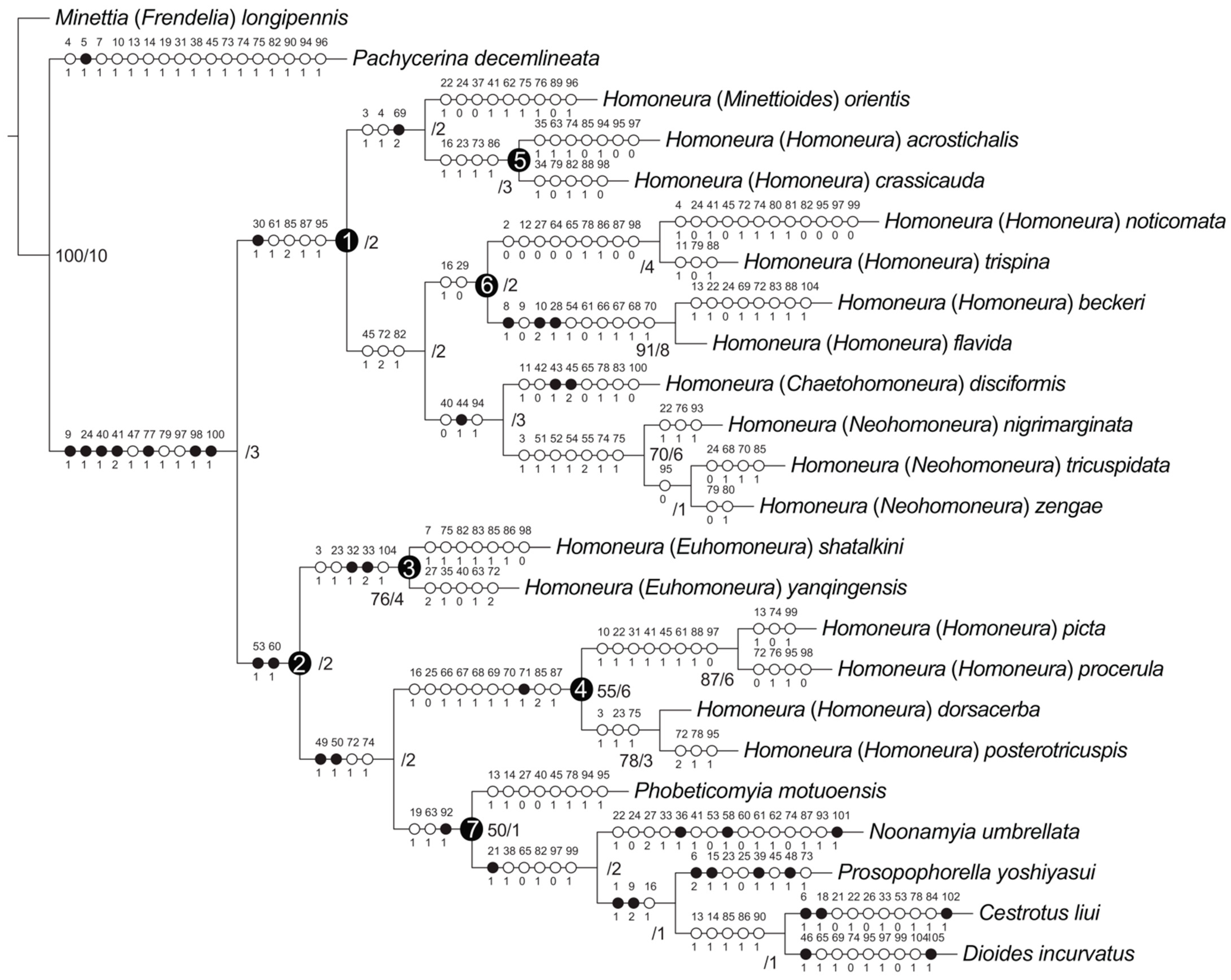
3.1. Phylogenetic Analysis
3.2. Subfamily Homoneurinae
3.3. The Genus Homoneura and Allies
3.4. Monophyly and Relationships among Chinese Subgenera of Homoneura
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Li, W.L.; Qi, L.; Yang, D. Four new species of the subfamily Homoneurinae (Diptera, Lauxaniidae) from southwestern China. ZooKeys 2020, 953, 119–136. [Google Scholar] [CrossRef] [PubMed]
- Dvořáková, K. The results of a preliminary survey of the families Lauxaniidae and Heleomyzidae (Diptera, Acalyptrata) in the Upper and Lower Palatinate forest. Erica 2008, 15, 108–112. [Google Scholar]
- Shewell, G.E. Lauxaniidae. In Manual of Nearctic Diptera McAlpine; McAlpine, J.F., Peterson, B.V., Shewell, G.E., Teskey, H.J., Vockeroth, J.R., Wood, D.M., Eds.; Agriculture Canada: Ottawa, ON, Canada, 1987; Volume 2, pp. 951–964. [Google Scholar]
- Broadhead, E.C. The species of Poecilominettia, Homoeominettia and Floriminettia (Diptera: Lauxaniidae) in Panama. Bull. Br. Mus. (Nat. Hist.) Entomol. 1989, 58, 185–226. [Google Scholar]
- Hering, E. Biology of the Leaf Miners; Springer Netherlands: Berlin, Germany, 1951; pp. 1–420. [Google Scholar]
- Papp, L.; Shatalkin, A.I. Family Lauxaniidae. In Manual of Palaearctic Diptera; Papp, L., Darvas, B., Eds.; Science Herald: Budapest, Hungary, 1998; Volume 3, pp. 383–400. [Google Scholar]
- Silva, V.C.; Mello, R.L. Occurrence of Physoclypeus farinosus Hendel (Diptera: Lauxaniidae) in flowerheads of Asteraceae (Asterales). Neotrop. Entomol. 2008, 37, 92–96. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, R.M.; Foote, B.A. Biology and immature stages of eight species of Lauxaniidae (Diptera) II. Descriptions of immature stages and discussion of larval feeding habits and morphology. P. Entomol. Soc. Wash. 1976, 78, 16–37. [Google Scholar]
- Moretti, T.C.; Ribeiro, O.B.; Thyssen, P.J.; Solis, D.R. Insects on decomposing carcasses of small rodents in a secondary forest in Southeastern Brazil. Eur. J. Entomol. 2008, 105, 691–696. [Google Scholar] [CrossRef] [Green Version]
- Shewell, G.E. Family Lauxaniidae. In A Catalogue of the Diptera of America North of Mexico; Stone, A., Sabrosky, C.W., Wirth, W.W., Foote, R.H., Coulson, J.R., Eds.; United States Department of Agriculture: Washington, DC, USA, 1965; pp. 696–704. [Google Scholar]
- Reddersen, J. Feeding biology of fungivorous insects from Danish cereal fields. Pedobiologia 1995, 39, 370–384. [Google Scholar]
- Stuckenberg, B.R. A review of the Old World genera of Lauxaniidae (Diptera). Ann. Natal Mus. 1971, 20, 499–610. [Google Scholar]
- Kim, S.P. Australian Lauxaniid Flies. Revision of the Australian Species of Homonera van der Wulp, Trypetisoma Malloch, and Allied Genera (Diptera, Lauxaniidae). In Monographs on Invertebrate Taxonomy; Morton, D.W., Nielsen, E.S., Reekie, P.W., Eds.; Commonwealth Scientific and Industrial Research Organisation: Canberra, Australia, 1994; Volume 1, pp. 1–445. [Google Scholar]
- Shi, L.; Zhang, M.J.; Shen, R.R.; Li, S.D.; Bai, J.; Wang, Y.J.; He, G.W.; Cui, Y. Preliminary study on phylogeny of species groups in subgenus Homoneura. J. Environ. Entomol. 2017, 39, 351–356. [Google Scholar]
- Cumming, J.M.; Wood, D.M. Adult morphology and terminology. In Manual of Afrotropical Diptera; Kirk-Spriggs, A.H., Sinclair, B.J., Eds.; South African National Biodiversity Institute: Pretoria, South Africa, 2017; Volume 1, pp. 89–133. [Google Scholar]
- Gaimari, S.D.; Silva, V.C. Lauxaniidae (Lauxaniid flies). In Manual of Afrotropical Diptera; Kirk-Spriggs, A.H., Sinclair, B.J., Eds.; South African National Biodiversity Institute: Pretoria, South Africa, 2021; Volume 3, pp. 1757–1781. [Google Scholar]
- Goloboff, P.A. NONA ver. 2; Goloboff, P.A.: Tucuman, Argentina, 1999. [Google Scholar]
- Goloboff, P.A.; Farris, J.S.; Nixon, K.C. TNT, a free program for phylogenetic analysis. Cladistics 2008, 24, 774–786. [Google Scholar] [CrossRef]
- Bremer, K. Branch support and tree stability. Cladistics 1994, 10, 295–304. [Google Scholar] [CrossRef]
- Nixon, K.C. WinClada; ver. 1.00.08; Nixon, K.C.: Ithaca, NY, USA, 2002. [Google Scholar]
- Sasakawa, M. Lauxaniidae (Diptera) of Malaysia (part 2): A revision of Homoneura van der Wulp. Insect. Mats. 1992, 46, 133–210. [Google Scholar]
- Shi, L.; Wang, J.C.; Yang, D. Nine new species from China of the subgenera Chaetohomoneura and Neohomoneura in the genus Homoneura (Diptera, Lauxaniidae). Zootaxa 2011, 2975, 1–28. [Google Scholar] [CrossRef] [Green Version]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kong, C.; Feng, K.; Zhao, S.; Li, W.; Li, X. Phylogeny of the Chinese Subgenera of the Genus Homoneura (Diptera, Lauxaniidae, Homoneurinae) Based on Morphological Characters. Insects 2022, 13, 665. https://doi.org/10.3390/insects13080665
Kong C, Feng K, Zhao S, Li W, Li X. Phylogeny of the Chinese Subgenera of the Genus Homoneura (Diptera, Lauxaniidae, Homoneurinae) Based on Morphological Characters. Insects. 2022; 13(8):665. https://doi.org/10.3390/insects13080665
Chicago/Turabian StyleKong, Chaoyang, Keli Feng, Shengjuan Zhao, Wenliang Li, and Xuankun Li. 2022. "Phylogeny of the Chinese Subgenera of the Genus Homoneura (Diptera, Lauxaniidae, Homoneurinae) Based on Morphological Characters" Insects 13, no. 8: 665. https://doi.org/10.3390/insects13080665