
Larval Crowding Did Not Enhance Adult Migration Propensity in Spodoptera frugiperda


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Larval Density Treatment
2.3. Pupal Weight, Body Weight and Wing Expansion
2.4. Reproductive Parameter Determination
2.5. Flight Capacity Determination
2.6. Data Analysis
3. Results
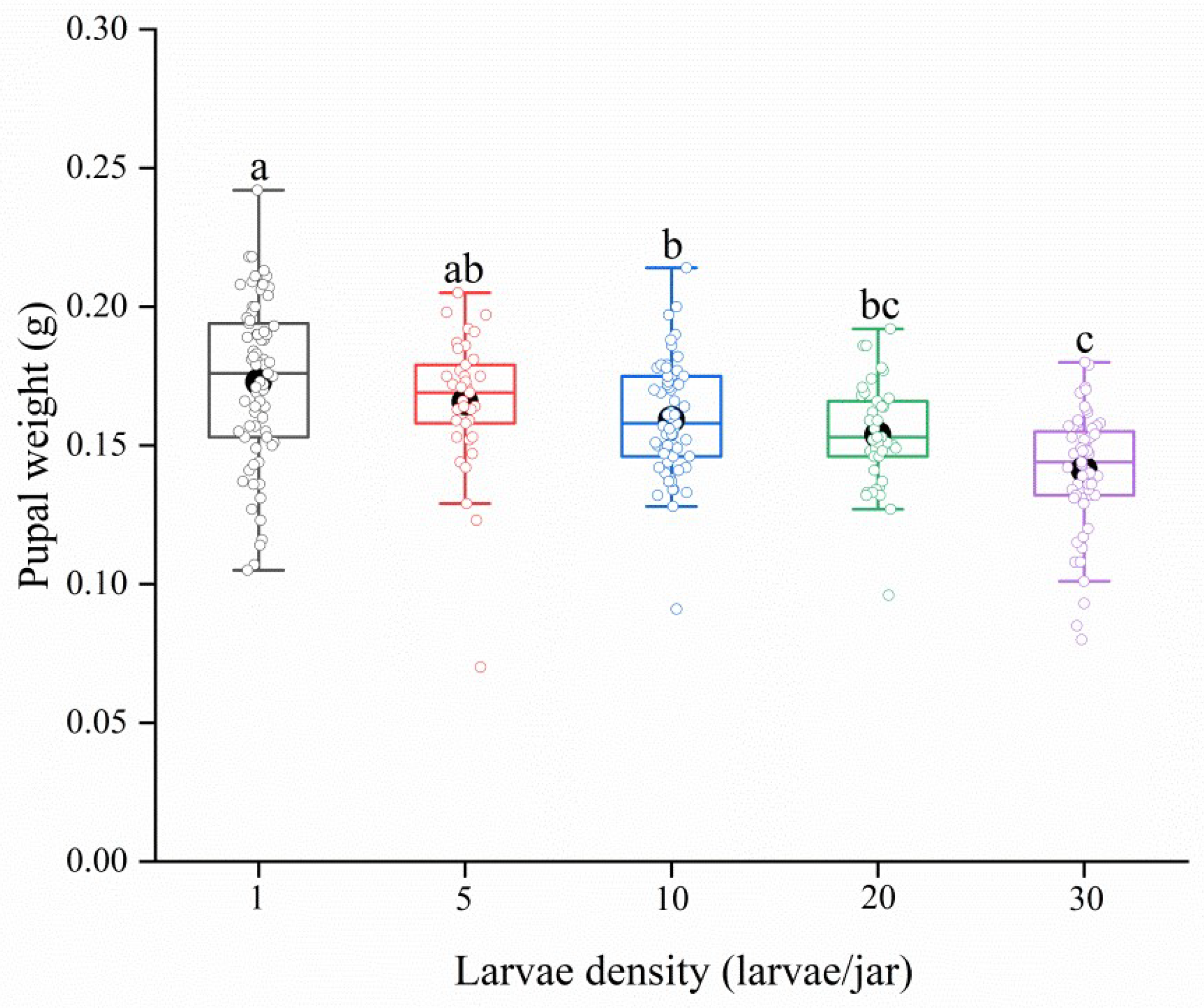
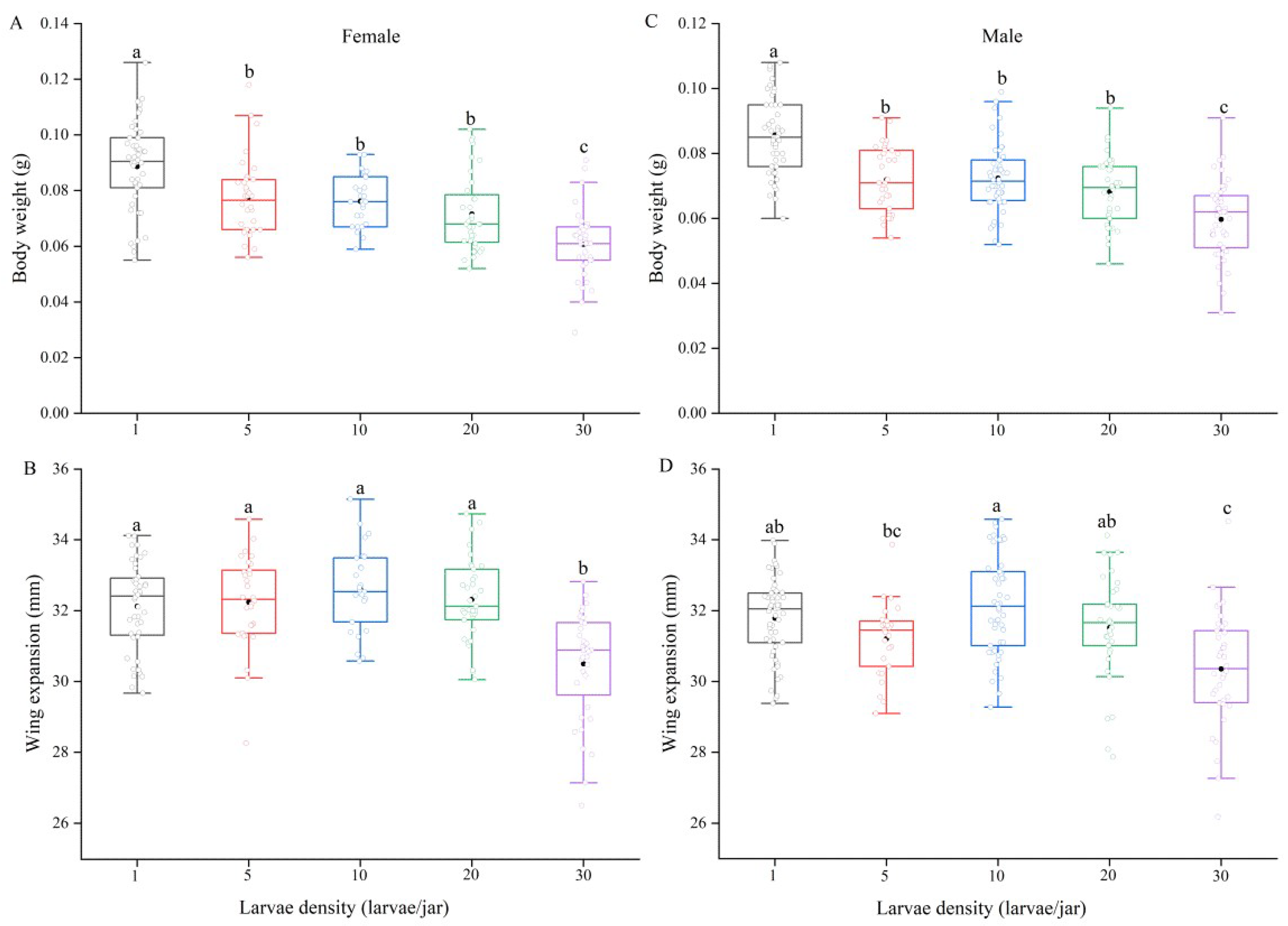
3.1. Effect of Larval Density on Pupal Weight, Body Weight and Wing Expansion in Spodoptera frugiperda
3.2. Effect of Larval Density on Reproduction in Spodoptera frugiperda
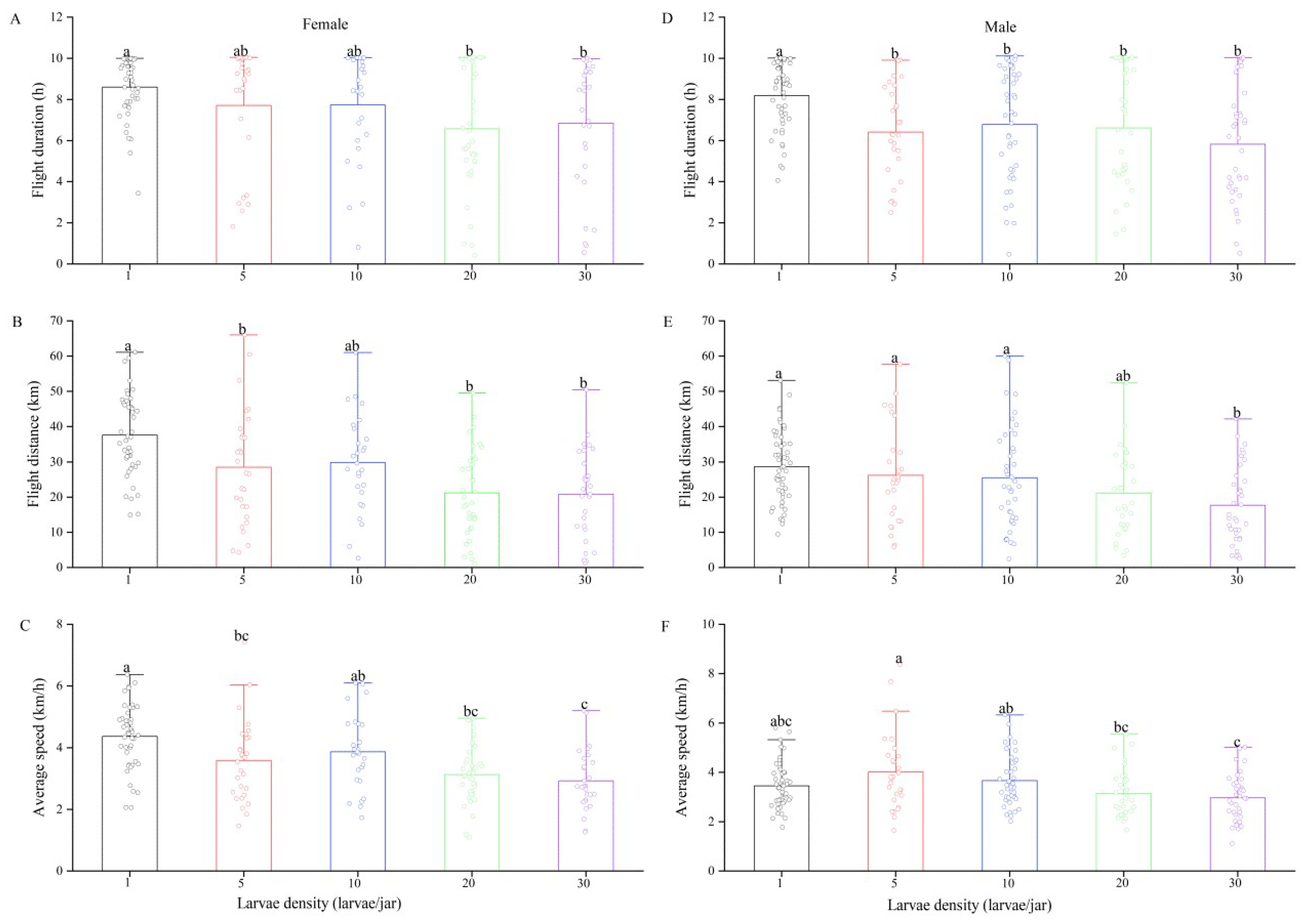
3.3. Effect of Larval Density on Flight Capacity in Spodoptera frugiperda
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Shenoi, V.N.; Syed, Z.A.; Prasad, N.G. Evolution of increased adult longevity in Drosophila melanogaster populations selected for adaptation to larval crowding. J. Evol. Biol. 2015, 29, 407–417. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisher, A.M.; Page, S.L.; Manser, A.; Lewis, D.R.; Holwell, G.I.; Wigby, S. Relatedness modulates density-dependent cannibalism rates in drosophila. Funct. Ecol. 2020, 89, 1788–1798. [Google Scholar] [CrossRef]
- He, H.l.; Zhou, A.L.; He, L.; Qiu, L.; Ding, W.B.; Li, Y. The frequency of cannibalism by Spodoptera frugiperda larvae determines their probability of surviving food deprivation. J. Pest. Sci. 2022, 95, 145–157. [Google Scholar] [CrossRef]
- Luo, L.Z.; Li, G.B.; Cao, Y.Z.; Hu, Y. The influence of larval rearing density on flight capacity and fecundity of adult oriental armyworm, Mythimna seperata (Walker). Acta Entomol. Sin. 1995, 38, 38–45. [Google Scholar]
- Santos, M.; Borash, D.J.; Joshi, A.; Bounlutay, N.; Mueller, L.D. Density-dependent natural selection in Drosophila: Evolution of growth rate and body size. Evolution 1997, 51, 420–432. [Google Scholar]
- Coll, M.; Gavish, S.; Dori, I. Population biology of the potato tuber moth, Phthorimaea operculella (Lepidoptera: Gelechiidae), in two potato cropping systems in Israel. Bull. Entomol. Res. 2000, 90, 309–315. [Google Scholar] [CrossRef] [Green Version]
- Morales-Ramos, J.A.; Rojas, M.G. Effect of larval density on food utilization efficiency of Tenebrio molitor (coleoptera: Tenebrionidae). J. Econ Entomol. 2015, 108, 2259–2267. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Kang, L. Molecular mechanisms of phase change in locusts. Annu. Rev. Entomol. 2014, 59, 225–244. [Google Scholar] [CrossRef] [Green Version]
- Zera, A.J.; Denno, R.F. Physiology and ecology of dispersal polymorphism in insects. Annu. Rev. Entomol. 1997, 42, 207–230. [Google Scholar] [CrossRef] [Green Version]
- Kong, H.L.; Luo, L.Z.; Jiang, X.F.; Zhang, L. Effects of larval density on flight potential of the beet webworm, Loxostege sticticalis (Lepidoptera: Pyralidae). Environ. Entomol. 2010, 39, 1579–1585. [Google Scholar] [CrossRef]
- Cheng, Y.X.; Sappington, T.W.; Luo, L.Z.; Zhang, L.; Jiang, X.F. Starvation on first or second day of adulthood reverses larval-stage decision to migrate in beet webworm (lepidoptera: Pyralidae). Environ. Entomol. 2021, 50, 523–531. [Google Scholar] [CrossRef] [PubMed]
- Lv, W.X.; Xie, X.C. Effect of fluctuating temperatures on development, reproduction and energy of oriental armyworm populations, Mythimna separata. J. Appl. Entomol. 2022, 146, 511–524. [Google Scholar] [CrossRef]
- Chapman, J.W.; Reynolds, D.R.; Wilson, K. Long-range seasonal migration in insects: Mechanisms, evolutionary drivers and ecological consequences. Ecol. Lett. 2015, 18, 287–302. [Google Scholar] [CrossRef] [Green Version]
- Johnson, C.G. Migration and Dispersal of Insects by Flight; Methuen: London, UK, 1969. [Google Scholar]
- Du, B.; Ding, D.; Ma, C.; Guo, W.; Kang, L. Locust density shapes energy metabolism and oxidative stress resulting in divergence of flight traits. Proc. Natl. Acad. Sci. USA 2022, 119, e2115753118. [Google Scholar] [CrossRef]
- Zhang, L.; Cheng, L.L.; Chapman, J.W.; Sappington, T.W.; Liu, J.J.; Cheng, Y.X.; Jiang, X.F. Juvenile hormone regulates the shift from migrants to residents in adult oriental armyworm, Mythimna separata. Sci. Rep. 2020, 10, 11626. [Google Scholar] [CrossRef]
- Zhang, L.; Jiang, X.F.; Luo, L.Z. Determination of sensitive stage for switching migrant oriental armyworms into residents. Environ. Entomol. 2008, 37, 1389–1395. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Luo, L.Z.; Jiang, X.F. Starvation influences allatotropin gene expression and juvenile hormone titer in the female adult oriental armyworm, Mythimna separate. Arch. Insect Biochem. Physiol. 2008, 68, 63–70. [Google Scholar] [CrossRef]
- Sappington, T.W.; Showers, W.B. Lack of translation of density-induced morphological polyphenism to long duration flight behavior of black cutworm (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 1992, 85, 188–194. [Google Scholar] [CrossRef]
- Hughes, J.; Herm, A.; Dorn, S. Preimaginal environment influences adult flight in Cydia molesta (Lepidoptera: Tortricidae). Environ. Entomol. 2004, 33, 1155–1162. [Google Scholar] [CrossRef]
- Poirier, L.M.; Borden, J.H. Some effects of population density on the life history of the obliquebanded leafroller Choristoneura rosaceana Harris (Lep., Tortricidae). J. Appl. Entomol. 1992, 113, 307–314. [Google Scholar] [CrossRef]
- Fantinou, A.A.; Tsitsipis, J.A. Effect of larval density on development and diapause of Sesamia nonagrioides (Lef.) (Lep. Noctuidae) under laboratory conditions. J. Appl. Entomol. 1999, 123, 187–190. [Google Scholar] [CrossRef]
- Fox, R.L. Cannibalism in natural populations. Ann. Rev. Ecol. Syst. 1975, 6, 87–106. [Google Scholar] [CrossRef]
- Vijendravarma, R.K.; Narasimha, S.; Kawecki, T.J. Predatory cannibalism in Drosophila melanogaster larvae. Nat. Commun. 2013, 4, 1789. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wise, D.H. Cannibalism, food limitation, intraspecific competition of spider populations. Annu. Rev. Entomol. 2006, 51, 441–465. [Google Scholar] [CrossRef] [PubMed]
- Ye, J.W.; Xu, Q.Y.; Li, Z.G.; Lu, X.; Han, S.C. Effect of cannibalism on the growth and development of Mallada basalis (Neuroptera: Chrysopidae). Fla. Entomol. 2014, 97, 1075–1080. [Google Scholar] [CrossRef]
- Siva-Jothy, J.A.; Monteith, K.M.; Vale, P.F. Navigating infection risk during oviposition and cannibalistic foraging in a holometabolous insect. Behav. Ecol. 2018, 29, 1426–1435. [Google Scholar] [CrossRef]
- Tsuyoshi, K.; Kenji, F.; Takahisa, M. Egg laying preference, larval dispersion, and cannibalism in Helicoverpa armigera (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 2003, 6, 793–798. [Google Scholar]
- Wang, J.; Jiang, X.F.; Wu, D.L.; Luo, L.Z. Effect of larval rearing density on development and fecundity of the beet armyworm, Spodoptera exigua (Lepidoptera: Noctuidae). Acta Entomol. Sin. 2020, 51, 889–894. [Google Scholar]
- Tang, T.; Zhao, C.Q.; Li, X.; Qiu, L.H. Factors affecting larval cannibalism in the cotton bollworm, Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). Orient. Insects 2016, 50, 23–33. [Google Scholar] [CrossRef]
- Barros, E.M.; Torres, J.B.; Bueno, A.F. Oviposition, development, and reproduction of Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae) fed on different hosts of economic importance. Neotrop. Entomol. 2010, 39, 996–1001. [Google Scholar] [CrossRef] [Green Version]
- Westbrook, J.K.; Nagoshi, R.N.; Meagher, R.L.; Fleischer, S.J.; Jairam, S. Modeling seasonal migration of fall armyworm moths. Int. J. Biometeorol. 2016, 60, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Baudron, F.; Zaman-Allah, M.A.; Chaipa, I.; Chari, N.; Chinwada, P. Understanding the factors influencing fall armyworm (Spodoptera frugiperda JE Smith) damage in African smallholder maize fields and quantifying its impact on yield. A case study in Eastern Zimbabwe. Crop Prot. 2019, 120, 141–150. [Google Scholar] [CrossRef]
- Goergen, G.; Kumar, P.L.; Sankung, S.B.; Togola, A.; Tamò, M. First report of outbreaks of the fall armyworm Spodoptera frugiperda (J E Smith) (Lepidoptera, Noctuidae), a new alien invasive pest in West and Central Africa. PLoS ONE 2016, 11, e165632. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Stokstad, E. New crop pest takes Africa at lightning speed. Science 2017, 356, 473–474. [Google Scholar] [CrossRef]
- Sharanabasappa, D.; Kalleshwaraswamy, C.M.; Asokan, R.; Swamy, M.H.M.; Maruthi, M.S.; Pavithra, H.B.; Hegde, K.; Navi, S.; Prabhu, S.T.; Goergen, G. First report of the fall armyworm, Spodaptera frugiperda (J E Smith) (Lepidoptera: Noctuidae), an alien invasive pest on maize in India. Pest. Manag. Hortic. Ecosyst. 2018, 24, 23–29. [Google Scholar]
- Guo, J.; Zhao, J.; He, K.; Zhang, F.; Wang, Z. Potential invasion of the crop-devastating insect pest fall armyworm Spodoptera frugiperda to China. Plant Prot. 2018, 44, 1–10. [Google Scholar]
- Sun, X.X.; Hu, C.X.; Jia, H.R.; Wu, Q.L.; Shen, X.J.; Zhao, S.Y.; Jiang, Y.Y.; Wu, K.M. Case study on the first immigration of fall armyworm Spodoptera frugiperda invading into China. J. Integr. Agric. 2019, 20, 664–672. [Google Scholar] [CrossRef]
- Jing, D.P.; Guo, J.F.; Jiang, Y.Y.; Zhao, J.Z.; Sethi, A.; He, K.L.; Wang, Z.Y. Initial detections and spread of invasive Spodoptera frugiperda in China and comparisons with other noctuid larvae in cornfields using molecular techniques. Insect Sci. 2020, 27, 780–790. [Google Scholar] [CrossRef]
- Jiang, Y.Y.; Liu, J.; Xie, M.C.; Li, Y.H.; Yang, J.J.; Zhang, M.L.; Qiu, K. Observation on law of diffusion damage of Spodoptera frugiperda in China in 2019. Plant Prot. 2019, 45, 10–19. [Google Scholar]
- Song, Y.; Yang, X.; Zhang, H.; Zhang, D.; He, W.; Wyckhuys, K.A.G.; Wu, K.M. Interference competition and predation between invasive and native herbivores in maize. J. Pest. Sci. 2021, 94, 1053–1063. [Google Scholar] [CrossRef]
- Wang, D.T.; Zhang, L.; Cheng, Y.X.; Jing, X.F. Larval stage related cannibalism in the fall armyworm, Spodoptera frugiperda. Plant Prot. 2020, 46, 94–98. [Google Scholar]
- Chapman, J.W.; Trevor, W.; Ana, E.; Primitivo, C.; Cave, R.D.; Dave, G. Fitness consequences of cannibalism in the fall armyworm, Spodoptera frugiperda. Behav. Ecol. 1999, 3, 298–303. [Google Scholar] [CrossRef] [Green Version]
- Pener, M.P.; Simpson, S.J. Locust phase polyphenism: An update. Adv. Insect Physiol. 2009, 36, 1–272. [Google Scholar]
- Johnson, S.J. Migration and the life history strategy of the fall armyworm, Spodoptera frugiperda in the Western Hemisphere. Int. J. Trop. Insect Sci. 1987, 8, 543–549. [Google Scholar] [CrossRef]
- Nagoshi, R.N.; Nagoshi, B.Y.; Cañarte, E.; Navarrete, B.; Solórzano, R.; Garcés-Carrera, S. Genetic characterization of fall armyworm (Spodoptera frugiperda) in Ecuador and comparisons with regional populations identify likely migratory relationships. PLoS ONE 2019, 14, e0222332. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.X.; Luo, L.Z.; Jiang, X.F.; Sappington, T.W. Synchronized oviposition triggered by migratory flight intensifies larval outbreaks of beet webworm. PLoS ONE 2012, 7, e31562. [Google Scholar] [CrossRef] [Green Version]
- Lv, W.X.; Jiang, X.F.; Chen, X.J.; Cheng, Y.X.; Xia, J.X.; Zhang, L. Flight and reproduction variations of rice leaf roller, Cnaphalocrocis medinalis in response to different rearing temperatures. Insects 2021, 12, 1083. [Google Scholar] [CrossRef]
- Zhang, L.; Pan, P.; Sappington, T.W.; Lu, W.X.; Luo, L.Z.; Jiang, X.F. Accelerated and synchronized oviposition induced by flight of young females may intensify larval outbreaks of the rice leaf roller. PLoS ONE 2015, 10, e0121821. [Google Scholar] [CrossRef] [Green Version]
- Li, Y.; Jiang, X.F.; Zhang, L.; Cheng, Y.X.; Liu, Y.Q.; Luo, L.Z. Effects of larval density on the growth, development and reproduction of Athetis lepigone (Moschler). Chin. J. Appl. Entomol. 2014, 51, 623–629. [Google Scholar]
- Andow, D.A.; Farias, J.R.; Horikoshi, R.J.; Bernardi, D.; Nascimento, A.R.; Omoto, C. Dynamics of cannibalism in equal-aged cohorts of Spodoptera frugiperda. Ecol. Entomol. 2015, 40, 229–236. [Google Scholar] [CrossRef]
- Bolívar-Silva, D.A.; Guedes, N.; Guedes, R. Larval cannibalism and fitness in the stored grain weevils Sitophilus granarius and Sitophilus zeamais. J. Pest. Sci. 2018, 91, 707–716. [Google Scholar] [CrossRef]
- Knorr, I.B.; Bashev, A.N.; Alekseev, A.A.; Naumova, E.N. Effect of population density on ecological characteristics of the grass moth Loxostege sticticalis L. (Lepidoptera: Pyralidae) in the gradation cycle. Biol. Bull. Russ. Acad. Sci. 2000, 27, 63–70. [Google Scholar]
- Ferguson, H.J. Rearing Density Effects on Premigrant Traits of the Fall Armyworm, Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae). Ph.D. Thesis, Virginia Polytechnic Institute and State University, Blacksburg, VG, USA, 1992. [Google Scholar]
- Parker, W.E.; Gatehouse, A.G. The effect of larval rearing conditions of flight performance in females of the African armyworm, Spodoptera exempta (Walker) (Lepidoptera: Noctuidae). Bull. Entomol. Res. 1985, 75, 35–48. [Google Scholar] [CrossRef]
- Jiang, X.F.; Luo, L.Z.; Sappington, T.W. Relationship of flight and reproduction in beet armyworm, Spodoptera exigua (Lepidoptera: Noctuidae), a migrant lacking the oogenesis-flight syndrome. J. Insect Physiol. 2010, 56, 1631–1637. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.W.; Li, W.P.; Zhang, J.; Liu, X.D.; Zhai, B.P.; Hu, G. Cnaphalocrocis medinalis moths decide to migrate when suffering nutrient shortage on the first day after emergence. Insects 2019, 10, 364. [Google Scholar] [CrossRef] [Green Version]
- Jiang, X.F.; Luo, Z.L.; Zhang, L.; Sappington, T.W.; Hu, Y. Regulation of migration in the oriental armyworm, Mythimna separata (Walker) in China: A review integrating environmental, physiological, hormonal, genetic, and molecular factors. Environ. Entomol. 2011, 40, 516–533. [Google Scholar] [CrossRef]
- Yang, F.; Hu, G.; Shi, J.J.; Zhai, B.P. Effects of larval density and food stress on life-history traits of Cnaphalocrocis medinalis (lepidoptera: Pyralidae). J. Appl. Entomol. 2015, 139, 370–380. [Google Scholar] [CrossRef]
- Wu, K.M.; Guo, Y.Y. Effects of food quality and larval density on flight capacity of cotton bollworm. Acta Entomol. Sin. 1997, 40, 51–57. [Google Scholar]
- Ferguson, H.J.; Eaton, J.L.; Rogers, C.E.; Simmons, A.M. Rearing density on pupal weight, wing width, development, and female adult activity of the fall armyworm (Lepidoptera: Noctuidae). Ann. Entomol. Soc. Am. 1994, 87, 823–830. [Google Scholar] [CrossRef]
- Ferguson, H.J.; Eaton, J.L.; Rogers, C.E. Larval rearing density effects on lipid reserves and wing-loading in fall armyworm adults (Lepidoptera: Noctuidae). J. Agric. Entomol. 1997, 14, 369–384. [Google Scholar]
- Sun, X.X.; Zhao, S.Y.; Jin, M.H.; Zhao, H.Y.; Li, G.P.; Zhang, H.W.; Jiang, Y.Y.; Yang, X.M.; Wu, K.M. Larval spatial distribution pattern and sampling technique of the fall armyworm Spodoptera frugiperda in maize fields. Plant Prot. 2019, 45, 13–18. [Google Scholar]
- Yang, X.M.; Sun, X.X.; Zhao, H.Y.; Li, G.P.; Chi, X.C.; Jiang, Y.Y.; Wu, K.M. Population occurrence, spatial distribution and sampling technique of fall armyworm Spodoptera frugiperda in wheat fields. Plant Prot. 2020, 46, 10–16. [Google Scholar]
- Yang, X.M.; Zhao, H.Y.; Wu, K.M. Population occurrence and sampling technique of fall armyworm Spodoptera frugiperda in barley fields. Plant Prot. 2020, 46, 18–23. [Google Scholar]
- Booth, E.; Alyokhin, A.; Pinatti, S. Adult cannibalism in an oligophagous herbivore, the Colorado potato beetle. Insect Sci. 2017, 24, 295–302. [Google Scholar] [CrossRef] [PubMed]
- McDonald, G.; Cole, P.G. Factors influencing oocyte development in Mythimna convecta (Lepidoptera: Noctuidae) and their possible impact on migration in eastern Australia. Bull. Entomol. Res. 1991, 81, 175–184. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Parameters | Larval Densities | Body Weight | Wing Expansion | |||
|---|---|---|---|---|---|---|
| r | p-Value | r | p-Value | r | p-Value | |
| Preoviposition period | 0.028 | 0.642 | 0.448 | <0.001 * | 0.532 | <0.001 * |
| Period of first oviposition | 0.091 | 0.133 | 0.307 | <0.001 * | 0.328 | <0.001 * |
| Lifetime fecundity | −0.171 | 0.005 * | 0.482 | <0.001 * | 0.604 | <0.001 * |
| Oviposition period | −0.052 | 0.390 | 0.484 | <0.001 * | 0.632 | <0.001 * |
| Female longevity | −0.023 | 0.708 | 0.488 | <0.001 * | 0.605 | <0.001 * |
| Male longevity | 0.008 | 0.893 | 0.499 | <0.001 * | 0.633 | <0.001 * |
| Larvae Density (Larvae/Jar) | Pre-Oviposition Period (d) | Period of First Oviposition (d) | Lifetime Fecundity (Per Female) | Oviposition Period (d) | Female Longevity (d) | Male Longevity (d) |
|---|---|---|---|---|---|---|
| 1 | 4.74 ± 0.30 b | 1.74 ± 0.30 b | 1494.00 ± 157.27 a | 5.39 ± 0.49 a | 11.48 ± 0.66 a | 12.48 ± 0.94 a |
| 5 | 5.90 ± 0.34 ab | 1.90 ± 0.34 b | 1104.47 ± 44.05 b | 5.50 ± 0.22 a | 13.63 ± 0.52 a | 13.67 ± 0.55 a |
| 10 | 7.04 ± 0.82 a | 4.04 ± 0.82 a | 926.96 ± 80.30 b | 5.00 ± 0.32 a | 14.08 ± 1.17 a | 13.71 ± 0.97 a |
| 20 | 6.75 ± 0.52 ab | 3.75 ± 0.52 ab | 812.61 ± 63.93 b | 4.39 ± 0.32 a | 12.86 ± 0.55 a | 14.64 ± 0.65 a |
| 30 | 6.20 ± 0.57 ab | 3.20 ± 0.57 ab | 750.65 ± 91.03 b | 5.25 ± 0.48 a | 12.50 ± 0.84 a | 14.10 ± 1.01 a |
| Sex | Parameters | Flight Duration | Flight Distance | Flight Velocity | |||
|---|---|---|---|---|---|---|---|
| r | p-Value | r | p-Value | r | p-Value | ||
| Female | Larval densities | −0.147 | 0.015 * | −0.246 | <0.001 * | −0.200 | 0.001 * |
| Pupa weight | −0.046 | 0.447 | −0.012 | 0.842 | 0.001 | 0.993 | |
| Body weight | 0.714 | <0.001 * | 0.668 | <0.001 * | 0.755 | <0.001 * | |
| Wing expansion | 0.750 | <0.001 * | 0.692 | <0.001 * | 0.800 | <0.001 * | |
| Male | Larval densities | −0.168 | 0.005 * | −0.215 | <0.001 * | −0.145 | 0.016 * |
| Pupa weight | 0.067 | 0.269 | 0.122 | 0.043 * | 0.109 | 0.072 | |
| Body weight | 0.602 | <0.001 * | 0.543 | <0.001 * | 0.596 | <0.001 * | |
| Wing expansion | 0.697 | <0.001 * | 0.610 | <0.001 * | 0.723 | <0.001 * | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lü, W.; Meng, L.; Jiang, X.; Cheng, Y.; Zhang, L. Larval Crowding Did Not Enhance Adult Migration Propensity in Spodoptera frugiperda. Insects 2022, 13, 581. https://doi.org/10.3390/insects13070581
Lü W, Meng L, Jiang X, Cheng Y, Zhang L. Larval Crowding Did Not Enhance Adult Migration Propensity in Spodoptera frugiperda. Insects. 2022; 13(7):581. https://doi.org/10.3390/insects13070581
Chicago/Turabian StyleLü, Weixiang, Linghe Meng, Xingfu Jiang, Yunxia Cheng, and Lei Zhang. 2022. "Larval Crowding Did Not Enhance Adult Migration Propensity in Spodoptera frugiperda" Insects 13, no. 7: 581. https://doi.org/10.3390/insects13070581