
Assessing the Diversity and Systematics of Brachyopini Hoverflies (Diptera: Syrphidae) in the Iberian Peninsula, Including the Descriptions of Two New Species †


Abstract
:Simple Summary
Abstract
1. Introduction
2. Material and Methods
2.1. Examined Material
2.2. Morphological Study
2.3. Molecular Study
3. Results
3.1. New Brachyopini Species
- Holotype
- Paratypes
- Holotype
- Paratypes
3.2. Chrysogaster coerulea Strobl in Czerny and Strobl, 1909 stat. n.
3.3. The Holotype of Orthonevra incisa (Loew, 1843)
3.4. The Lectotype of Orthonevra plumbago (Loew, 1840)
3.5. Key to the Studied Brachyopini genera
- Wing: basal section of radial vein (Figure 12C) with some long hairs dorsally … 2
- -
- Wing: basal section of radial vein (Figure 12C) bare, without hairs … 3
- Male holoptic and female dichoptic; ventral side of metafemur with black setulae, at least apically; face, from antennae to mouth edge, flat or slightly convex in lateral view; basoflagellomere elongate, 1.5-4× longer than width … Riponnensia (go to Section 3.5.5)
- -
- Male and female dichoptic; metafemur without setulae; face, from antennae to mouth edge, concave in lateral view; basoflagellomere large, roundish to oval, up to 1.5× longer than width … Lejogaster (go to Section 3.5.2)
- Lateral margins of tergum I entirely shiny; vein M1 usually recessive (as in Figure 6A); sternum I usually shiny … Orthonevra (go to Section 3.5.4)
- Basoflagellomere brownish black (Figure 2A); tergum II with short to very short hairs on its central area; in male, face sometimes slightly concave, with a conspicuous medial tubercle (Figure 2A,C) … Melanogaster (go to Section 3.5.3)
- -
- Basoflagellomere orange to red (Figure 10); tergum II often with long recumbent hairs on its central area; in males, face straight, usually with a less-conspicuous medial tubercle (Figure 10A,B), sometimes with a bumped area at each side … Chrysogaster (go to Section 3.5.1)
3.5.1. Chrysogaster Meigen, 1803
- Proepimeron completely pollinose … C. coemiteriorum
- -
- Proepimeron with a large central shiny area, sometimes barely pollinose … 2
- Sternum 2 with long hairs, of about two-thirds the length of the scutum hairs … C. rondanii
- -
- Sternum 2 only with short recumbent hairs, occasionally with a few longer hairs but never as long as in C. rondanii … 3
- Narrow-faced species; in frontal view, the lowest part of frons (female) is narrower than the maximum width of an eye or equal in width; males with frontal triangle of 90° or less; scutum, in males, with long inclined hairs at least on the anterior half … 4
- -
- Broad-faced species; in frontal view, the lowest part of frons (female) wider than the maximum width of an eye; males with frontal triangle of more than 90°; scutum, in male, with long nearly erect hairs at least on the anterior half … C. virescens
- Mesonotum shiny black; male with a small facial tubercle (Figure 10B) … C. basalis (females of this species cannot always be separated from C. coerulea, but females of C. coerulea usually have more bluish reflections than C. basalis)
- Chrysogaster basalis Loew, 1857
- Chrysogaster coemiteriorum (Linnaeus, 1758)
- Chrysogaster coerulea Strobl in Czerny and Strobl, 1909
- Chrysogaster rondanii Maibach and Goeldlin de Tiefenau, 1995
- Chrysogaster solstitialis (Fallén, 1817)
- Chrysogaster virescens Loew, 1854
3.5.2. Lejogaster Rondani, 1857
- Anterior anepisternum entirely bare; male basoflagellomere wider than long … L. metallina
- -
- Anterior anepisternum pilose postero-dorsally; male basoflagellomere as long and wide or slightly longer tan wide … L. tarsata
- Lejogaster metallina (Fabricius, 1781)
- Lejogaster tarsata(Meigen, 1822)
3.5.3. Melanogaster Rondani, 1857
- Eyes holoptic (males) … 2
- -
- Eyes dichoptic (females) … 5
- Scutum with sparse pilosity, especially noticeable on the posterior part; surstylus curved and less than two times as long as width … M. nuda
- Scutum mainly with brownish black to black hairs; superior lobe of hypandrium with a long tooth pointing opposite the epandrium [7] (figure 4) … M. aerosa
- -
- Scutum mainly with brown to white hairs; superior lobe of hypandrium with a long tooth pointing towards the epandrium (Figure 3A) … 4
- Scutum with brown hairs; head and thorax without blue reflections; surstylus with a conspicuously elongated finger-like process apically [9] (figure 20) … M. hirtella
- Scutum with very-short sparse pilosity, especially noticeable on the posterior part (hairs shorter than the diameter of a posterior ocellus) … M. nuda
- -
- Scutum with longer dense pilosity all over (hairs longer than the diameter of a posterior ocellus) … 6
- Scutum with short (length equalling the diameter of an ocellus) semi-recumbent hairs; metatibia with at least some black hairs … M. aerosa
- -
- Scutum with longer (length about twice the diameter of an ocellus) erect or suberect hairs; metatibia completely pale pilose … 7
- Head and thorax without blue reflections; wing membrane hyaline over all … M. hirtella
- Melanogaster aerosa (Loew, 1843)
- Melanogaster baetica sp. n.
- Melanogaster hirtella (Loew, 1843)
- Melanogaster nuda (Macquart, 1829)
3.5.4. Orthonevra Macquart, 1829
- Sternum I densely pollinose … 2
- -
- Sternum I shiny or slightly pollinose … 3
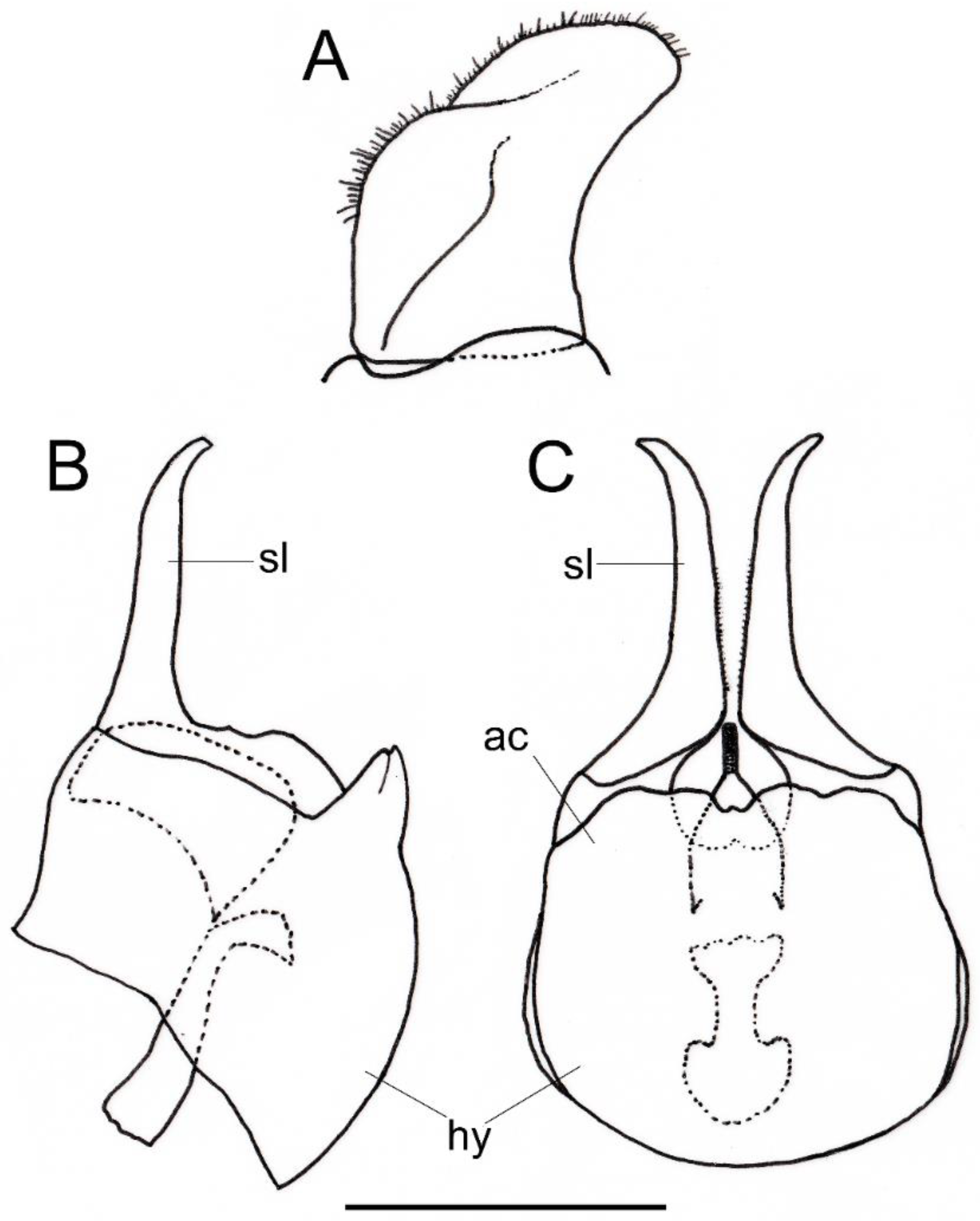
- Body, in general, with shorter hairs (Figure 11A,B); eye virtually bare; katepisternum with two separate patches of hairs, one dorsal and another ventral; metafemur entirely white pilose, at most with some scattered black hairs ventrally (Figure 12C) … O. incisa
- -
- Body, in general, with longer hairs (Figure 4); eye with very short but obvious scattered hairs; katepisternum with a third patch of hairs below the dorsal patch in such a way that dorsal and ventral patches might be connected or almost so; metafemur with black, sometimes spiny, black hairs ventrally … O. arcana sp. n.
- Basoflagellomere elongate, 1.5× longer than wide or more … 4
- -
- Basoflagellomere round to oval, less than 1.5× longer than wide … 5
- Basoflagellomere bar-shaped, with subparallel dorsal and ventral margins … O. frontalis
- -
- Basoflagellomere tapering towards the apex, with a pointing end dorso-apically (more conspicuous in female) … O. nobilis
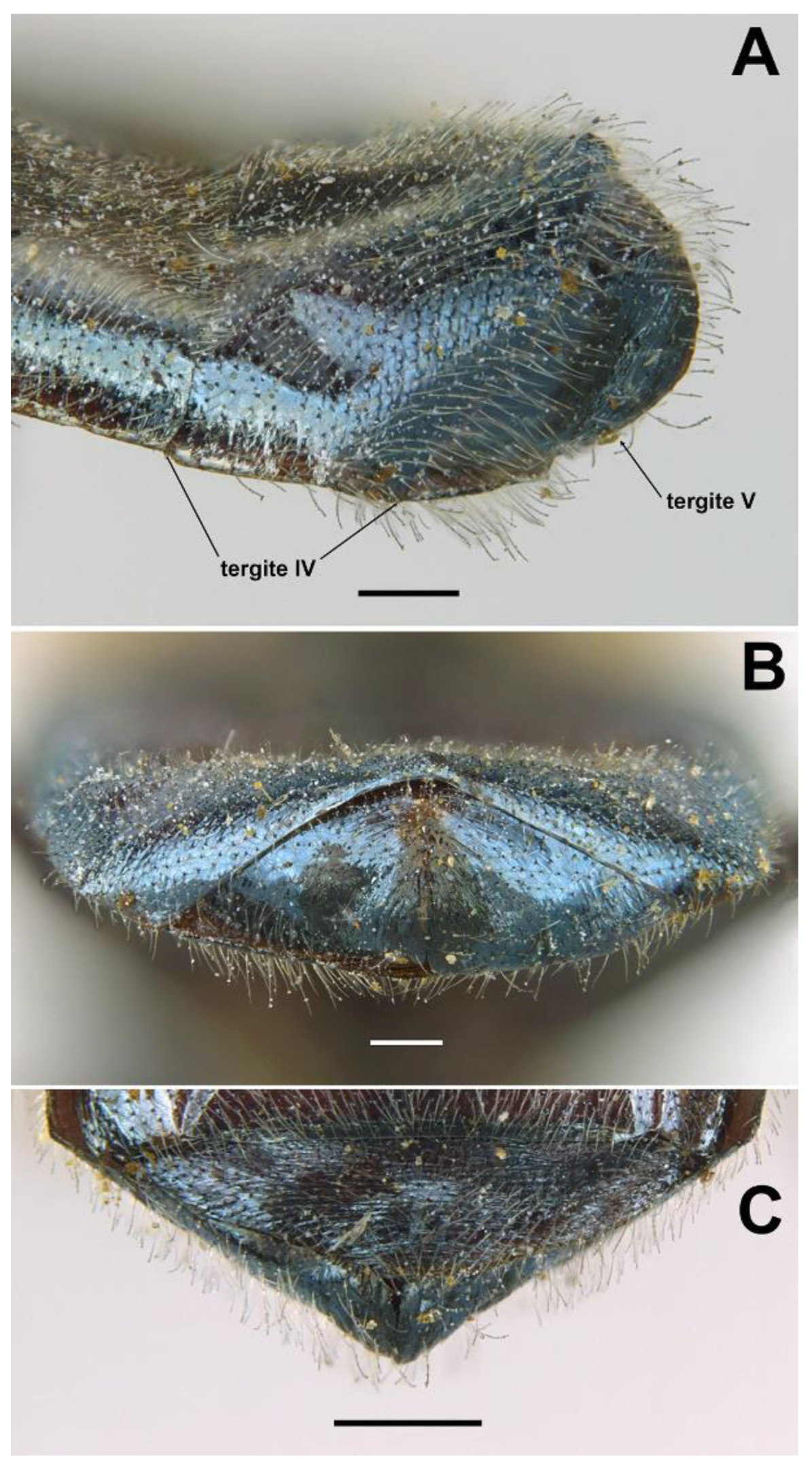
- Basoflagellomere 1.3× (Figure 16B). Only females: tergum IV with a small wart-like elevation in the middle of posterior margin; tergum V with a medial sulcus starting on the posterior margin as an incision and then extending almost to the anterior margin as a seam (in posterior view, the sulcus divides the tergum giving it the appearance of two valves) (Figure 17B) … O. plumbago
- -
- Basoflagellomere less than 1.3× longer than wide, usually almost as long as wide. Only females: posterior margin of tergum IV flat, without elevations; posterior margin of tergum V simple, arched in posterior view … 6
- Basoflagellomere oval, 1.1–1.2× longer than wide; scutum hairs erect; sternum II with erect hairs … O. brevicornis
- -
- Basoflagellomere round, as long and wide or slightly wider than long; scutum hairs recumbent; sternum-II hairs semi-recumbent … O. tristis
- Orthonevra arcana sp. n.
- Orthonevra brevicornis (Loew, 1843)
- Orthonevra frontalis (Loew, 1843)
- Orthonevra nobilis (Fallén, 1817)
- Orthonevra tristis (Loew, 1781)
3.5.5. Riponnensia Maibach, Goeldlin de Tiefenau and Speight, 1994a
- -
- Basoflagellomere elongated, over four times longer than wide; antenna black or blackish brown; surstylus of the male genitalia with a small triangular process near the base [6] (figure 18d) … R. longicornis
- -
- Basoflagellomere oval, always much less than four times longer than wide; antenna partly orange; surstylus of the male genitalia without such a process [6] (figure 18a) … R. splendens
- Riponnensia longicornis (Loew, 1843)
- Riponnensia splendens (Meigen, 1822)
4. Molecular Study
5. Discussion
6. Conclusions
- (1)
- A total of 18 species of Chrysogaster, Lejogaster, Melanogaster, Orthonevra and Riponnensia (Brachyopini) are present in the Iberian–Balearic Region. Chrysogaster coerulea is reinstated as a valid species and M. aerosa is removed from the Iberian checklist. A lectotype is designated for the non-Iberian species Orthonevra plumbago.
- (2)
- Two new Spanish endemic species, M. baetica sp. n. and O. arcana sp. n., are described and illustrated.
- (3)
- From the molecular point of view, high levels of interspecific variability in COI-5′ were revealed for Chrysogaster and Orthonevra, but low levels for Lejogaster and Melanogaster.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hull, F.M. The Morphology and Inter-relationships of the Genera of Syrphid Flies, Recent and Fossil. Trans. Zool. Soc. Lond. 1949, 26, 257–408. [Google Scholar] [CrossRef]
- Thompson, F.C. A contribution to a generic revision of the Neotropical Milesinae (Diptera: Syrphidae). Arq. Zool. São Paulo 1972, 23, 73–215. [Google Scholar]
- Thompson, F.C. Notes on the status and relationships of some genera in the tribe Milesiini (Diptera: Syrphidae). Proc. Entomol. Soc. Wash. 1975, 77, 291–305. [Google Scholar]
- Thompson, F.C.; Rotheray, G. Family Syrphidae. In Contributions to a Manual of Palaearctic Diptera; Papp, L., Darvas, B., Eds.; Science Herald: Budapest, Hungary, 1998; Volume 3, pp. 81–139. [Google Scholar]
- Peck, L.V. Syrphidae. In Catalogue of Palaearctic Diptera; Soos, A., Papp, L., Eds.; Akademia Kiado: Budapest, Hungary, 1988; pp. 1–327. [Google Scholar]
- Maibach, A.; Goeldlin de Tiefenau, P.; Speight, M.C.D. Limites génériques et caractéristiques taxonomiques de plusieurs genres de la Tribu des Chrysogasterini (Diptera: Syrphidae). I. Diagnoses génériques et description de Riponnensia Gen. Nov. Ann. Soc. Entomol. Fr. (N.S.) 1994, 30, 217–247. [Google Scholar]
- Maibach, A.; Goeldlin de Tiefenau, P.; Speight, M.C.D. Limites génériques et caractéristiques taxonomiques de plusieurs genres de la Tribu des Chrysogasterini (Diptera: Syrphidae). II. Statut taxonomique de plusieurs des espèces êtudiées et analyse du complexe Melanogaster macquarti (Loew). Ann. Soc. Entomol. Fr. (N.S.) 1994, 30, 253–271. [Google Scholar]
- Kassebeer, C.F. Zur Kenntnis einiger Gattungen der Chrysogasterini in Marokko (Diptera: Syrphidae). Beiträge zur Schwebfliegenfauna Marokkos, IV. Entomol. Z. 1999, 109, 155–164. [Google Scholar]
- Vujić, A. The tribe Chrysogasterini (Diptera: Syrphidae) in the Balkan Peninsula, with the description of three new cryptic species. Studia Dipterol. 1999, 6, 405–423. [Google Scholar]
- Kassebeer, C.F. Eine neue Gattung der Brachyopini (Diptera, Syrphidae) aus dem Mittleren Atlas. Beiträge zur Schwebfliegenfauna Marokkos 8. Dipteron 1999, 2, 11–24. [Google Scholar]
- Moran, K.M.; Skevington, J.H.; Kelso, S.; Mengual, X.; Jordaens, K.; Young, A.D.; Ståhls, G.; Mutin, V.; Bot, S.; van Zuijen, M.; et al. A multigene phylogeny of the eristaline flower flies (Diptera: Syrphidae), with emphasis on the subtribe Criorhinina. Zool. J. Linn. Soc. 2022, 194, 120–135. [Google Scholar] [CrossRef]
- van Eck, A. A checklist of the hoverflies of the Portugal (Diptera, Syrphidae). Boletín Soc. Entomológica Aragonesa 2011, 49, 127–144. [Google Scholar]
- van Eck, A. Hoverflies (Diptera, Syrphidae) new to the fauna of mainland Portugal, with updated hoverfly checklist. Boletín Soc. Entomológica Aragonesa 2016, 59, 187–203. [Google Scholar]
- Ricarte, A.; Marcos-García, M.A. A checklist of the Syrphidae (Diptera) of Spain, Andorra and Gibraltar. Zootaxa 2017, 4216, 401–440. [Google Scholar] [CrossRef]
- Maibach, A.; Goeldlin de Tiefenau, P. Chrysogaster rondanii sp. n. From Western and Central Europe (Diptera: Syrphidae). Bull. Soc. Entomol. Suisse 1995, 68, 459–464. [Google Scholar]
- Gil-Collado, J. Monografía de los Sírfidos de España. Trab. Mus. Nac. Cienc. Nat. 1930, 54, 1–376. [Google Scholar]
- Ricarte, A.; Nedeljković, Z.; Rotheray, G.E.; Lyszkowski, R.M.; Hancock, E.G.; Watt, K.; Hewitt, S.M.; Horsfield, D.; Wilkinson, G. Syrphidae (Diptera) from the Greek island of Lesvos, with description of two new species. Zootaxa 2012, 3175, 1–23. [Google Scholar] [CrossRef]
- Thompson, F.C. A key to the genera of the flower flies (Diptera: Syrphidae) of the Neotropical Region including redescriptions of new genera and species and a glossary of taxonomic terms. Contrib. Entomol. Int. 1999, 3, 320–378. [Google Scholar]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar]
- Ståhls, G.; Barkalov, A. Taxonomic review of the Palaearctic species of the Cheilosia caerulescens-group (Diptera, Syprhidae). ZooKeys 2017, 662, 137–171. [Google Scholar] [CrossRef] [Green Version]
- Ratnasingham, S.; Hebert, P.D.N. BOLD: The Barcode of Life Data System (http://www.barcodinglife.org). Mol. Ecol. Notes 2007, 7, 355–364. [Google Scholar] [CrossRef] [Green Version]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017, 20, 1160–1166. [Google Scholar] [CrossRef] [Green Version]
- Larsson, A. AliView: A fast and lightweight alignment viewer and editor for large datasets. Bioinformatics 2014, 30, 3276–3278. [Google Scholar] [CrossRef] [PubMed]
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tamura, K.; Nei, M.; Kumar, S. Prospects for inferring very large phylogenies by using the neighbour-joining method. Proc. Natl. Acad. Sci. USA 2004, 101, 11030–11035. [Google Scholar] [CrossRef] [Green Version]
- Nei, T.; Kumar, S. Molecular Evolution and Phylogenetics; Oxford University Press: New York, NY, USA, 2000; p. 348. [Google Scholar]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pérez-Bañón, C.; Radenković, S.; Vujić, A.; Ståhls, G.; Rojo, S.; Grković, A.; Petanidou, T. Brachyopa minima (Diptera: Syrphidae), a new species from Greece with notes on the biodiversity and conservation of the genus Brachyopa Meigen in the Northern Aegean Islands. Zootaxa 2016, 4072, 217–234. [Google Scholar] [CrossRef] [PubMed]
- van Steenis, J.; van Zuijen, M.P.; van Steenis, W.; Makris, C.; van Eck, A.; Mengual, X. Hoverflies (Diptera: Syrphidae) of Cyprus: Results from a collecting trip in October 2017. Bonn Zool. Bull. 2019, 68, 125–146. [Google Scholar] [CrossRef]
- Roslin, T.; Somervuo, P.; Pentinsaari, M.; Hebert, P.D.N.; Agda, J.; Ahlroth, P.; Anttonen, P.; Aspi, J.; Blagoev, G.; Blanco, S.; et al. A molecular-based identification resource for the arthropods of Finland. Mol. Ecol. Resour. 2021, 22, 803–822. [Google Scholar] [CrossRef]
- Mengual, X.; Ståhls, G.; Rojo, S. Phylogenetic relationships and taxonomic ranking of pipizine flower flies (Diptera: Syrphidae) with implications for the evolution of aphidophagy. Cladistics 2015, 31, 491–508. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Ratnasingham, S.; Zakharov, E.V.; Telfer, A.C.; Levesque-Beaudin, V.; Milton, M.A.; Pedersen, S.; Jannetta, P.; de Waard, J.R. Counting animal species with DNA barcodes: Canadian insects. Proc. R. Soc. Lond. Ser. B 2016, 371, 20150333. [Google Scholar] [CrossRef] [Green Version]
- Geiger, M.F.; Moriniere, J.; Hausmann, A.; Haszprunar, G.; Wägele, W.; Hebert, P.D.N.; Rulik, B. Testing the Global Malaise Trap Program–How well does the current barcode reference library identify flying insects in Germany? Biodivers. Data J. 2016, 4, e10671. [Google Scholar] [CrossRef] [Green Version]
- Ricarte, A. Biodiversidad de Sírfidos (Diptera: Syrphidae) y Conservación de los Hábitats en el Parque Nacional de Cabañeros, España. Ph.D. Thesis, Universidad de Alicante, Alicante, Spain, 2008. [Google Scholar]
- Czerny, L.; Strobl, P.G. Spanische Dipteren. III. Verh. Der Zool. Bot. Ges. Wien 1909, 59, 121–301. [Google Scholar]
- Chvála, M. The Types of Diptera (Insecta) described by Pater Gabriel Strobl. Studia Dipterol. Suppl. 2008, 17, 1–281. [Google Scholar]
- Systema Dipterorum, Version 2.10. Available online: http://diptera.org/ (accessed on 23 March 2021).
- Marcos-García, M.A. Catálogo preliminar de los Syrphidae (Diptera) de la Cordillera Cantábrica (España). Eos 1990, 66, 83–100. [Google Scholar]
- Marcos-García, M.A. Los Syrphidae (Diptera) de las sierras occidentales del Sistema Central español: Subfamilias: Chrysotoxinae, Sphegininae, Chilosiinae, Pelecocerinae, Volucellinae y Cinxiinae. Boletín Asoc. Española Entomol. 1986, 10, 159–180. [Google Scholar]
- Marcos-García, M.A. Nuevas citas para la fauna ibérica de Sírfidos (Diptera). Misc. Zool. 1986, 10, 205–211. [Google Scholar]
- Stackelberg, A.A. The Palearctic species of the genus Chrysogaster Mg. (Diptera, Syrphidae). Entomol. Rev. 1959, 38, 806–811. [Google Scholar]
- Stackelberg, A.A. Palearkticheskie vidy roda Orthoneura Macq. (Diptera, Syrphidae). Entomol. Obozr. 1953, 33, 342–357. (In Russian) [Google Scholar]
- van Veen, M.P. Hoverflies of Northwest Europe: Identification Keys to the Syrphidae; KNNV Publishing: Utrecht, The Netherlands, 2004; p. 256. [Google Scholar]
- Bartsch, H.; Binkiewicz, E.; Klintbjer, A.; Rådén, A.; Nasibov, E. Blomflugor: Eristalinae and Microdontinae. In Nationalnyckeln Till Sveriges Flora Och Flora, DH 53b; Artdatabanken; SLU: Uppsala, Sweden, 2009; p. 478. [Google Scholar]
- Speight, M.C.D.; Castella, E. Dix-neuf additions à la liste des Syrphidae (Diptera) de Haute-Savoie, incluant Orthonevra plumbago (Loew, 1840) et Xanthogramma stackelbergi Violovitsh, 1975, deux espèces nouvelles pour la France. Entomo Helv. 2011, 4, 45–58. [Google Scholar]
- Haarto, A.; Kerppola, S. Finnish Hoverflies and Some Species in Adjacent Countries; Otavan Kirjapaino Oy: Keuruu, Finland, 2007; p. 647. [Google Scholar]
- Stackelberg, A.A. New data on the taxonomy of Palaearctic hover flies (Diptera, Syrphidae). Entomol. Obozr. 1965, 44, 907–926, (In Russian; republished in English in Entomological Society of America. Scripta Technology, Washington 4, 528–537). [Google Scholar]
- Marcos-García, M.A. Contribución al conocimiento de la sirfidofauna del Pirineo del Alto Aragón. I (Diptera, Syrphidae). In Actas do II Congresso Ibérico de Entomologia; de Carvalho, L., Ed.; Boletim da Sociedade Portuguesa de Entomologia: Lisboa, Portugal, 1985; pp. 511–520. [Google Scholar]
- Speight, M.C.D. Species accounts of European Syrphidae, 2018. In Syrph the Net, the Database of European Syrphidae (Diptera); Syrph the Net publications: Dublin, Ireland, 2018; Volume 103, p. 302. [Google Scholar]
- van der Goot, V.S. Quelques Syrphides (Diptera) des Pyrénées et de la Sierra Nevada. Entomol. Ber. 1958, 18, 93–96. [Google Scholar]
- Carles-Tolrá, M. Sírfidos nuevos para Andorra y la Península Ibérica (Diptera: Syrphidae). Heteropterus Rev. Entomol. 2006, 6, 145–156. [Google Scholar]
- Andréu, J. Notas dipterológicas. Una lista de Sírfidos para contribuir al conocimiento de los dípteros de España. Boletín Soc. Entomol. Esp. 1926, 9, 98–126. [Google Scholar]
- Ricarte, A.; Nedeljković, Z.; Marcos-García, M.A. An exploratory survey and assessment of the hoverfly diversity (Diptera: Syrphidae) from the Pyrenees of Girona, Spain. Rev. Suisse Zool. 2021, 128, 381–398. [Google Scholar] [CrossRef]
- Leclercq, M. Syrphidae (Diptera) des Pyrénées (Huesca, Lérida, Andorra, Gerona). Pirineos 1971, 102, 21–25. [Google Scholar]
- Kehlmaier, C. Hoverflies (Diptera, Syrphidae) from northern Spain, with notes on Pelecocera tricincta Meigen, 1822. Volucella 2002, 6, 139–153. [Google Scholar]
- Carles-Tolrá, M.; Verdugo, A. Algunos dípteros capturados en el Parque Natural de los Alcornocales (España, Cádiz) mediante mangueo y trampas (Insecta, Diptera). Boletín Soc. Entomológica Aragonesa 2009, 44, 139–142. [Google Scholar]
- van Eck, A. Poorly recorded Syrphidae (Diptera) from Spain including species new to its fauna. Boletín Soc. Entomológica Aragonesa 2010, 46, 299–300. [Google Scholar]
- Speight, M.C.D. Species accounts of European Syrphidae, 2020. In Syrph the Net, the Database of European Syrphidae (Diptera); Syrph the Net publications: Dublin, Ireland, 2020; Volume 104, p. 314. [Google Scholar]
- De-la-Fuente-Morales, J.M. Dípteros de la provincia de Ciudad Real. Boletín de la Soc. Entomol. Esp. 1926, 9, 130–133. [Google Scholar]
- Kanervo, E. Inventa entomologica itineris Hispanici et Maroccani, quod a. 1926 fecerunt Harald et Hakan Lindberg. XXIV Diptera. Syrphidae. Soc. Sci. Fenn. Comment. Biol. 1939, 7, 1–6. [Google Scholar]
- van der Goot, V.S.; Lucas, J.A.W. Recolección de sírfidos en Albarracín, provincia de Teruel durante el verano de 1965. Graellsia 1967, 23, 111–119. [Google Scholar]
- Pedersen, E.T. Some Syrphidae from Spain, with descriptions of two new species (Insecta, Diptera). Steenstrupia 1971, 21, 229–245. [Google Scholar]
- Carles-Tolrá, M. Algunos dípteros capturados en el País Vasco y Navarra (España) (Insecta, Diptera). Heteropterus Rev. Entomol. 2011, 11, 35–48. [Google Scholar]
- Carles-Tolrá, M.; Camaño-Portela, J.L.; Pino-Pérez, J.J.; Pino-Pérez, R. 212 especies de dípteros de Galicia (NO España) (Insecta: Diptera). Boletín BIGA 2010, 9, 89–108. [Google Scholar]
- Röder, V. Ueber von Herr Dr. Schmiedeknecht in Spanien, bei Elche, Ibiza und auf Mallorca gesammelte Dipteren. Entomol. Nachr. 1884, 10, 253–257. [Google Scholar]
- Kassebeer, C.F. Zur Schwebfliegenfauna (Diptera, Syrphidae) der Balearen. Dipteron 2002, 4, 131–156. [Google Scholar]
- Gomes, A. Sirfideos colhidos em Portugal durante 1977 (Diptera, Syrphidae). Agron. Lusit. 1981, 41, 5–24. [Google Scholar]
- Isidro, P.M. Biología y Ecología de los Sírfidos (Diptera, Syrphidae) del Parque Natural del Carrascal de la Font Roja (Alicante, España). Graduate Thesis, Universidad de Alicante, Alicante, Spain, 1995. [Google Scholar]
- Lucas, J.A.W. Notes on syrphid flies of Portugal (Diptera, Syrphidae). Agron. Lusit. 1996, 45, 317–326. [Google Scholar]
- Pérez-Bañón, C. Estudio de la Comunidad de Sírfidos de las Sierras Valencianas del Negrete y Utiel (Diptera, Syrphidae). Graduate Thesis, Universidad de Alicante, Alicante, Spain, 1995. [Google Scholar]
- Vujić, A.; Stuke, J.-H. A new hoverfly species of the genus Melanogaster from Central Europe (Diptera, Syrphidae). Studia Dipterol. 1998, 5, 343–347. [Google Scholar]
- Marcos-García, M.A. Contribución al conocimiento de los Syrphidae (Dipt.) de la zona noreste de la provincia de Salamanca. Boletín Asoc. Española Entomol. 1981, 4, 157–171. [Google Scholar]
- Anadón, A. La colección de sírfidos del departamento de Zoología de la Universidad de Oviedo (Diptera: Syrphidae). Boletín Cienc. Nat. 1984, 33, 121–138. [Google Scholar]
- van Steenis, W.; van Steenis, J.; van der Ent, L.-J. Chrysogaster rondanii. IUCN Red List. Threat. Species 2021, e.T149165436A149165446. [Google Scholar] [CrossRef]
- Nedeljković, Z.; Ačanski, J.; Vujić, A.; Obreht, D.; Đan, M.; Ståhls, G.; Radenković, S. Taxonomy of Chrysotoxum festivum Linnaeus, 1758 (Diptera: Syrphidae)—An integrative approach. Zool. J. Linn. Soc. 2013, 169, 84–102. [Google Scholar] [CrossRef] [Green Version]
- Marcos-García, M.A.; Vujić, A.; Mengual, X. Revision of Iberian species of the genus Merodon (Diptera: Syrphidae). Eur. J. Entomol. 2007, 104, 531–572. [Google Scholar] [CrossRef] [Green Version]
- Hebert, P.D.N.; Ratnasingham, S.; de Waard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. Lond. Ser. B 2003, 270, S96–S99. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wiemers, M.; Fiedler, K. Does the DNA barcoding gap exist?—A case study in blue butterflies (Lepidoptera: Lycaenidae). Front. Zool. 2007, 4, 8. [Google Scholar] [CrossRef] [Green Version]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; de Waard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. Ser. B 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rotheray, G.E. Colour guide to hoverfly larvae (Diptera, Syrphidae) in Britain and Europe. Dipter. Dig. 1993, 9, 1–156. [Google Scholar]
- van Steenis, W.; van Steenis, J.; van der Ent, L.-J. Chrysogaster basalis. IUCN Red List. Threat. Species 2021, e.T149165454A149165458. [Google Scholar] [CrossRef]
- van Steenis, W.; van Steenis, J.; van der Ent, L.-J. Riponnensia longicornis. IUCN Red List. Threat. Species 2021, e.T149169180A149169185. [Google Scholar] [CrossRef]



















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Life Stage | Country of Origin | DNA Voucher | Acc. Number (Genbank) | Specimen ID (BOLD) | Reference |
|---|---|---|---|---|---|---|
| Brachyopa minimaVujić and Pérez-Bañón in Pérez-Bañón et al. (2016) | Larva | Greece | MZH_Y543 | KU315385 | GBDP31699-19 | [28] |
| Ceriana vespiformis (Latreille, 1809) | Adult | Spain | Jeff_Skevington_Specimen25788 | MK972432 | SYCNC169-16 | [29] |
| Chrysogaster coemiteriorum (Linnaeus, 1758) | Adult (♂) | Finland | KWi-325 | MZ625176 | DIFIA390-12 | [30] |
| C. coemiteriorum (Linnaeus, 1758) | Adult (♀) | Norway | NORSY472 | - | NORSY472-15 | [21] |
| C. coemiteriorum (Linnaeus, 1758) | Adult (♀) | Norway | NORSY473 | - | NORSY473-15 | [21] |
| C. coeruleaStrobl in Czerny and Strobl, 1909 | Adult (♀) | Spain | CEUA_S109 | ON103531 | - | Present work |
| C. solstitialis(Fallén, 1817) | Adult (♀) | Norway | NORSY474 | - | NORSY474-15 | [21] |
| C. solstitialis(Fallén, 1817) | - | Finland | MZH_DNA_voucher_Y583_2007 | EU431480 | - | [31] |
| C. solstitialis(Fallén, 1817) | Adult (♂) | Finland | jka11-00486 | MZ624811 | FIDIP2612-12 | [30] |
| C. solstitialis(Fallén, 1817) | Adult (♂) | Spain | CEUA_S74 | ON103525 | - | Present work |
| C. solstitialis(Fallén, 1817) | Adult (♂) | Spain | CEUA_S75 | ON103526 | - | Present work |
| C. solstitialis(Fallén, 1817) | Adult (♀) | Spain | CEUA_S76 | ON103527 | - | Present work |
| C. solstitialis(Fallén, 1817) | Adult (♀) | Spain | CEUA_S103 | ON103529 | - | Present work |
| C. solstitialis(Fallén, 1817) | Adult (♂) | Spain | CEUA_S106 | ON103530 | - | Present work |
| Lejogaster metallina(Fabricius, 1777) | Adult (♂) | Norway | NorSy92 | - | NORSY092-12 | [21] |
| L. metallina(Fabricius, 1777) | Adult (♂) | United Kingdom | BMNH(E)#1731751 | - | BEEEE381-16 | [21] |
| L. tarsata(Meigen, 1822) | Adult (♀) | Finland | jka07-01903 | MZ628003 | FIDIP2534-12 | [30] |
| Melanogaster aerosa(Loew, 1843) | Adult (♂) | Finland | MZH_HP.691 | MZ631040 | FIDIP4127-14 | [30] |
| M. aerosa(Loew, 1843) | Adult (♀) | Finland | MZH_HP.749 | MZ609615 | FIDIP955-12 | [30] |
| M. aerosa(Loew, 1843) | Adult (♂) | Norway | NorSy331 | - | NORSY331-12 | [21] |
| M. baeticaRicarte and Nedeljković sp. n. | Adult (♂) | Spain | CEUA_S73 | ON103524 | - | Present work |
| M. hirtella(Loew, 1843) | Adult (♀) | United Kingdom | BMNH(E)#1731754 | - | BEEEE384-16 | [21] |
| M. hirtella(Loew, 1843) | Adult (♂) | United Kingdom | BMNH(E)#1731756 | - | BEEEE386-16 | [21] |
| M. parumplicata(Loew, 1840) | Adult (♀) | Norway | NORSY483 | - | NORSY483-15 | [21] |
| Orthonevra anniae(Sedman, 1966) | Adult (♀) | Canada | BIOUG<CAN>:10JSROW-0602 | JF873508 | JSDIP602-10 | [32] |
| O. anniae(Sedman, 1966) | Adult (♂) | Canada | BIOUG73902-G04 | - | BCBEE171-21 | [21] |
| O. arcanaRicarte and Nedeljković sp. n. | Adult (♀) | Spain | CEUA_S59 | ON103520 | - | Present work |
| O. arcanaRicarte and Nedeljković sp. n. | Adult (♂) | Spain | CEUA_S60 | ON103521 | - | Present work |
| O. arcanaRicarte and Nedeljković sp. n. | Adult (♀) | Spain | CEUA_S61 | ON103522 | - | Present work |
| O. erythrogona(Malm, 1863) | Adult (♂) | Finland | MZH_HP.1138 | MZ629570 | FIDIP3700-13 | [30] |
| O. erythrogona(Malm, 1863) | Adult (♀) | Finland | MZH_HP.1139 | MZ630425 | FIDIP3701-13 | [30] |
| O. frontalis(Loew, 1843) | Adult (♂) | Spain | CEUA_S63 | ON103523 | - | Present work |
| O. geniculata(Meigen, 1830) | Adult (♀) | Finland | KWi-095 | MZ627126 | DIFIA539-12 | [30] |
| O. geniculata(Meigen, 1830) | Adult (♂) | Norway | NorSy311 | - | NORSY311-12 | [21] |
| O. intermedia(Lundbeck, 1916) | Adult (♂) | Finland | MZH_HP.37 | MZ631691 | FIDIP037-11 | [30] |
| O. intermedia(Lundbeck, 1916) | Adult (♀) | Finland | MZH_HP.756 | MZ609029 | FIDIP962-12 | [30] |
| O. nitida(Wiedemann, 1830) | Adult (♀) | Canada | BIOUG<CAN>:PCPP10-0132 | JF867625 | BBDCN037-10 | [32] |
| O. nitida(Wiedemann, 1830) | Adult (♂) | United States | BIOUG<CAN>:09BBDIP-0534 | GU803796 | USDIP534-09 | [21] |
| O. nobilis(Fallén, 1817) | Adult (♀) | Finland | MZH_HP.33 | MZ610319 | FIDIP033-11 | [30] |
| O. nobilis(Fallén, 1817) | - | Germany | BIOUG17224-E02 | - | GMGMK1311-14 | [33] |
| O. nobilis(Fallén, 1817) | Adult (♂) | Norway | NorSy85 | - | NORSY085-12 | [21] |
| O. nobilis(Fallén, 1817) | Adult (♂) | Spain | CEUA_S77 | ON103528 | - | Present work |
| O. pictipennis(Loew, 1863) | Adult (♀) | Canada | BIOUG<CAN>:10BBCDIP-1237 | JF867314 | BBDCM522-10 | [32] |
| O. pulchella(Williston, 1887) | Adult (♂) | Canada | BIOG<CAN>:09BBEDI-0456 | HM435787 | BBDEC456-09 | [32] |
| O. pulchella(Williston, 1887) | Adult (♀) | Canada | BIOUG<CAN>:09BBEDI-0480 | HM435810 | BBDEC480-09 | [32] |
| O. robusta(Shannon, 1916) | Adult (♂) | Canada | BIOUG73902-G01 | - | BCBEE168-21 | [21] |
| O. stackelbergiThompson and Torp, 1982 | Adult (♀) | Norway | NorSy335 | - | NORSY335-12 | [21] |
| O. stackelbergiThompson and Torp, 1982 | Adult (♂) | Norway | NorSy337 | - | NORSY337-12 | [21] |
| O. tristis(Loew, 1871) | Adult (♀) | Germany | BC-ZSM-DIP-24053-C01 | - | AMTPE1070-15 | [21] |
| O. weemsi(Sedman, 1966) | Adult (♂) | United States | BIOUG<CAN>:10BBDIP-0834 | JF872322 | DIPUS834-10 | [21] |
| O. weemsi(Sedman, 1966) | Adult (♀) | Canada | BIOUG17325-E11 | KR978766 | SSKJC2166-15 | [32] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ricarte, A.; Nedeljković, Z.; Aguado-Aranda, P.; Marcos-García, M.Á. Assessing the Diversity and Systematics of Brachyopini Hoverflies (Diptera: Syrphidae) in the Iberian Peninsula, Including the Descriptions of Two New Species. Insects 2022, 13, 648. https://doi.org/10.3390/insects13070648
Ricarte A, Nedeljković Z, Aguado-Aranda P, Marcos-García MÁ. Assessing the Diversity and Systematics of Brachyopini Hoverflies (Diptera: Syrphidae) in the Iberian Peninsula, Including the Descriptions of Two New Species. Insects. 2022; 13(7):648. https://doi.org/10.3390/insects13070648
Chicago/Turabian StyleRicarte, Antonio, Zorica Nedeljković, Pablo Aguado-Aranda, and Mª Ángeles Marcos-García. 2022. "Assessing the Diversity and Systematics of Brachyopini Hoverflies (Diptera: Syrphidae) in the Iberian Peninsula, Including the Descriptions of Two New Species" Insects 13, no. 7: 648. https://doi.org/10.3390/insects13070648