
Ultrastructure of Antennal Sensory Organs in Nine Flesh Flies (Diptera: Sarcophagidae): New Insight into the Definition of Family Sarcophagidae


 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
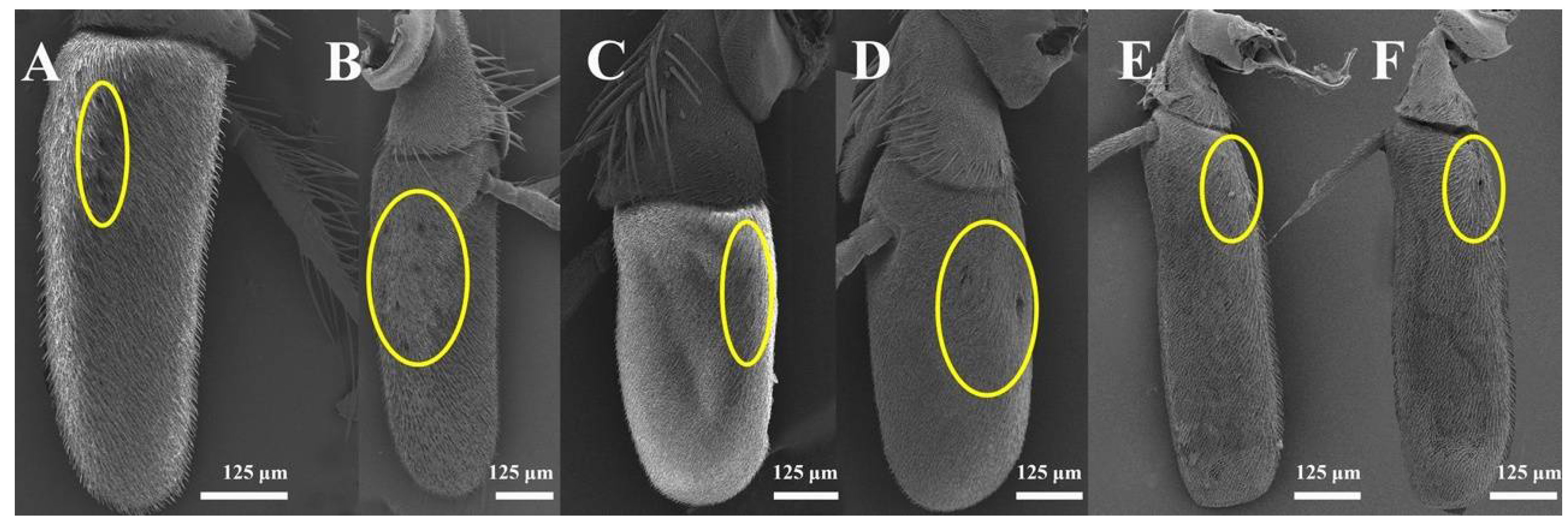
3.1. General Description of the Antennal Morphology of the Nine Species of Sarcophagidae
3.1.1. Scape and Pedicel
3.1.2. Postpedicel
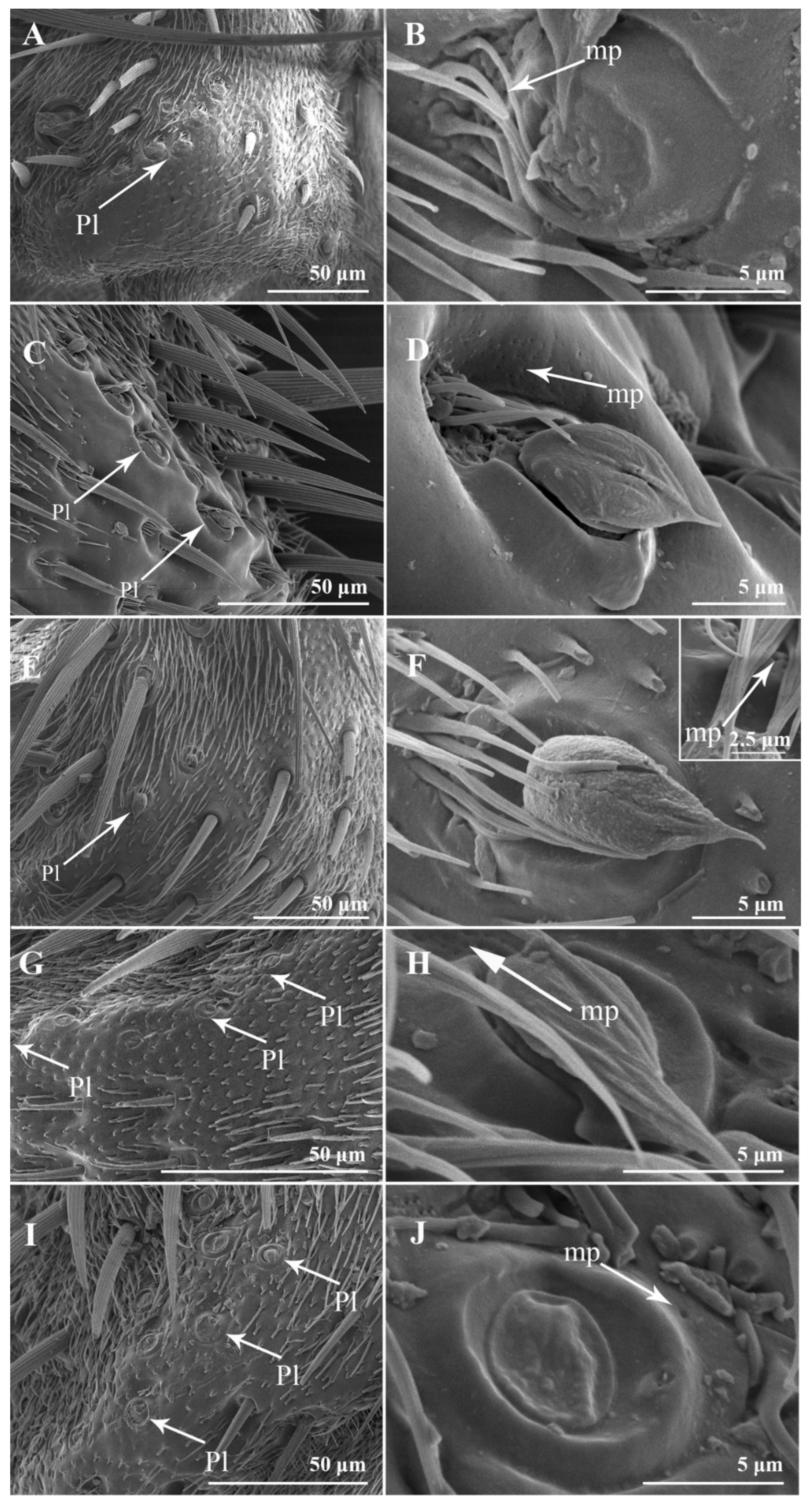
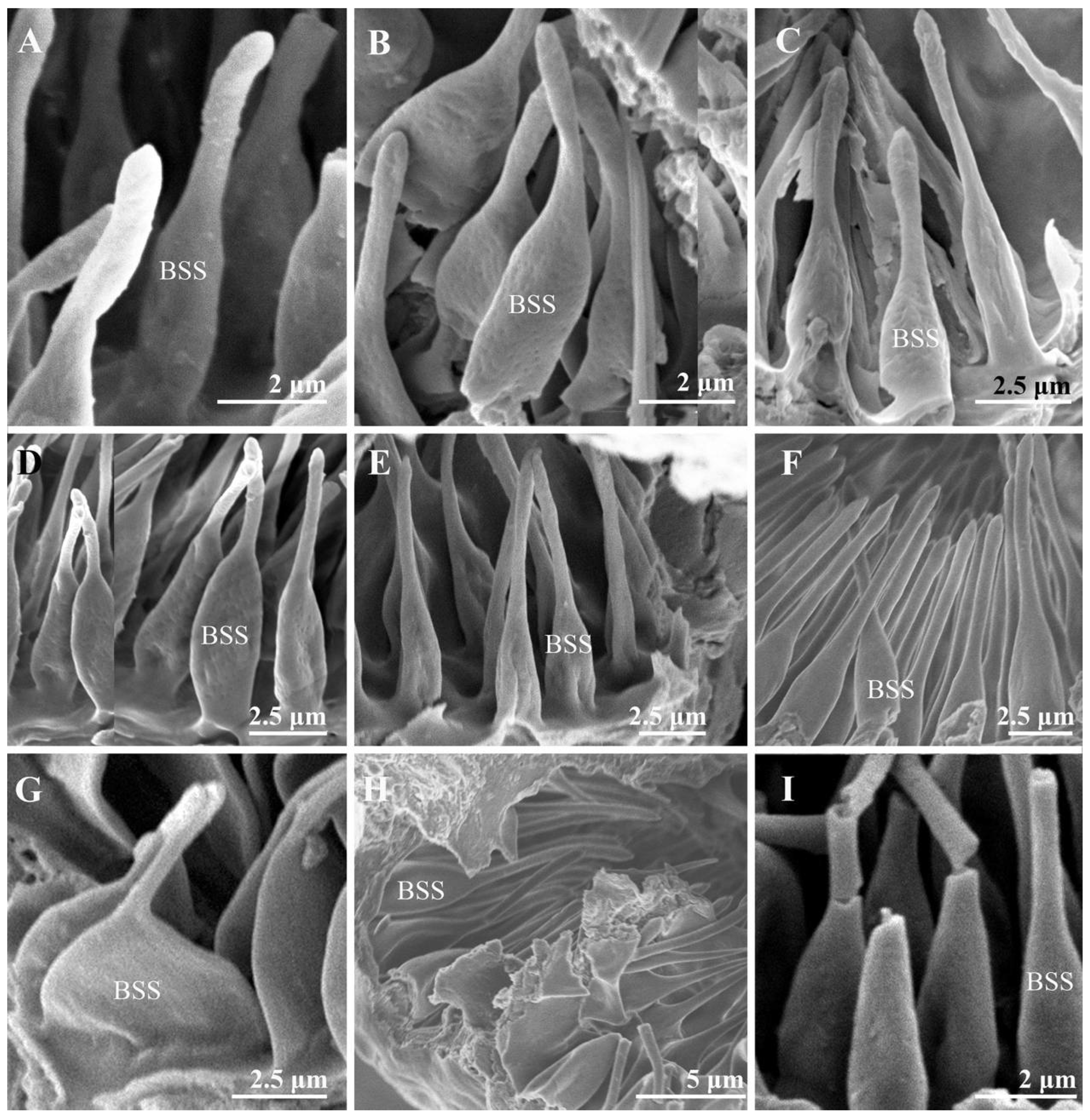
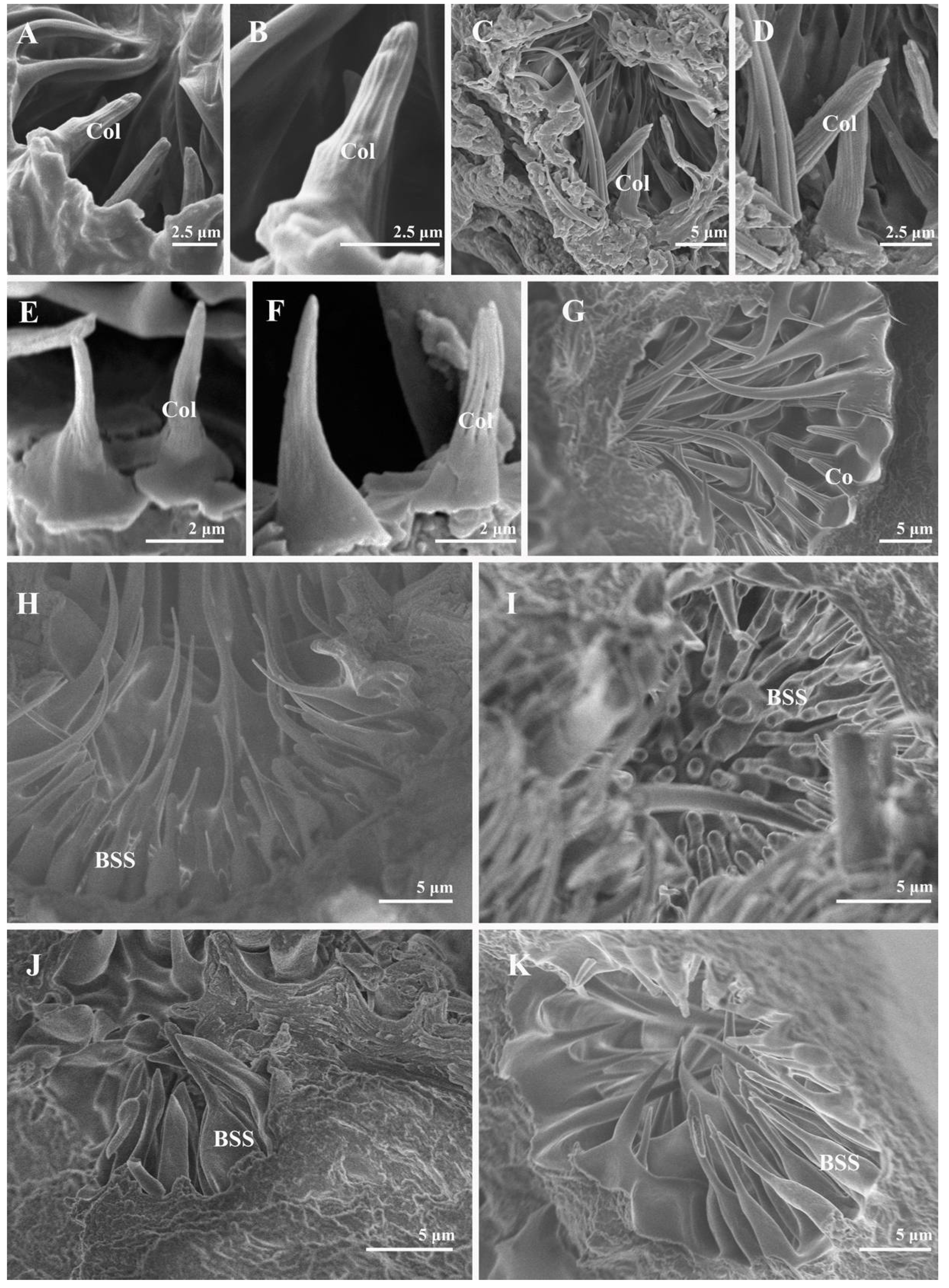
3.2. Sensilla and Sensory Pits on the Postpedicel
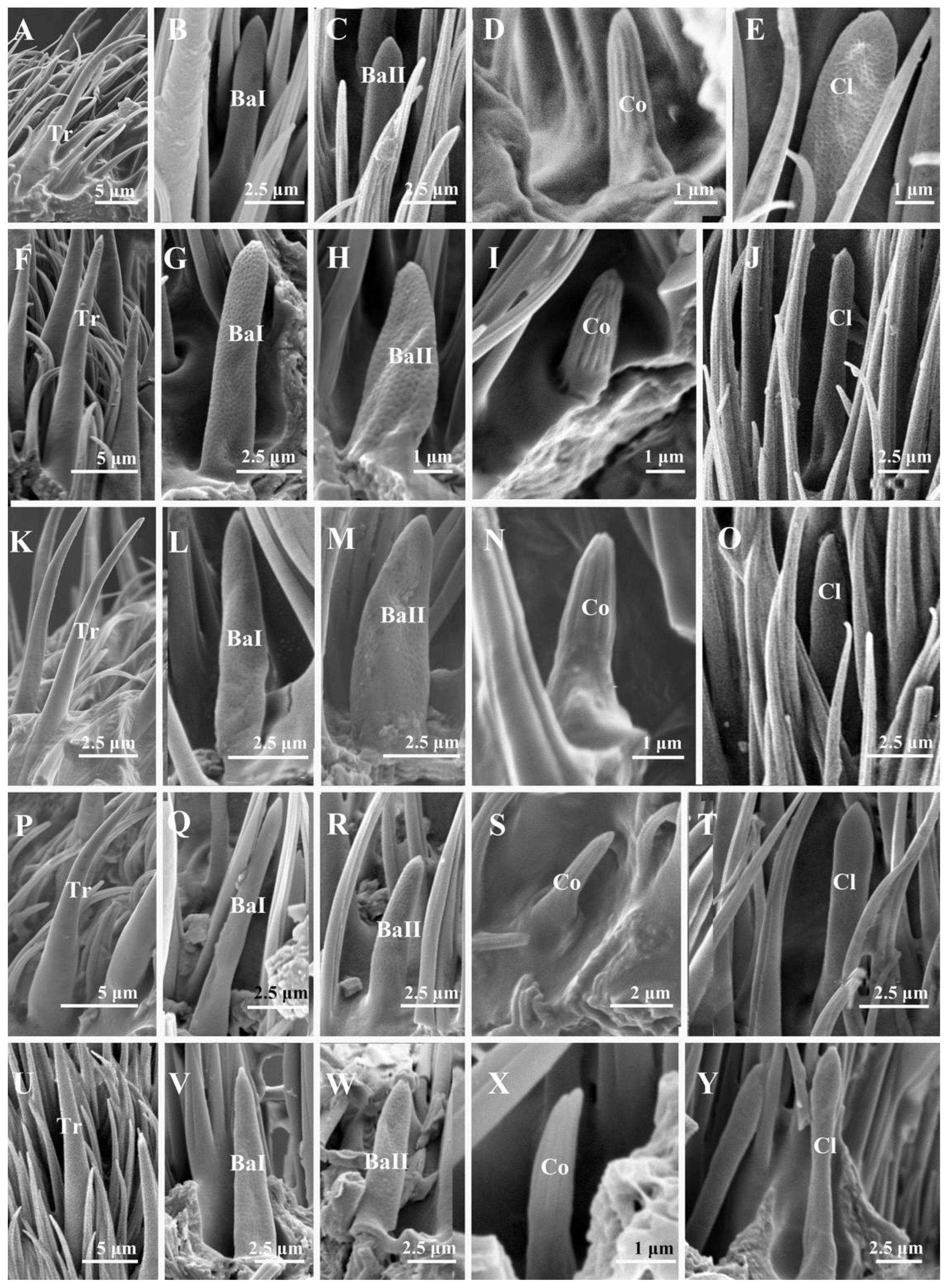
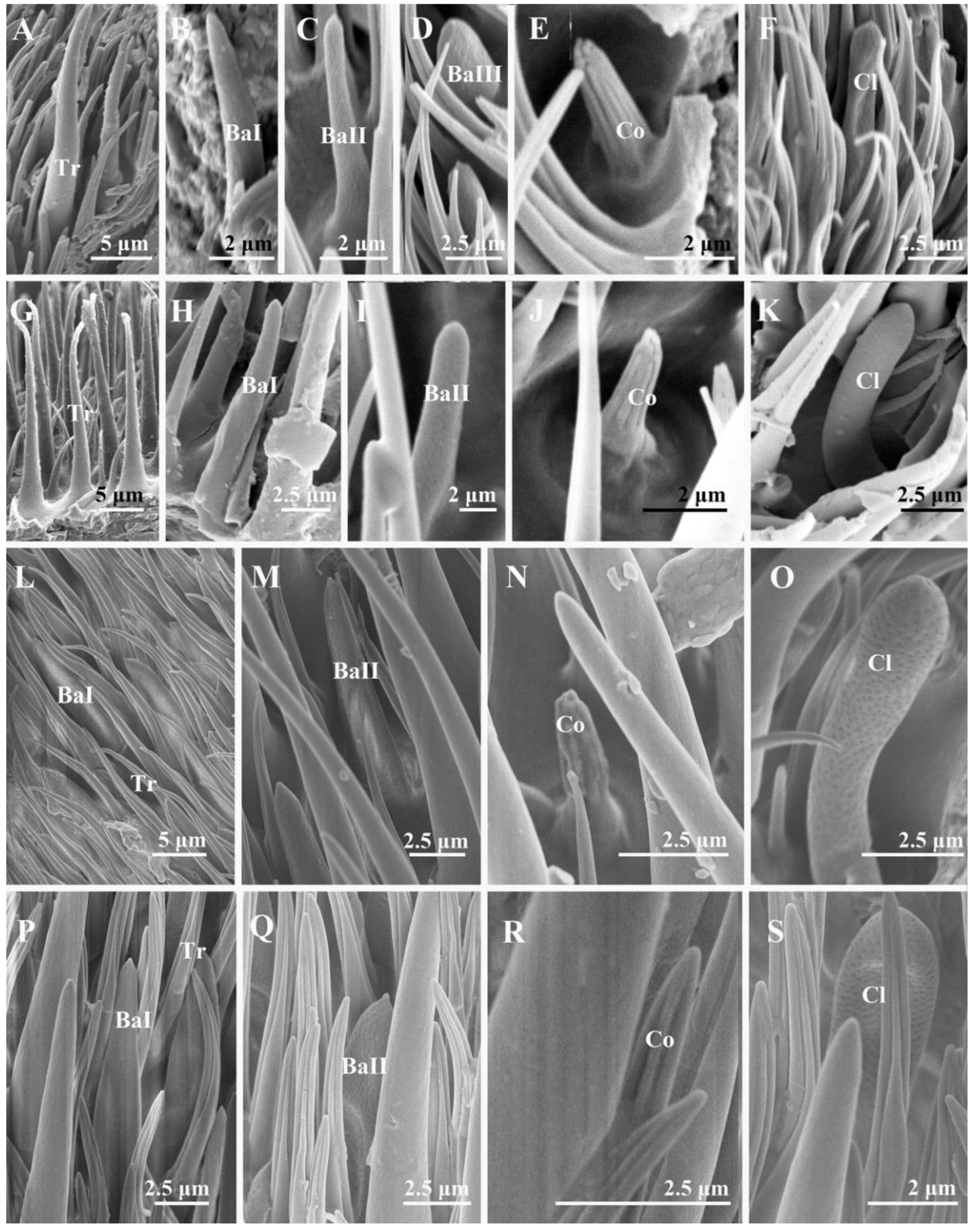
3.2.1. Trichoid Sensilla
3.2.2. Basiconic Sensilla
3.2.3. Coeloconic Sensilla
3.2.4. Clavate Sensilla
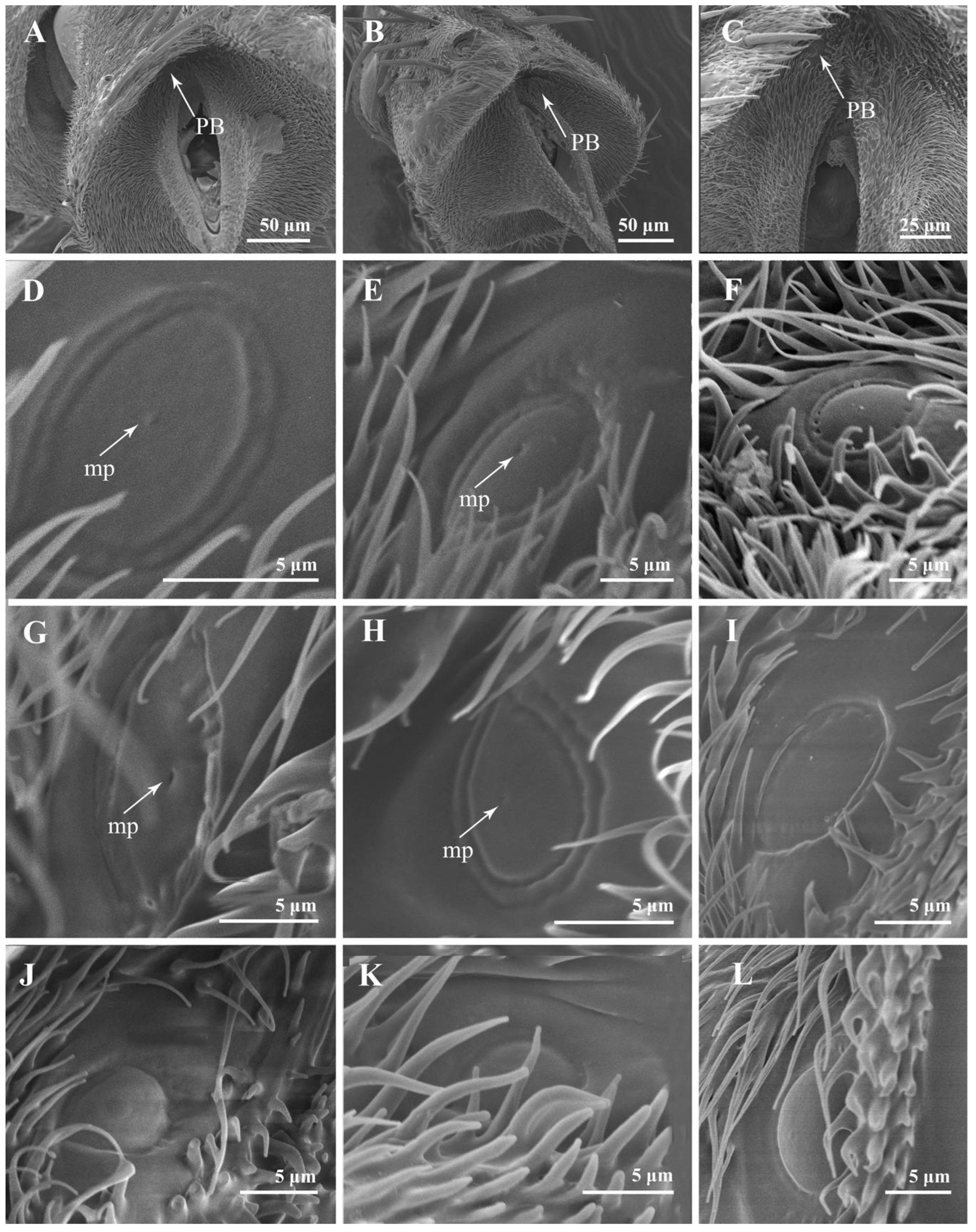
3.2.5. Sensory Pits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pape, T. Catalogue of the Sarcophagidae of the World (Insecta: Diptera); Memoirs of Entomology International: Gainesville, FL, USA, 1996; Volume 8, pp. 1–558. [Google Scholar]
- Pape, T.; Blagoderov, V.; Mostovski, M.B. Order Diptera Linnaeus, 1758. In Animal Biodiversity: An Outline of Higher-Level Classification and Survey of Taxonomic Richness; Zhang, Z.-Q., Ed.; Magnolia Press: Auckland, New Zealand, 2011; Volume 3148, pp. 222–229. [Google Scholar] [CrossRef] [Green Version]
- Piwczyński, M.; Pape, T.; Dejasikora, E.; Sikora, M.; Akbarzadeh, K.; Szpila, K. Molecular phylogeny of Miltogramminae (Diptera: Sarcophagidae): Implications for classification, systematics and evolution of larval feeding strategies. Mol. Phylogenet. Evol. 2017, 116, 49–60. [Google Scholar] [CrossRef] [PubMed]
- Buenaventura, E.; Szpila, K.; Cassel, B.K.; Wiegmann, B.M.; Pape, T. Anchored hybrid enrichment challenges the traditional classification of flesh flies (Diptera: Sarcophagidae). Syst. Entomol. 2020, 45, 281–301. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.P.; Buenaventura, E.; Pape, T.; Kutty, S.N.; Bayless, K.M.; Zhang, D. A phylotranscriptomic framework for flesh fly evolution (Diptera, Calyptratae, Sarcophagidae). Cladistics 2020, 37, 540–558. [Google Scholar] [CrossRef]
- Daniel, M.; Šrámová, H.; Zálabská, E. Lucilia sericata (Diptera: Calliphoridae) causing hospital-acquired myiasis of a traumatic wound. J. Hosp. Infect. 1994, 28, 149–152. [Google Scholar] [CrossRef]
- Zaglool, D.A.; Tayeb, K.; Khodari, Y.A.; Farooq, M.U. First case report of human myiasis with Sarcophaga species in Makkah city in the wound of a diabetic patient. J. Nat. Sci. Biol. Med. 2013, 4, 225–228. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giangaspero, A.; Marangi, M.; Balotta, A.; Venturelli, C.; Szpila, K.; Di Palma, A. Wound Myiasis Caused by Sarcophaga (Liopygia) Argyrostoma (Robineau-Desvoidy) (Diptera: Sarcophagidae): Additional Evidences of the Morphological Identification Dilemma and Molecular Investigation. Sci. World J. 2017, 2017, 9064531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ren, L.P.; Shang, Y.J.; Chen, W.; Meng, F.M.; Cai, J.F.; Zhu, G.H.; Chen, L.S.; Wang, Y.; Deng, J.Q.; Guo, Y.D. A brief review of forensically important flesh flies (Diptera: Sarcophagidae). Forensic. Sci. Res. 2018, 3, 16–26. [Google Scholar] [CrossRef] [Green Version]
- Yan, L.P.; Zhang, M.; Tang, L.P.; Ente, M.; Ma, X.P.; Chu, H.J.; Li, K.; Hu, D.F.; Zhang, D. First reports of nasal and traumatic myiasis infection in endangered Przewalski’s horses (Equus ferus przewalskii). Int. J. Parasitol. Parasites Wildl. 2019, 9, 21–24. [Google Scholar] [CrossRef]
- Spofford, M.G.; Kurczewski, F.E.; Downes, W.L., Jr. Nearctic Species of Miltogrammini (Diptera: Sarcophagidae) Associated with Species of Aculeata (Hymenoptera: Vespoidea, Pompiloidea, Sphecoidea, Apoidea). J. Kans. Entomol. Soc. 1989, 62, 254–267. [Google Scholar]
- Spofford, M.G.; Kurczewski, F.E. Counter-cleptoparasitic behaviours of species of Sphecidae (Hymenoptera) in response to miltogrammini larviposition (Diptera: Sarcophagidae). Ann. Mag. Nat. Hist. 2007, 26, 993–1012. [Google Scholar] [CrossRef]
- Rohdendorf, B.B.; Verves, Y.G. On the fauna of Sarcophagidae (Diptera) of the Mongolian People’s Republic. III. Miltogrammatinae. Insects Mong. 1980, 7, 445–518. (In Russian) [Google Scholar]
- Szpila, K.; Voss, J.G.; Pape, T. A new forensic indicator for buried bodies (Diptera, Sarcophagidae, Miltogramminae). Med. Vet. Entomol. 2010, 24, 278–283. [Google Scholar] [CrossRef] [PubMed]
- Roback, S.S. The evolution and taxonomy of the Sarcophaginae (Diptera: Sarcophagidae). Ill. Biol. Monogr. 1954, 23, 55–56. [Google Scholar]
- Pape, T. The Sarcophagidae (Diptera) of Fennoscandia and Denmark. Fauna Entomol. Scand. 1987, 19, 1–203. [Google Scholar]
- Kuhlmann, U. Biology and predation rate of the Sarcophagid fly, Agria mamillata, a predator of European small ermine moths. Int. J. Pest Manag. 1995, 41, 67–73. [Google Scholar] [CrossRef]
- Zumpt, F. Myiasis in Man and Animals in the Old World. A Textbook for Physicians, Veterinarians and Zoologists; Butter-Worth: London, UK, 1965; pp. 106–110. [Google Scholar]
- Hall, M.J.R.; Wall, R.L.; Stevens, J.R. Traumatic Myiasis: A Neglected Disease in a Changing World. Annu. Rev. Entomol. 2016, 61, 159. [Google Scholar] [CrossRef] [PubMed]
- Buenaventura, E. Museomics and phylogenomics with protein-encoding ultraconserved elements illuminate the evolution of life history and phallic morphology of flesh flies (Diptera: Sarcophagidae). BMC Ecol. Evol. 2021, 21, 70. [Google Scholar] [CrossRef]
- Fernandes, F.D.F.; Freitas, E.D.P.E.S.; Linardi, P.M.; Pimenta, P.F.P. Ultrastructure of contact-chemoreceptor sensilla found among the genae of female Gasterophilus nasalis. J. Parasitol. 2005, 91, 1218–1220. [Google Scholar] [CrossRef]
- Guha, L.; Seenivasagan, T.; Bandyopadhyay, P.; Iqbal, S.T.; Sathe, M.; Sharma, P.; Parashar, B.D.; Kaushik, M.P. Oviposition and flight orientation response of Aedes aegypti to certain aromatic aryl hydrazono esters. Parasitol. Res. 2012, 111, 975–982. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, Q.K.; Hu, D.F.; Li, K. Sensilla on the antennal funiculus of the horse stomach bot fly, Gasterophilus nigricornis. Med. Vet. Entomol. 2012, 26, 314–322. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, Q.K.; Hu, D.F.; Li, K. Cuticular structures on antennae of the bot fly, Portschinskia magnifica (Diptera: Oestridae). Parasitol. Res. 2012, 111, 1651–1659. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, Q.K.; Liu, X.H.; Li, K. Sensilla on antenna and maxillary palp of predaceous fly, Lispe neimongola Tian et Ma (Diptera: Muscidae). Micron 2013, 49, 33–39. [Google Scholar] [CrossRef] [PubMed]
- Klowden, M.J. Physiological Systems in Insects, 2nd ed.; Elsevier Academic Press: London, UK, 2007; pp. 1–579. [Google Scholar]
- Elgar, M.A.; Zhang, D.; Wang, Q.K.; Wittwer, B.; Pham, H.T.; Johnson, T.L.; Freelance, C.B.; Coquilleau, M. Insect antennal morphology: The evolution of diverse solutions to odorant perception. Yale J. Biol. Med. 2018, 91, 457–469. [Google Scholar] [PubMed]
- Wang, Q.K.; Shang, Y.D.; Hilton, D.S.; Inthavong, K.; Zhang, D.; Elgar, M.A. Antennal scales improve signal detection efficiency in moths. Proc. R. Soc. Lond. Ser. B 2018, 285, 20172832. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ross, K.T.A. Comparative study of the antennal sensilla of five species of root maggots: Delia radicum, L., D. floralis, F., D. antiqua, Mg., D. platura, MG. (Diptera: Anthomyiidae) and Psila rosae, F. (Diptera: Psilidae). Int. J. Insect Morphol. Embryol. 1992, 21, 175–197. [Google Scholar] [CrossRef]
- Sukontason, K.; Sukontason, K.L.; Piangjai, S.; Boonchu, N.; Chaiwong, T.; Ngern-klun, R.; Sripakdee, D.; Vogtsberger, R.C.; Olson, J.K. Antennal sensilla of some forensically important flies in families Calliphoridae, Sarcophagidae and Muscidae. Micron 2004, 35, 671–679. [Google Scholar] [CrossRef]
- Yeates, D.K.; Wiegmann, B.M. Phylogeny and evolution of Diptera: Recent insights and new perspectives. In The Evolutionary Biology of Flies; Yeates, D.K., Wiegmann, B.M., Eds.; Columbia University Press: New York, NY, USA, 2005; pp. 14–44. [Google Scholar]
- Wang, Q.K.; Liu, X.H.; Lu, P.F.; Zhang, D. Ultrastructure of antennal sensilla in Hydrotaea armipes (Fallén) (Diptera: Muscidae): New evidence for taxonomy of the genus Hydrotaea. Zootaxa 2014, 3790, 577–586. [Google Scholar] [CrossRef] [Green Version]
- Urbanek, A.; Piotrowicz, M.; Szadziewski, R.; GiŁka, W. Sensilla coeloconica ringed by microtrichia in host-seeking biting midges. Med. Vet. Entomol. 2014, 28, 355–363. [Google Scholar] [CrossRef]
- Giroux, M.; Pape, T.; Wheeler, T.A. Towards a phylogeny of the flesh flies (Diptera: Sarcophagidae): Morphology and phylogenetic implications of the acrophallus in the subfamily Sarcophaginae. Zool. J. Linn. Soc. 2010, 158, 740–778. [Google Scholar] [CrossRef] [Green Version]
- Lambkin, C.L.; Sinclair, B.J.; Pape, T.; Courtney, G.W.; Skevington, J.H.; Meier, R.; Yeates, D.K.; Blagoderov, V.; Wiegmann, B.M. The phylogenetic relationships among infraorders and superfamilies of Diptera based on morphological evidence. Syst. Entomol. 2013, 38, 164–179. [Google Scholar] [CrossRef]
- Smith, K.M.; Lefroy, H.M. A comparative study of certain sense-organs in the antennae and palpi of Diptera. Proc. Zool. Soc. Lond. 1919, 89, 31–69. [Google Scholar] [CrossRef]
- Slifer, E.H.; Sekhon, S.S. Fine structure of the sense organs on the antennal flagellum of a flesh fly, Sarcophaga argyrostoma R.-D. (Diptera, Sarcophagidae). J. Morphol. 1964, 114, 185–207. [Google Scholar] [CrossRef] [PubMed]
- Abouzied, E.M. Antennal and maxillary palp sensillae of male and female Liosarcophaga babiyari Lehrer (Diptera: Sarcophagidae). Bull. Entomol. Soc. Egypt 2008, 85, 29–48. [Google Scholar]
- Pezzi, M.; Whitmore, D.; Chicca, M.; Semeraro, B.; Brighi, F.; Leis, M. Ultrastructural morphology of the antenna and maxillary palp of Sarcophaga tibialis (Diptera: Sarcophagidae). J. Med. Entomol. 2016, 53, 807–814. [Google Scholar] [CrossRef]
- Khedre, A.M. Olfactory sensilla on the antennae and maxillary palps of the fleshfly Wohlfahrtia nuba (Wied.) (Diptera: Sarcophagidae). J. Egypt. Ger. Soc. Zool. 1997, 24, 171–193. [Google Scholar]
- Wasserman, S.L.; Itagaki, H. The olfactory responses of the antenna and maxillary palp of the fleshfly, Neobellieria bullata (Diptera: Sarcophagidae), and their sensitivity to blockage of nitric oxide synthase. J. Insect Physiol. 2003, 49, 271–280. [Google Scholar] [CrossRef]
- Greenberg, B.; Ash, N. Setiferous plaques on antennal pedicels of muscoid diptera: Appearance in various species and tests of function. Ann. Entomol. Soc. Am. 1972, 65, 1340–1346. [Google Scholar] [CrossRef]
- Ross, K.T.A.; Anderson, M. Morphology of the antennal sensilla of the cabbage root fly, Delia radicum L. (Diptera: Anthomyiidae). Int. J. Insect Morphol. Embryol. 1987, 16, 331–342. [Google Scholar] [CrossRef]
- Kelling, F.J. Olfaction in Houseflies: Morphology and Electrophysiology. Ph.D. Thesis, University of Groningen, Groningen, The Netherlands, 2001; pp. 1–144. [Google Scholar]
- Smallegange, R.C.; Kelling, F.J.; Den Otter, C.J. Types and numbers of sensilla on antennae and maxillary palps of small and large houseflies, Musca domestica (Diptera, Muscidae). Microsc. Res. Tech. 2008, 71, 880–886. [Google Scholar] [CrossRef]
- Zhang, D.; Wang, Q.K.; Yang, Y.Z.; Chen, Y.O.; Li, K. Sensory organs of the antenna of two Fannia species (Diptera: Fanniidae). Parasitol. Res. 2013, 112, 2177–2185. [Google Scholar] [CrossRef]
- Cumming, J.M.; Wood, D.M. Adult morphology and terminology. In Manual of Afrotropical Diptera; Kirk-Spriggs, A.H., Sinclair, B.J., Eds.; South African National Biodiversity Institute: Pretoria, South Africa, 2017; Volume 1, pp. 89–133. [Google Scholar]
- Zhang, D.; Liu, X.H.; Li, X.Y.; Zhang, M.; Li, K. Antennal sensilla of the green bottle fly, Lucilia sericata (Meigen) (Diptera: Calliphoridae). Parasitol. Res. 2013, 112, 3843–3850. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Li, X.Y.; Liu, X.H.; Wang, Q.K.; Pape, T. The antenna of horse stomach bot flies: Morphology and phylogenetic implications (Oestridae, Gasterophilinae: Gasterophilus Leach). Sci. Rep. 2016, 6, 34409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, F.F.; Adserballe, C.F. Cuticular structures on the antennae of Hypoderma bovis De Geer (Diptera: Oestridae) females. Int. J. Insect Morphol. Embryol. 1996, 25, 173–181. [Google Scholar] [CrossRef]
- Wang, Q.K.; Zhang, M.; Li, K.; Zhang, D. Olfactory Sensilla on Antennae and Maxillary Palps of Fannia hirticeps (Stein, 1892) (Diptera: Fanniidae). Microsc. Res. Tech. 2012, 75, 1313–1320. [Google Scholar] [CrossRef]
- Andreani, A.; Sacchetti, P.; Belcari, A. Comparative morphology of the deer ked Lipoptena fortisetosa first recorded from Italy. Med. Vet. Entomol. 2019, 33, 140–153. [Google Scholar] [CrossRef] [Green Version]
- Liu, X.H.; Li, X.Y.; Li, K.; Zhang, D. Ultrastructure of antennal sensory organs of horse nasal-myiasis fly, Rhinoestrus purpureus (Diptera: Oestridae). Parasitol. Res. 2015, 114, 2527–2533. [Google Scholar] [CrossRef]
- Liu, X.H.; Zhang, M.; Shi, J.N.; Li, K.; Zhang, D. Ultrastructure of antennal sensilla of a parasitoid fly, Pales pavida Meigen (Diptera: Tachinidae). Micron 2013, 55, 36–42. [Google Scholar] [CrossRef]
- Liu, X.H.; Liu, J.J.; Li, X.Y.; Zhang, D. Antennal sensory organs of Scathophaga stercoraria (Linnaeus, 1758) (Diptera: Scathophagidae): Ultramorphology and phylogenetic implications. Zootaxa 2016, 4067, 361–372. [Google Scholar] [CrossRef]
- Ashworth, J.R.; Wall, R. Responses of the sheep blowflies Lucilia sericata and L. cuprina to odour and the development of semiochemical baits. Med. Vet. Entomol. 1994, 8, 303–309. [Google Scholar] [CrossRef]
- Wang, Q.K.; Yang, Y.Z.; Liu, M.Q.; Zhang, D. Fine Structure of Delia platura (Meigen) (Diptera: Anthomyiidae) Revealed by Scanning Electron Microscopy. Microsc. Res. Tech. 2014, 77, 619–630. [Google Scholar] [CrossRef]
- Greenberg, B. Species distribution of new structures on fly antennae. Nature 1970, 228, 1338–1339. [Google Scholar] [CrossRef] [PubMed]
- Liu, G.T.; Wang, Q.K.; Liu, X.H.; Li, X.Y.; Pang, X.N.; Zhang, D. Antennal and palpal sensilla of three predatory Lispe species (Diptera: Muscidae): An ultrastructural investigation. Sci. Rep. 2021, 11, 18357. [Google Scholar] [CrossRef] [PubMed]
- Archer, M.S.; Elgar, M.A. Effects of decomposition on carcass attendance in a guild of carrion-breeding flies. Med. Vet. Entomol. 2003, 17, 263–271. [Google Scholar] [CrossRef] [PubMed]
- Goff, M.L. Early postmortem changes and stages of decomposition. In Current Concepts in Forensic Entomology; Amendt, J., Campobasso, C.P., Goff, M.L., Grassberger, M., Eds.; Springer: London, UK, 2010; pp. 1–24. [Google Scholar]
- Dethier, V.G. The Physiology of Insect Senses; Chapman & Hall: London, UK, 1971; pp. 1–266. [Google Scholar]
- Chapman, R.F. Chemoreception: The significance of receptor numbers. Adv. Insect Physiol. 1982, 16, 247–356. [Google Scholar]
- Isaac, C.; Ravaiano, S.V.; Pascini, T.V.; Martins, G.F. The Antennal Sensilla of Species of the Palpalis Group (Diptera: Glossinidae). J. Med. Entomol. 2015, 52, 614–621. [Google Scholar] [CrossRef]
- Zhang, D.; Liu, X.H.; Li, X.Y.; Cao, J.; Chu, H.J.; Li, K. Ultrastructural investigation of antennae in three cutaneous myiasis flies: Melophagus ovinus, Hippobosca equina, and Hippobosca longipennis (Diptera: Hippoboscidae). Parasitol. Res. 2015, 114, 1887–1896. [Google Scholar] [CrossRef]
- Been, T.H.; Schomaker, C.H.; Thomas, G. Olfactory sensilla on the antenna and maxillary palp of the sheep head fly, Hydrotaea irritans (Fallen) (Diptera: Muscidae). Int. J. Insect Morphol. Embryol. 1988, 17, 121–133. [Google Scholar] [CrossRef]
- Sukontason, K.; Methanitikorn, R.; Chaiwong, T.; Kurahashi, H.; Vogtsberger, R.C.; Sukontason, K.L. Sensilla of the antenna and palp of Hydrotaea chalcogaster (Diptera: Muscidae). Micron 2007, 38, 218–223. [Google Scholar] [CrossRef]
- Hore, G.; Saha, G.K.; Banerjee, D. Sensory organs of forensically important fly Ophyra capensis (Wiedemann, 1818) (Diptera: Muscidae): A scanning electron microscopic study. Acta Trop. 2018, 185, 400–411. [Google Scholar] [CrossRef]
- Ross, K.T.A.; Anderson, M. Ultrastucture of the funicular sensilla of the cabbage root fly, Delia radicum L. (Diptera: Anthomyiidae). Int. J. Insect Morphol. Embryol. 1991, 20, 83–101. [Google Scholar] [CrossRef]
- Hassan, M.I.; Fouda, M.A.; Hammad, K.M.; Basiouny, A.L.; Kamel, M.R. The ultrastructure of sensilla associated with mouthparts and antennae of Lucilia cuprina. J. Egypt. Soc. Parasitol. 2013, 43, 777–785. [Google Scholar] [CrossRef]
- Felt, B.T.; Vande Berg, J.S. Ultrastructure of the blowfly chemoreceptor sensillum (Phormia regina). J. Morphol. 1976, 150, 763–783. [Google Scholar] [CrossRef] [PubMed]
- Setzu, M.D.; Poddighe, S.; Angioy, A.M. Sensilla on the antennal funiculus of the blow fly, Protophormia terraenovae (Diptera: Calliphoridae). Micron 2011, 42, 471–477. [Google Scholar] [CrossRef] [PubMed]
- Hore, G.; Maity, A.; Naskar, A.; Ansar, W.; Ghosh, S.; Saha, G.K.; Banerjee, D. Scanning electron microscopic studies on antenna of Hemipyrellia ligurriens (Wiedemann, 1830) (Diptera: Calliphoridae)—A blow fly species of forensic importance. Acta Trop. 2017, 172, 20–28. [Google Scholar] [CrossRef] [PubMed]
- Sukontason, K.; Methanitikorn, R.; Kurahashi, H.; Vogtsberger, R.C.; Sukontason, K.L. External morphology of Chrysomya pinguis (Walker) (Diptera: Calliphoridae) revealed by scanning electron microscopy. Micron 2008, 39, 190–197. [Google Scholar] [CrossRef] [PubMed]
- Zhang, D.; Liu, X.H.; Wang, Q.K.; Li, K. Sensilla on the antenna of blow fly, Triceratopyga calliphoroides Rohdendorf (Diptera: Calliphoridae). Parasitol. Res. 2014, 113, 2577–2586. [Google Scholar] [CrossRef]
- Caetano, R.L.; Carriço, C.; de Abreu Freitas, D.M.; Pinto, Z.T. Ultrastructure of sensilla on antennae and maxillary palps in three Mesembrinellidae species. Rev. Chil. Hist. Nat. 2018, 91, 7. [Google Scholar] [CrossRef]
- Rahal, Y.; Barry, P.; Hawlitzky, N.; Renou, M. Antennal olfactory sensilla of the parasitoid fly, Pseudoperichaeta nigrolineata Walker (Diptera: Tachinidae). Int. J. Insect Morphol. Embryol. 1996, 25, 145–152. [Google Scholar] [CrossRef]
- Roh, G.H.; Lee, Y.J.; Park, C.G. Morphology and distribution of antennal sensilla in a parasitoid fly, Gymnosoma rotundatum (Diptera: Tachinidae). Microsc. Res. Tech. 2020, 83, 589–596. [Google Scholar] [CrossRef]
- Giangiuliani, G.; Lucchi, A.; Vinson, S.B.; Bin, F. External anatomy of adult antennal sensilla of the fly, Trichopoda pennipes F. (Diptera: Tachinidae). Int. J. Insect Morphol. Embryol. 1994, 23, 105–113. [Google Scholar] [CrossRef]
- Poddighe, S.; Dekker, T.; Scala, A.; Angioy, A.M. Olfaction in the female sheep botfly. Naturwissenschaften 2010, 97, 827–835. [Google Scholar] [CrossRef] [PubMed]
- Li, X.Y.; Liu, X.H.; Ge, Y.Q.; Zhang, D. Scanning electron microscopy of antennal sensory organs of the cattle grub, Hypoderma lineatum (Diptera: Oestridae). Parasitol. Res. 2015, 114, 3865–3871. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Number of Sensory Pits (Ventral Surface) | Number of Sensory Pits (Dorsal Surface) | ||
|---|---|---|---|---|
| Male | Female | Male | Female | |
| Sarcophaga albiceps | 12 | − | 4 | − |
| Sarcophaga africa | 10 | − | 6 | − |
| Sarcophaga peregrina | 6 | − | 4 | − |
| Sarcophaga portschinskyi | 10 | 15 | 6 | 7 |
| Agria mihalyii | 14 | − | 11 | − |
| Wohlfahrtia magnifica | 1 | − | 1 | − |
| Wohlfahrtia bella | 6 | 10 | 2 | 16 |
| Metopia campestris | 3 | − | 1 | − |
| Mesomelena mesomelaena | 2 | 8 | 0 | 26 |
| Species | Type | Length | Basal Diameter | Degree of Basal Swelling |
|---|---|---|---|---|
| Sarcophaga albiceps | Tr | 19.64 ± 2.14 | 3.07 ± 0.35 | − |
| Ba I | 7.77 ± 0.84 | 1.94 ± 0.30 | − | |
| Ba II | 5.50 ± 0.28 | 1.04 ± 0.17 | − | |
| Co | 3.39 ± 0.66 | 0.98 ± 0.39 | − | |
| Col | 5.85 ± 0.48 | 1.98 ± 0.41 | − | |
| Cl | 7.88 ± 0.66 | 1.72 ± 0.24 | − | |
| BSS | 7.02 ± 0.38 | 1.76 ± 0.22 | 0.51 ± 0.03 | |
| Sarcophaga portschinskyi | Tr | 15.71 ± 1.24 | 2.51 ± 0.37 | − |
| Ba I | 9.01 ± 0.93 | 1.81 ± 0.25 | − | |
| Ba II | 6.43 ± 0.88 | 1.85 ± 0.32 | − | |
| Co | 3.45 ± 0.84 | 1.06 ± 0.25 | − | |
| Cl | 9.35 ± 1.18 | 1.85 ± 0.25 | − | |
| BSS | 7.69 ± 1.05 | 2.46 ± 0.55 | 0.41 ± 0.01 | |
| Sarcophaga africa | Tr | 18.30 ± 1.43 | 2.18 ± 0.27 | − |
| Ba I | 7.58 ± 0.40 | 1.87 ± 0.42 | − | |
| Ba II | 6.88 ± 0.56 | 1.28 ± 0.04 | − | |
| Co | 3.92 ± 0.53 | 1.35 ± 0.19 | − | |
| Cl | 6.80 ± 0.40 | 1.33 ± 0.42 | − | |
| BSS | 8.02 ± 0.69 | 1.81 ± 0.30 | 0.35 ± 0.01 | |
| Sarcophaga peregrina | Tr | 21.02 ± 3.43 | 2.20 ± 0.41 | − |
| Ba I | 9.36 ± 1.43 | 1.37 ± 0.46 | − | |
| Ba II | 6.00 ± 0.82 | 1.53 ± 0.12 | − | |
| Co | 3.41 ± 0.42 | 1.02 ± 0.09 | − | |
| Col | 8.30 ± 0.32 | 1.72 ± 0.21 | − | |
| Cl | 8.64 ± 0.96 | 1.87 ± 0.20 | − | |
| BSS | 9.42 ± 1.00 | 2.15 ± 0.23 | 0.21 ± 0.02 | |
| Wohlfahrtia bella | Tr | 18.23 ± 2.4 | 2.30 ± 0.26 | − |
| Ba I | 10.15 ± 0.54 | 1.45 ± 0.25 | − | |
| Ba II | 6.57 ± 0.21 | 1.74 ± 0.56 | − | |
| Co | 3.04 ± 1.13 | 1.25 ± 0.22 | − | |
| Cl | 9.53 ± 0.82 | 1.56 ± 0.25 | − | |
| BSS | 11.28 ± 1.31 | 2.32 ± 0.42 | 0.25 ± 0.01 | |
| Wohlfahrtia magnifica | Tr | 16.51 ± 1.64 | 2.47 ± 0.15 | − |
| Ba I | 10.12 ± 2.51 | 1.90 ± 0.65 | − | |
| Ba II | 5.84 ± 0.52 | 1.20 ± 0.32 | − | |
| Co | 3.48 ± 0.04 | 0.75 ± 0.02 | − | |
| Col | 4.57 ± 0.44 | 1.17 ± 0.01 | − | |
| Cl | 11.30 ± 1.32 | 1.70 ± 0.14 | − | |
| BSS | 9.29 ± 0.60 | 1.74 ± 0.14 | 0.34 ± 0.01 | |
| Agria mihalyii | Tr | 19.16 ± 1.49 | 2.90 ± 0.26 | − |
| Ba I | 7.70 ± 1.18 | 1.34 ± 0.38 | − | |
| Ba II | 6.00 ± 0.23 | 1.82 ± 0.77 | − | |
| Ba III | 4.86 ± 0.09 | 2.20 ± 0.81 | − | |
| Co | 2.93 ± 0.30 | 1.16 ± 0.05 | − | |
| Cl | 8.62 ± 0.66 | 1.68 ± 0.36 | − | |
| BSS | 7.89 ± 1.02 | 4.80 ± 0.84 | 0.09 ± 0.01 | |
| Metopia campestris | Tr | 19.07 ± 2.48 | 2.33 ± 0.48 | − |
| Ba I | 9.07 ± 1.44 | 1.50 ± 0.08 | − | |
| Ba II | 5.50 ± 0.45 | 1.35 ± 0.22 | − | |
| Co | 3.44 ± 0.49 | 1.09 ± 0.12 | − | |
| Cl | 8.90 ± 0.99 | 1.35 ± 0.43 | − | |
| BSS | 6.43 ± 0.27 | 1.81 ± 0.08 | 0.42 ± 0.02 | |
| Mesomelena mesomelaena | Tr | 20.05 ± 2.56 | 2.50 ± 0.56 | − |
| Ba I | 10.25 ± 1.23 | 1.50 ± 0.03 | − | |
| Ba II | 6.70 ± 1.62 | 1.25 ± 0.20 | − | |
| Co | 3.45 ± 0.24 | 1.20 ± 0.14 | − | |
| Cl | 10.05 ± 0.24 | 1.58 ± 0.36 | − | |
| BSS | 10.64 ± 0.45 | 2.05 ± 0.32 | 0.23 ± 0.01 |
| Species | Type | Anterior Surface | Posterior Surface | ||||
|---|---|---|---|---|---|---|---|
| Proximal | Median | Distal | Proximal | Median | Distal | ||
| Sarcophaga. albiceps | Tr | 4.60 ± 2.51 | 13.00 ± 1.92 | 13.60 ± 1.14 | 1.00 ± 1.00 | 10.00 ± 2.92 | 11.40 ± 3.36 |
| Ba | 0.00 | 11.40 ± 1.67 | 6.20 ± 1.79 | 0.00 | 9.20 ± 0.84 | 8.00 ± 2.12 | |
| Co | 0.00 | 0.00 | 1.40 ± 0.55 | 0.20 ± 0.45 | 3.00 ± 1.41 | 2.80 ± 0.84 | |
| Cl | 7.00 ± 1.22 | 0.00 | 0.00 | 4.20 ± 1.10 | 0.00 | 0.00 | |
| Sarcophaga portschinskyi | Tr | 2.80 ± 2.77 | 14.00 ± 4.53 | 22.20 ± 0.84 | 0.00 | 6.60 ± 2.19 | 13.40 ± 1.95 |
| Ba | 0.00 | 9.40 ± 2.07 | 8.60 ± 4.67 | 0.00 | 4.80 ± 1.64 | 5.20 ± 0.84 | |
| Co | 0.20 ± 0.45 | 0.00 | 4.00 ± 2.55 | 0.20 ± 0.45 | 0.20 ± 0.45 | 1.60 ± 1.34 | |
| Cl | 7.60 ± 2.30 | 0.00 | 0.00 | 5.00 ± 1.22 | 0.00 | 0.00 | |
| Sarcophaga africa | Tr | 9.80 ± 6.26 | 12.00 ± 2.74 | 20.80 ± 4.02 | 1.00 ± 1.00 | 4.00 ± 2.55 | 16.80 ± 2.17 |
| Ba | 0.00 | 12.20 ± 1.79 | 7.60 ± 1.52 | 0.00 | 4.00 ± 1.22 | 4.40 ± 1.14 | |
| Co | 0.20 ± 0.45 | 0.20 ± 0.45 | 2.60 ± 1.52 | 0.40 ± 0.89 | 0.00 | 2.20 ± 1.30 | |
| Cl | 7.20 ± 1.79 | 0.00 | 0.00 | 6.00 ± 1.00 | 0.00 | 0.00 | |
| Sarcophaga peregrina | Tr | 9.20 ± 1.48 | 9.80 ± 1.92 | 18.20 ± 3.63 | 0.00 | 4.20 ± 1.92 | 12.80 ± 2.49 |
| Ba | 0.00 | 7.60 ± 2.07 | 6.40 ± 2.30 | 0.00 | 7.00 ± 2.92 | 6.80 ± 4.60 | |
| Co | 0.80 ± 0.84 | 1.60 ± 1.14 | 2.60 ± 0.55 | 1.00 ± 0.71 | 1.20 ± 1.30 | 2.20 ± 1.79 | |
| Cl | 2.00 ± 1.58 | 0.00 | 0.00 | 7.00 ± 2.35 | 0.00 | 0.00 | |
| Wohlfahrtia bella | Tr | 4.25 ± 1.56 | 6.20 ± 3.25 | 21.00 ± 2.45 | 1.00 ± 1.24 | 5.60 ± 2.12 | 15.02 ± 1.67 |
| Ba | 0.00 | 3.50 ± 0.75 | 2.40 ± 1.63 | 0.00 | 1.20 ± 0.40 | 1.40 ± 1.45 | |
| Co | 0.00 | 0.40 ± 1.20 | 0.30 ± 0.54 | 0.00 | 0.00 | 0.20 ± 0.45 | |
| Cl | 1.50 ± 0.30 | 0.00 | 0.00 | 0.60 ± 0.25 | 0.00 | 0.00 | |
| Wohlfahrtia magnifica | Tr | 2.27 ± 2.85 | 5.33 ± 2.88 | 13.61 ± 4.54 | 1.81 ± 1.89 | 3.33 ± 5.41 | 13.04 ± 3.52 |
| Ba | 0.00 | 0.11 ± 0.51 | 0.79 ± 1.33 | 0.00 | 0.30 ± 0.80 | 1.13 ± 1.73 | |
| Co | 0.00 | 0.23 ± 0.70 | 0.23 ± 0.70 | 0.00 | 0.00 | 0.23 ± 0.70 | |
| Cl | 0.23 ± 0.70 | 0.00 | 0.00 | 0.45 ± 1.19 | 0.00 | 0.00 | |
| Agria mihalyii | Tr | 0.40 ± 0.55 | 7.00 ± 1.58 | 19.00 ± 4.24 | 0.20 ± 0.45 | 3.80 ± 1.64 | 17.20 ± 5.22 |
| Ba | 0.00 | 3.20 ± 1.79 | 6.00 ± 3.32 | 0.00 | 0.80 ± 1.30 | 7.00 ± 1.87 | |
| Co | 0.00 | 0.20 ± 0.45 | 1.60 ± 1.14 | 0.00 | 0.20 ± 0.45 | 1.20 ± 1.30 | |
| Cl | 1.60 ± 1.52 | 0.00 | 0.00 | 0.60 ± 0.55 | 0.00 | 0.00 | |
| Metopia campestris | Tr | 18.20 ± 7.19 | 30.40 ± 3.78 | 35.60 ± 3.51 | 14.00 ± 7.91 | 25.40 ± 6.88 | 34.00 ± 4.85 |
| Ba | 0.00 | 3.20 ± 1.64 | 2.00 ± 1.87 | 0.00 | 3.60 ± 0.89 | 1.40 ± 0.89 | |
| Co | 0.00 | 1.20 ± 1.30 | 2.00 ± 1.41 | 0.00 | 1.40 ± 1.52 | 2.00 ± 1.41 | |
| Cl | 3.00 ± 2.12 | 0.00 | 0.00 | 1.60 ± 1.34 | 0.00 | 0.00 | |
| Mesomelena mesomelaena | Tr | 17.50 ± 3.52 | 28.60 ± 2.63 | 32.20 ± 0.51 | 9.20 ± 3.62 | 22.30 ± 2.74 | 30.60 ± 0.62 |
| Ba | 0.00 | 2.54 ± 0.73 | 4.00 ± 2.43 | 0.00 | 2.40 ± 0.65 | 2.20 ± 1.32 | |
| Co | 0.00 | 2.40 ± 0.86 | 1.60 ± 0.13 | 0.00 | 0.45 ± 3.41 | 1.30 ± 0.70 | |
| Cl | 3.20 ± 0.23 | 0.00 | 0.00 | 0.00 | 0.00 | 0.00 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, W.; Liu, G.; Wang, Q.; Yan, L.; Liu, X.; Li, X.; Pape, T.; Zhang, D. Ultrastructure of Antennal Sensory Organs in Nine Flesh Flies (Diptera: Sarcophagidae): New Insight into the Definition of Family Sarcophagidae. Insects 2022, 13, 602. https://doi.org/10.3390/insects13070602
Xu W, Liu G, Wang Q, Yan L, Liu X, Li X, Pape T, Zhang D. Ultrastructure of Antennal Sensory Organs in Nine Flesh Flies (Diptera: Sarcophagidae): New Insight into the Definition of Family Sarcophagidae. Insects. 2022; 13(7):602. https://doi.org/10.3390/insects13070602
Chicago/Turabian StyleXu, Wentian, Genting Liu, Qike Wang, Liping Yan, Xianhui Liu, Xinyu Li, Thomas Pape, and Dong Zhang. 2022. "Ultrastructure of Antennal Sensory Organs in Nine Flesh Flies (Diptera: Sarcophagidae): New Insight into the Definition of Family Sarcophagidae" Insects 13, no. 7: 602. https://doi.org/10.3390/insects13070602