
Sex Pheromone of Cocoa Pod Borer, Conopomorpha cramerella: Field Activity Evaluation of Pheromone Formulations in an Indonesia Plantation


{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Lure Preparation
2.2. Field Location
2.3. Field Experiments
2.4. Statistical Analysis
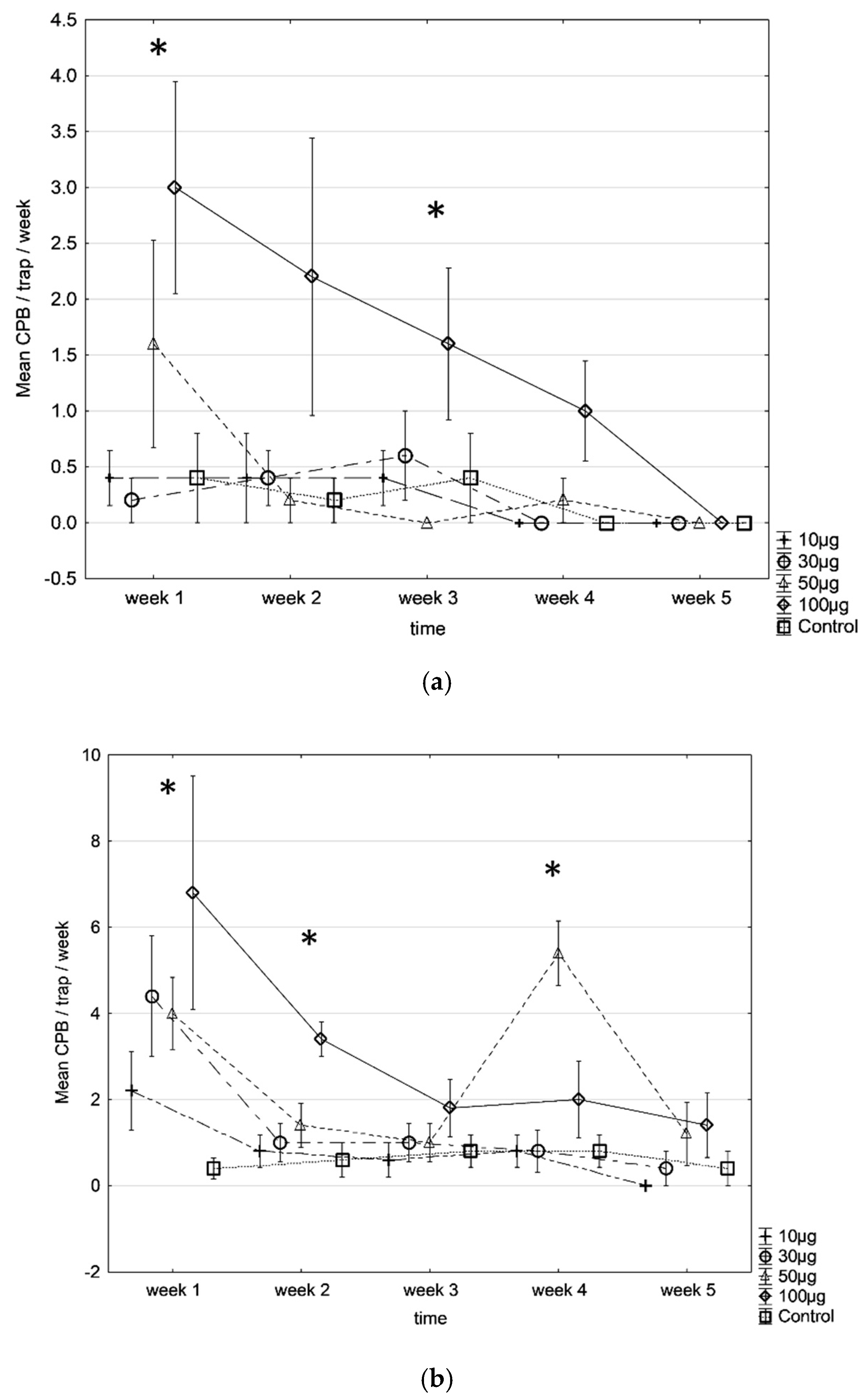
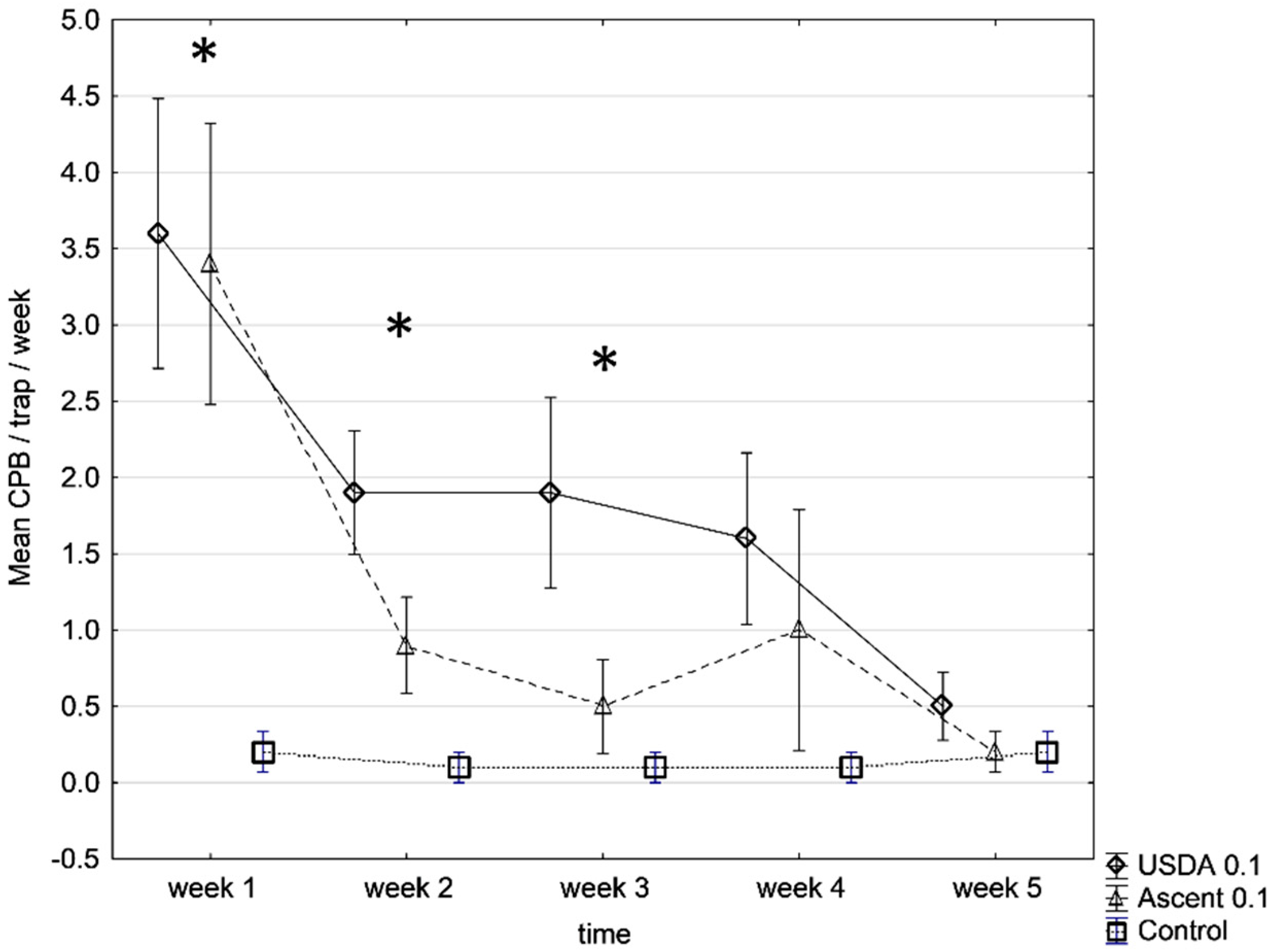
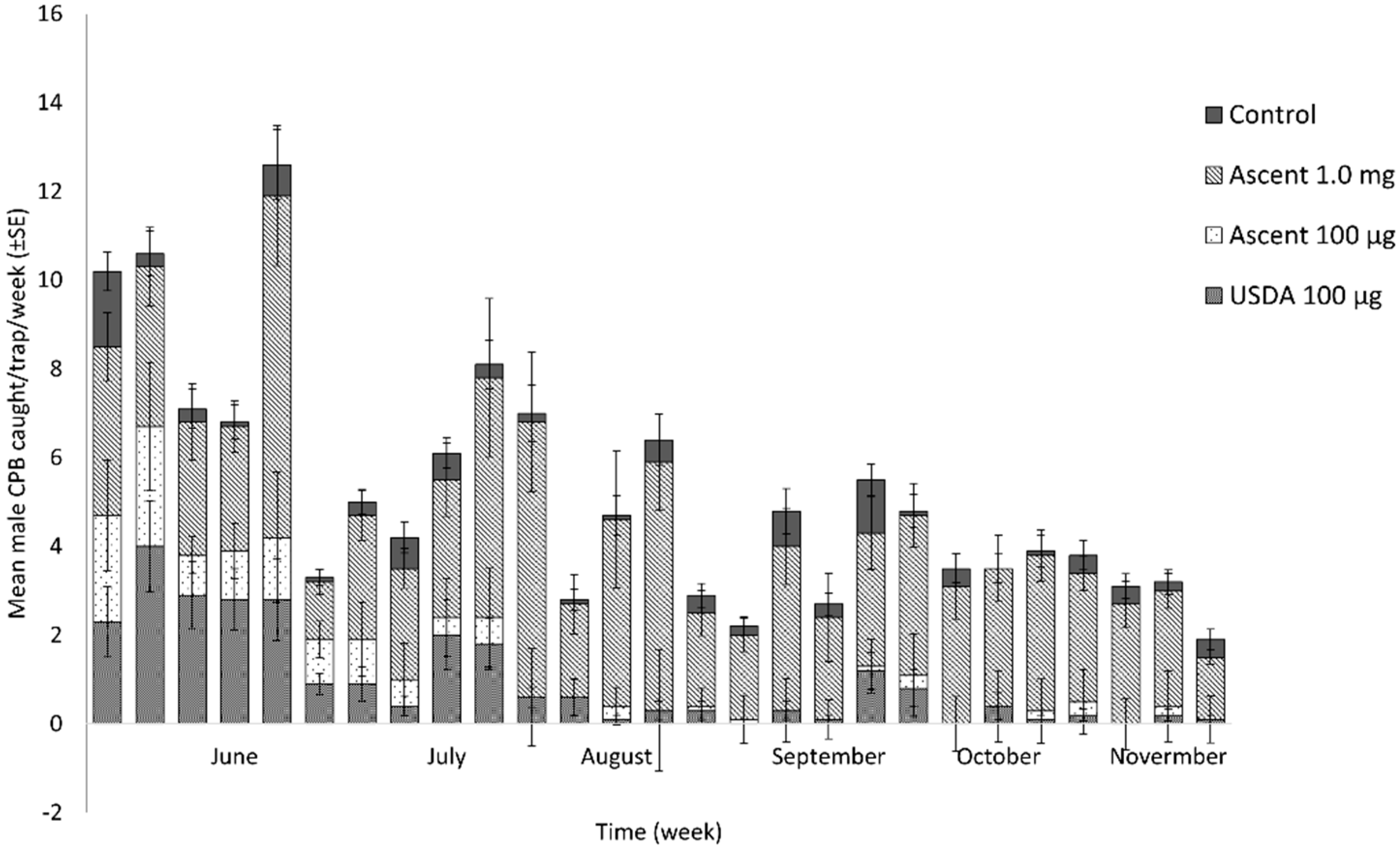
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beevor, P.S.; Cork, A.; Hall, D.R.; Nesbitt, B.F.; Day, R.K.; Mumford, J.D. Components of female sex pheromone of cocoa pod borer moth, Conopomorpha cramerella. J. Chem. Ecol. 1986, 12, 178. [Google Scholar] [CrossRef]
- Beevor, P.S.; Day, R.K.; Mumford, J.D. Female sex pheromone of the cocoa pod borer moth, Acrocercops cramerella: Identification and field evaluation. In Cocoa and Coconut: Progress and Outlook; Incorporated Society of Planters: Kuala Lumpur, Malaysia, 1986; pp. 287–292. [Google Scholar]
- Ho, S.H.; Beevor, P.S.; Mumford, J.D. A practical approach to the control of the cocoa pod borer moth using synthetic sex pheromone in an integrated system. In Management of the Cocoa Pod Borer; Ooi, P.A.C., Chan, L.G., Khoo, K.C., Teoh, C.H., Jusoh, M., Ho, C.T., Lim, G.S., Eds.; Malaysia Plant Protection Society: Kuala Lumpur, Malaysia, 1987; pp. 53–68. [Google Scholar]
- Beevor, P.S.; Mumford, J.D.; Shah, S.; Day, R.K.; Hall, D.R. Observations on pheromone-baited mass trapping for control of cocoa pod borer, Conopomorpha cramerella, in Sabah, East Malaysia. Crop Protect. 1993, 12, 134–140. [Google Scholar] [CrossRef]
- Matlick, B.K. Machete technology: What small cocoa farmers need! Review of pod borer control methods by small farmers in Indonesia. In Proceedings of the First International Workshop on Sustainable Cocoa Growing, Balboa Ancon, Panama, 29 March–3 April 1998; Smithsonian Tropical Research Institute: Balboa Ancon, Panama, 1998; Available online: https://nationalzoo.si.edu/scbi/migratorybirds/research/cacao/matlick.cfm (accessed on 21 July 2019).
- Zhang, A.; Kuang, L.F.; Maisin, N.; Karmuru, B.; Hall, D.R.; Virdiana, I.; Lamber, S.; Purung, H.B.; Wang, S.; Hebbar, P. Activity of cocoa pod borer sex pheromone in cocoa fields. Environ. Entomol. 2008, 37, 719–724. [Google Scholar] [CrossRef] [Green Version]
- Niogret, J.; Ekayanti, A.; Ingram, K.; Lambert, S.; Kendra, P.E.; Alborn, H.; Epsky, N.D. Development and behavioral ecology of Conopomorpha cramerella (Lepidoptera: Gracillariidae). Fla. Entomol. 2019, 102, 382–387. [Google Scholar] [CrossRef] [Green Version]
- Niogret, J.; Ekayanti, A.; Kendra, P.E.; Ingram, K.; Lambert, S.; Espky, N.D.; Marelli, J.-P. Host preferences of the cocoa pod borer, Conopomorpha cramerella, the main threat to cocoa production in Southeast Asia. Entomol. Exp. Appl. 2020, 168, 221–227. [Google Scholar] [CrossRef]
- Vos, J.G.M.; Ritchie, B.J.; Flood, J. Discovery Learning about Cacao: An Inspirational Guide for Training Facilitators; CABI Bioscience: Wallingford, UK, 2003. [Google Scholar]
- Rosmana, A.; Daha, L.; Ridayani, A.; Gassa, A.; Urling, A. Persistency and penetration of entomopathogenic nematode Steinernema carpocapsae on surface of cocoa pod and its infectivity to cocoa pod borer Conopomorpha cramerella (Lepidoptera, Gracillariidae). Agritrop 2009, 28, 161–168. [Google Scholar]
- Lim, G.T.; Pan, K.Y. Observations on the Sexual Activity and Egg Production of Cocoa pod Borer Conopomorpha cramerella (Snellen) in the Laboratory; Annual Research Report; Department of Agriculture: Kota Kinabalu, Malaysia, 1986. [Google Scholar]
- Anshary, A.; Basir-Cyio, M.; Hasanah, U.; Mahfudz; Saleh, S.; Edy, N.; Pasaru, F. Applications of biological agents and pruning effectively control cocoa pod borer. Asian J. Crop Sci. 1994, 9, 125–132. [Google Scholar] [CrossRef] [Green Version]
- Saripah, B.; Azhar, I. Five years of using cocoa black ants, to control cocoa pod borer at farmer plot—An epilogue. Malays. Cocoa J. 2012, 7, 8–14. [Google Scholar]
- See, Y.A.; Koo, K.C. Influence of Dolichoderus thoracicus (Hymenoptera: Formicidae) on cocoa pod damage by Conopomorpha cramerella (Lepidoptera: Gracillariidae) in Malaysia. Bull. Entomol. Res. 1996, 86, 467–474. [Google Scholar] [CrossRef]
- Rosmana, A.; Shepard, M.; Hebbar, P.; Mustari, A. Control of cocoa pod borer and phytophthora pod rot using degradable plastic pod sleeves and nematode, Steinernema carpocapsae. Indones. J. Agric. Sci. 2010, 11, 41–47. [Google Scholar] [CrossRef]
- Vanialingam, T.; Easaw, P.T.; Irshad, M.; Manals, J.; Cruz, P. Early work in the control of the cocoa pod borer Acrocercops cramerella in the Philippines. In Proceedings of the 8th International Cocoa Research Conference, Cartagena, Colombia, 18–23 October 1981; pp. 345–351. [Google Scholar]
- Youdeowei, A. Review of Cocoa Pests in Indonesia and the Philippines and the development of research strategies for cocoa pest management. In Proceedings of the Developing Bulk Cocoa in Indonesia, Report on a Visit, Mimeo, Jakarta, Indonesia, 7 February–14 March 1980; pp. 1–64. [Google Scholar]
- Lim, G.T. Biology, ecology and control of cocoa pod borer Conopomorpha cramerella (Snellen). In Cocoa Pest and Disease Management in Southeast Asia and Australia; Keane, P.J., Putter, C.A.J., Eds.; FAO Plant Production and Protection Paper; FAO: Rome, Italy, 1992; Volume 22, pp. 85–100. [Google Scholar]
- Wood, B.J.; Fee, C.G.; Cheong, S.S.; Foh, C.C. Trials on control of the cocoa pod borer Conopomorpha cramerella (Snellen) in Sabah by regular complete harvesting. Int. J. Pest Manag. 1992, 38, 271–278. [Google Scholar]
- Teh, C.-L.; Panga, J.T.-Y.; Ho, C.-T. Variation of the response of clonal cocoa to attack by cocoa pod borer Conopomorpha cramerella (Lepidoptera: Gracillariidae) in Sabah. Crop Protect. 2006, 25, 712–717. [Google Scholar] [CrossRef]
- Wiese, M.V. Crop management buy comprehensive appraisal of yield determining variables. Annu. Rev. Phytopathol. 1982, 23, 455–473. [Google Scholar]
- Valenzuela, I.; Bin Purung, H.; Roush, R.T.; Hamilton, A.J. Practical yield loss models for infestation of cocoa with cocoa pod borer moth, Conopomorpha cramerella (Snellen). Crop Protect. 2014, 66, 19–28. [Google Scholar] [CrossRef]
- Chiarappa, L. Crop Loss Assessment Methods; Supplement 3; Commonwealth Agricultural Bureaux: Famham Royal, UK, 1981. [Google Scholar]
- Walker, P.T. Crop diseases: The need to quantify the effects of pests, diseases and weeds on agricultural production. Agric. Ecosyst. Environ. 1983, 9, 119–158. [Google Scholar] [CrossRef]
- Zadoks, J.C. On the conceptual basis of crop loss assessment: The threshold theory. Annu. Rev. Phytopathol. 1985, 23, 455–473. [Google Scholar] [CrossRef]
- Vanhole, W.; Feng, Y.; Yu, M.; Hafiz, I.O.; Vanhoudt, N.; Van Damme, P.L.J.; Zhang, A. Evaluation of attract-and-kill strategy for management of cocoa pod borer, Conopomorpha cramerella, in Malaysia coca plantation. Int. J. Pest Manag. 2020, 66, 155–162. [Google Scholar] [CrossRef]
- Yamamura, K.; Yokozawa, M.; Nishimori, M.; Ueda, Y.; Yokosuka, T. How to analyse long-term insect population dynamics under climate change: 50-year data of three insect pests in paddy fields. Popul. Ecol. 2006, 48, 31–48. [Google Scholar] [CrossRef]
- Niogret, J. (Mars Wrigley, Smithfield, QLD, Australia). Unpublished Work. 2022. [Google Scholar]
- Niogret, J. (Mars Wrigley, Smithfield, QLD, Australia). Unpublished Data. 2018. [Google Scholar]
- Vanhove, W.; Vanhoudt, N.; Bhanu, K.R.M.; Abubeker, S.; Feng, Y.; Yu, M.; Van Damme, P.; Zhang, A. Geometric isomers of sex pheromone components do not affect attractancy of Conopomorpha cramerella in cocoa plantations. J. Appl. Entomol. 2015, 139, 660–668. [Google Scholar] [CrossRef]
- Mumford, J.D.; Beevor, P.S. Cost effectiveness of pheromone baited mass trapping in integrated control of cocoa pod borer. In Management of the Cocoa Pod Borer; Ooi, P.A.C., Chan, L.G., Khoo, K.C., Teoh, C.H., Jusoh, M., Ho, C.T., Lim, G.S., Eds.; Malaysian Plant Protection Society: Kuala Lumpur, Malaysia, 1987; pp. 167–170. [Google Scholar]
- Alias, A.; Sadao, W.; Tay, E.B. Efficacy of mating disruption using synthetic pheromone for the management of cocoa pod borer, Conopomorpha cramerella (Snellen) (Lepidoptera: Gracillariidae). Malays. Cocoa J. 2004, 1, 46–52. [Google Scholar]
- Preti, M.; Knight, A.L.; Favaro, R.; Basoalto, E.; Tasin, M.; Angeli, S. Comparison of New Kairomone-based lures for Cydia pomonella (Lepidoptera: Tortricidae) in Italy and USA. Insects 2021, 12, 72. [Google Scholar] [CrossRef] [PubMed]
- Knight, A.L.; Preti, M.; Basoalto, E.; Mujica, M.V.; Favaro, R.; Angeli, S. Combining female removal with mating disruption for management of Cydia pomonella in apple. Entomol. Gen. 2022, 42, 309–321. [Google Scholar] [CrossRef]



Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Niogret, J.; Ekayanti, A.; Zhang, A. Sex Pheromone of Cocoa Pod Borer, Conopomorpha cramerella: Field Activity Evaluation of Pheromone Formulations in an Indonesia Plantation. Insects 2022, 13, 663. https://doi.org/10.3390/insects13080663
Niogret J, Ekayanti A, Zhang A. Sex Pheromone of Cocoa Pod Borer, Conopomorpha cramerella: Field Activity Evaluation of Pheromone Formulations in an Indonesia Plantation. Insects. 2022; 13(8):663. https://doi.org/10.3390/insects13080663
Chicago/Turabian StyleNiogret, Jerome, Arni Ekayanti, and Aijun Zhang. 2022. "Sex Pheromone of Cocoa Pod Borer, Conopomorpha cramerella: Field Activity Evaluation of Pheromone Formulations in an Indonesia Plantation" Insects 13, no. 8: 663. https://doi.org/10.3390/insects13080663