
Characterization of Three Heat Shock Protein Genes in Pieris melete and Their Expression Patterns in Response to Temperature Stress and Pupal Diapause

 ,
, 

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
2.2. RNA Isolation, cDNA Synthesis, and RACE Amplification
2.3. Sequence and Phylogenetic Analysis
2.4. Preparation of Non-Diapause, Summer, and Winter Diapause Pupae
2.5. Cryogenic and Heat Shock Treatment on P. melete Pupae
2.6. Quantitative Real-Time PCR (qRT-PCR)
2.7. Statistical Analysis
3. Results
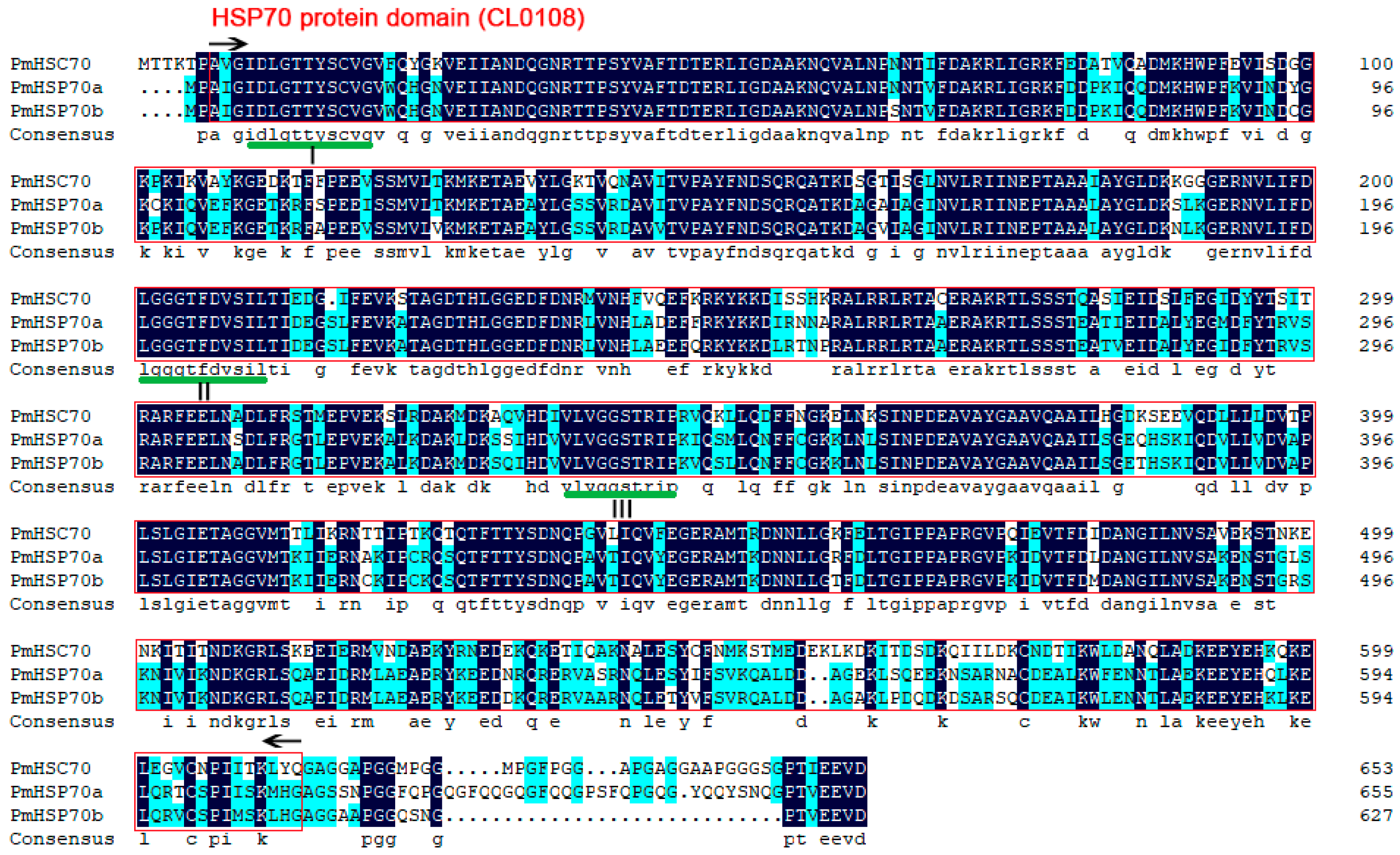
3.1. Sequence Analysis of PmHsc70, PmHsp70a, and PmHsp70b
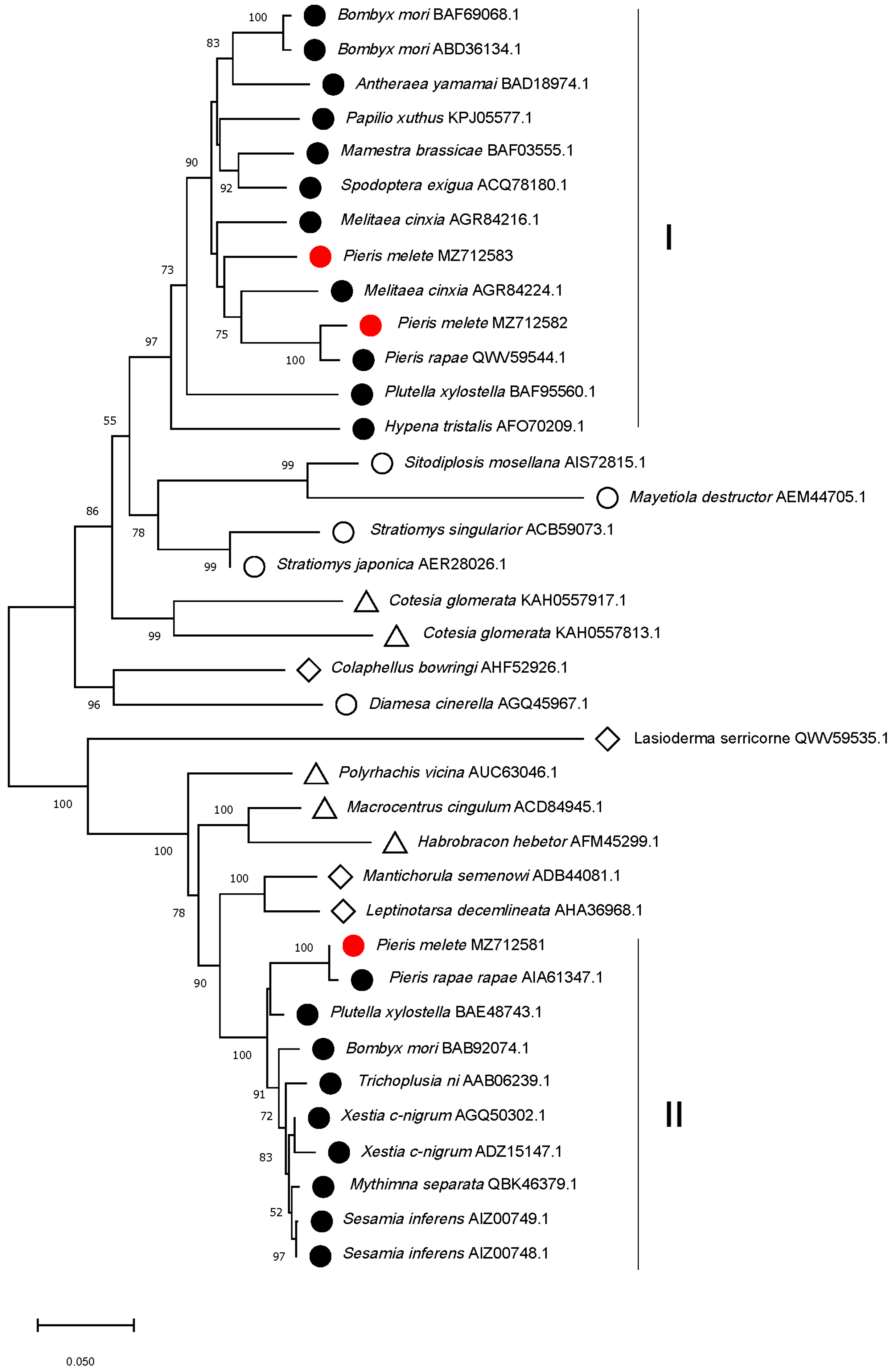
3.2. Sequence Alignment and Phylogenetic Relationship Analysis
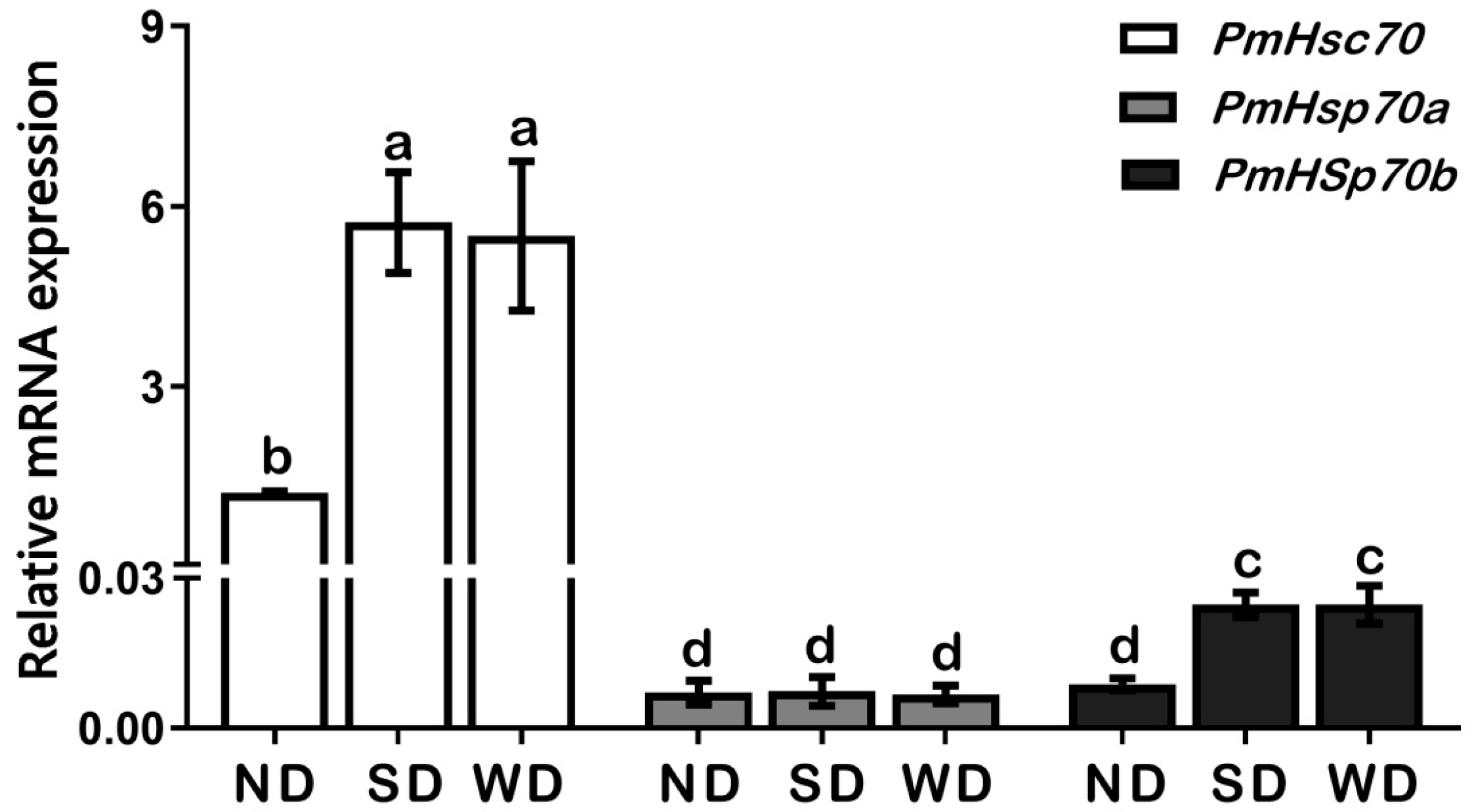
3.3. Expression Patterns of PmHsc70, PmHsp70a, and PmHsp70b in Pupae of ND, SD, and WD Pupae
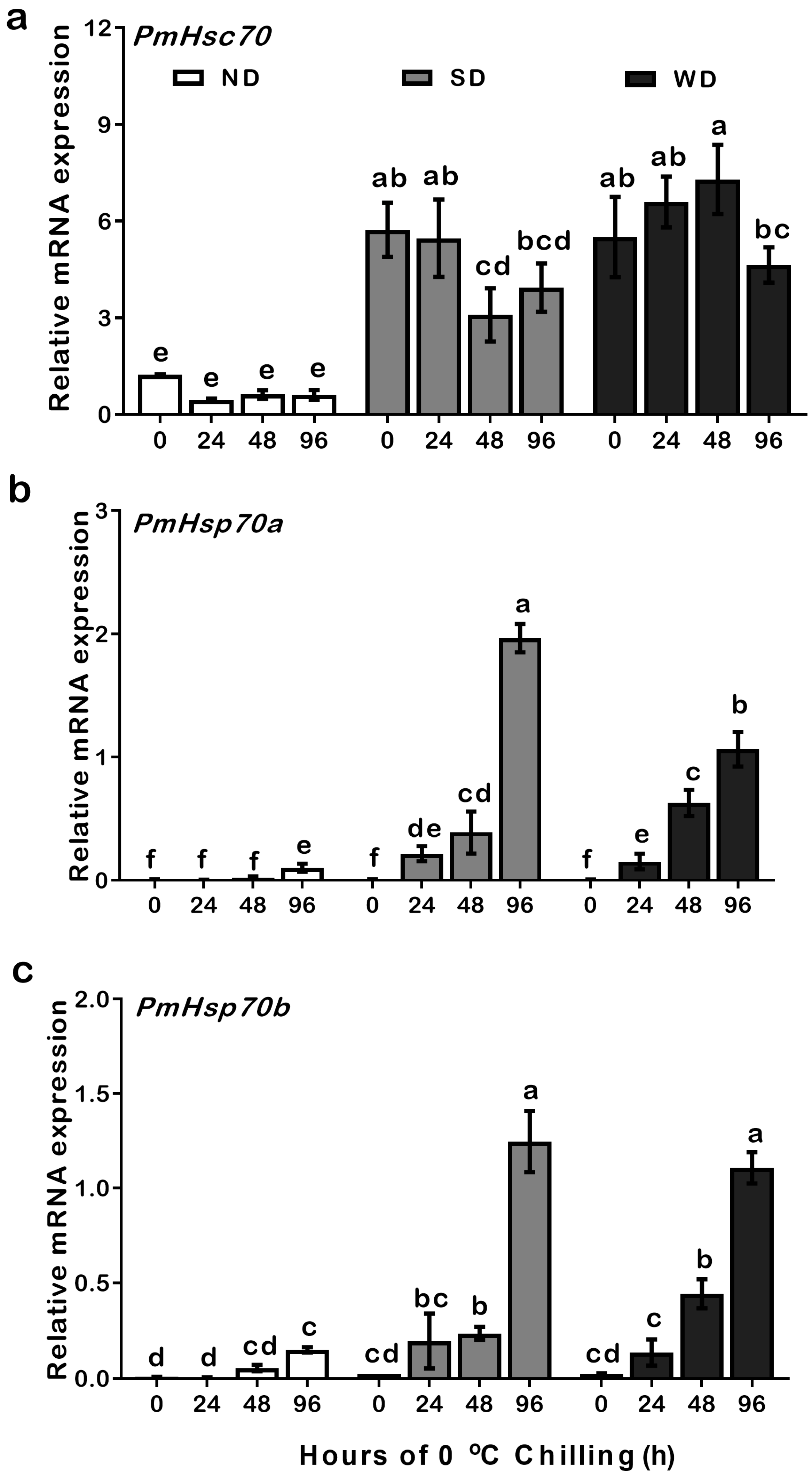
3.4. Expression Patterns of PmHsc70, PmHsp70a, and PmHsp70b in ND, SD, and WD Pupae at 0 °C Cryogenic Chilling
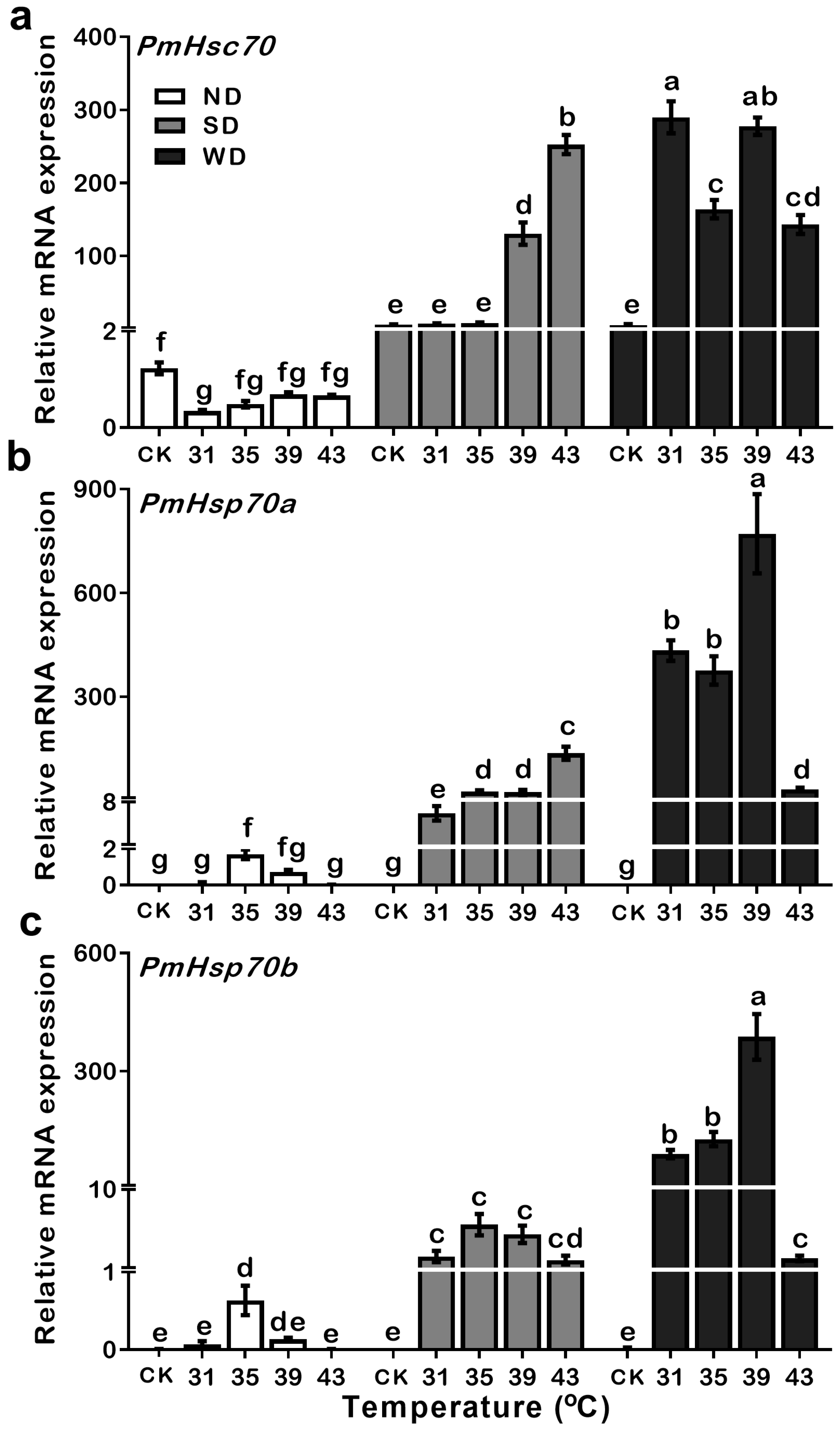
3.5. Expression Patterns of PmHsc70, PmHsp70a, and PmHsp70b in ND, SD and WD Pupae during Heat Treatments
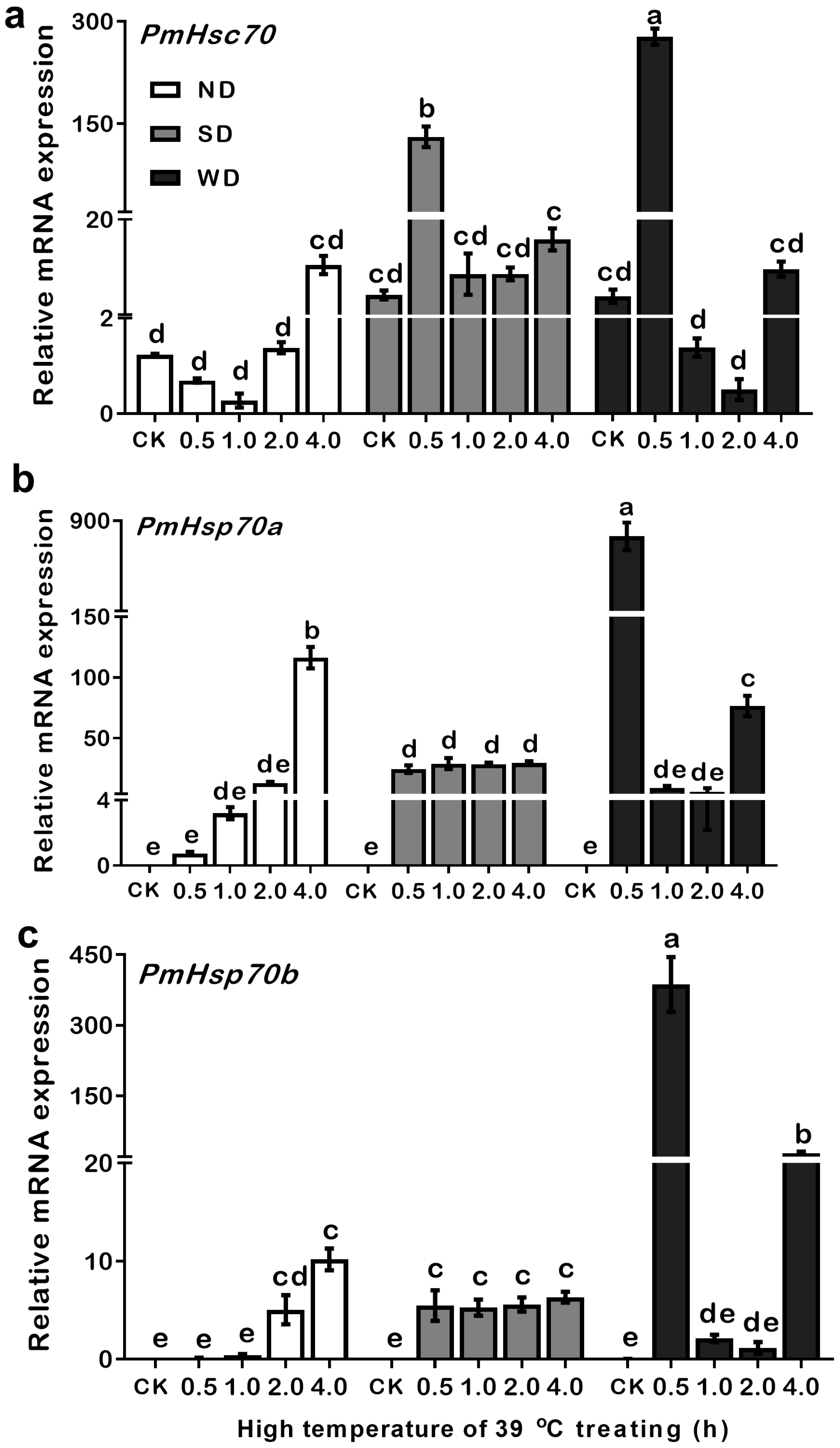
3.6. Expression Patterns of PmHsc70, PmHsp70a, and PmHsp70b in ND, SD, and WD Pupae at 39 °C
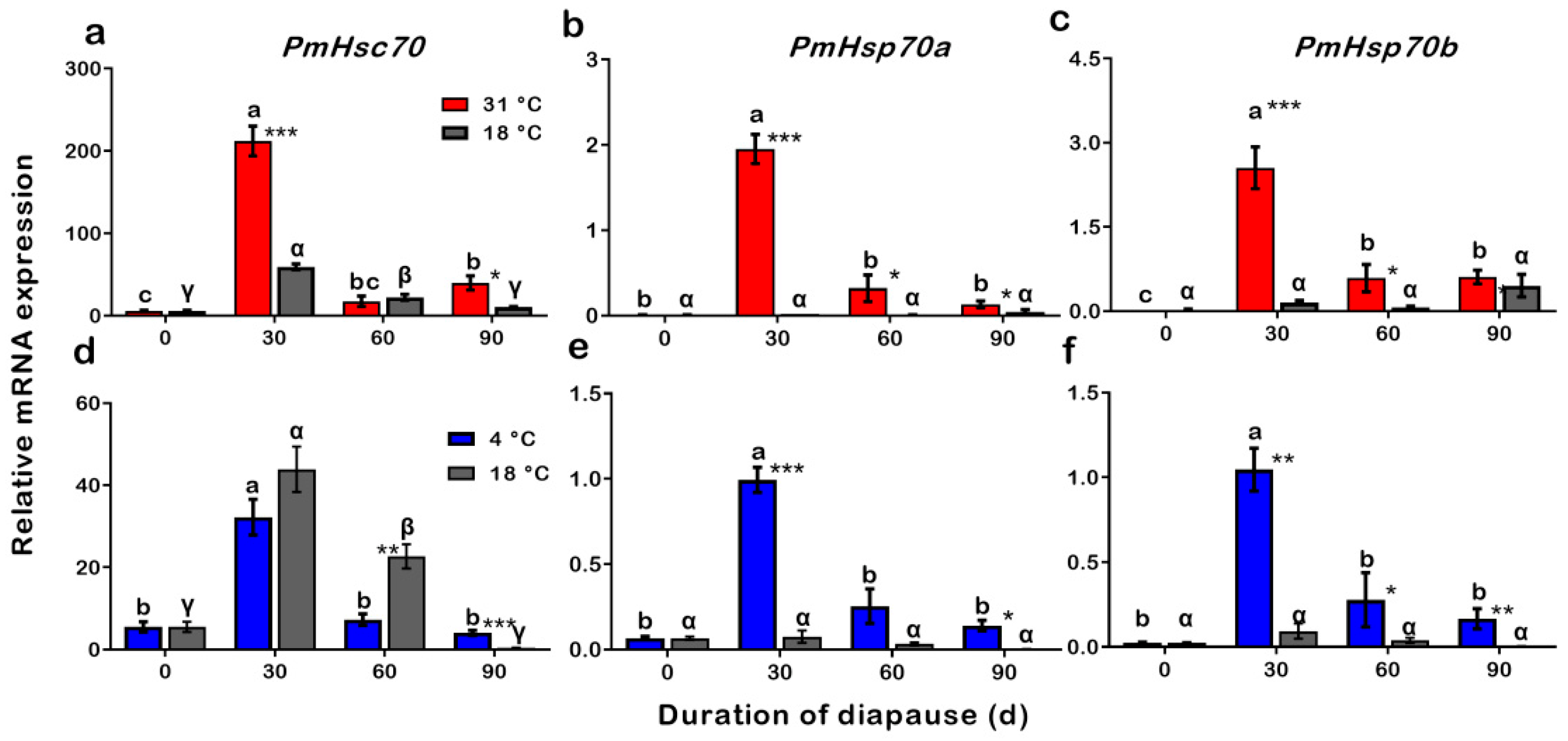
3.7. Expression Patterns of PmHsc70, PmHsp70a, and PmHsp70b during Procedures of Summer and Winter Diapause Development
4. Discussion
4.1. Expressions of PmHsc70, PmHsp70a, and PmHsp70b Related to Diapause and Cold Stress
4.2. Expressions of PmHsc70, PmHsp70a, and PmHsp70b Related to Heat Shock
4.3. Expressions of PmHsc70, PmHsp70a, and PmHsp70b during Summer and Winter Diapause
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Clarke, A. Costs and consequences of evolutionary temperature adaptation. Trends Ecol. Evol. 2003, 18, 573–581. [Google Scholar] [CrossRef]
- Zhang, Q.; Denlinger, D.L. Molecular characterization of heat shock protein 90, 70 and 70 cognate cDNAs and their expression patterns during thermal stress and pupal diapause in the corn earworm. J. Insect Physiol. 2010, 56, 138–150. [Google Scholar] [CrossRef] [PubMed]
- Zhao, L.; Jones, W.A. Expression of heat shock protein genes in insect stress responses. Invertebr. Surviv. J. 2012, 9, 93–101. [Google Scholar]
- King, A.M.; MacRae, T.H. Insect heat shock proteins during stress and diapause. Annu. Rev. Entomol. 2015, 60, 59–75. [Google Scholar] [CrossRef] [PubMed]
- Morimoto, R.I. Regulation of the heat shock transcriptional response: Cross talk between a family of heat shock factors, molecular chaperones, and negative regulators. Gene. Dev. 1998, 12, 3788–3796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calabria, G.; Dolgova, O.; Rego, C.; Castañeda, L.E.; Rezende, E.L.; Balanyà, J.; Pascual, M.; Sørensen, J.G.; Loeschcke, V.; Santos, M. Hsp70 protein levels and thermotolerance in Drosophila subobscura: A reassessment of the thermal co-adaptation hypothesis. J. Evolution. Biol. 2012, 25, 691–700. [Google Scholar] [CrossRef]
- Kiang, J.G.; Tsokos, G.C. Heat shock protein 70 kDa: Molecular biology, biochemistry, and physiology. Pharmacol. Therapeut. 1998, 80, 183–201. [Google Scholar] [CrossRef]
- Li, H.; Qiao, H.; Liu, Y.; Li, S.; Tan, J.; Hao, D. Characterization, expression profiling, and thermal tolerance analysis of heat shock protein 70 in pine sawyer beetle, Monochamus alternatus hope (Coleoptera: Cerambycidae). Bull. Entomol. Res. 2021, 111, 217–228. [Google Scholar] [CrossRef]
- Xiang, L.; Kang, Z.; Yu, X.; Fan, L.I.; Liu, T.; Qiang, L.I. Role of TRP channels and HSPs in thermal stress response in the aphid parasitoid Aphelinus asychis (Hymenoptera: Aphelinidae). J. Integr. Agric. 2020, 19, 1530–1542. [Google Scholar]
- Rinehart, J.P.; Hayward, S.A.; Elnitsky, M.A.; Sandro, L.H.; Lee, R.E.; Denlinger, D.L. Continuous up-regulation of heat shock proteins in larvae, but not adults, of a polar insect. Proc. Natl. Acad. Sci. USA 2006, 103, 14223–14227. [Google Scholar] [CrossRef] [Green Version]
- PAN, D.; CAO, S.; LU, M.; HANG, S.; DU, Y. Genes encoding heat shock proteins in the endoparasitoid wasp, Cotesia chilonis, and their expression in response to temperatures. J. Integr. Agric. 2018, 17, 1012–1022. [Google Scholar] [CrossRef]
- Chang, Y.; Zhang, X.; Chen, J.; Lu, M.; Gong, W.; Du, Y. Characterization of three heat shock protein 70 genes from Liriomyza trifolii and expression during thermal stress and insect development. Bull. Entomol. Res. 2019, 109, 150–159. [Google Scholar] [CrossRef]
- Shu, Y.; Du, Y.; Wang, J. Molecular characterization and expression patterns of Spodoptera litura heat shock protein 70/90, and their response to zinc stress. Comp. Biochem. Phys. A. 2011, 158, 102–110. [Google Scholar] [CrossRef]
- Paim, R.M.; Araujo, R.N.; Leis, M.; Sant’Anna, M.R.; Gontijo, N.F.; Lazzari, C.R.; Pereira, M.H. Functional evaluation of Heat Shock Proteins 70 (HSP70/HSC70) on Rhodnius prolixus (Hemiptera, Reduviidae) physiological responses associated with feeding and starvation. Insect Biochem. Mol. Biol. 2016, 77, 10–20. [Google Scholar] [CrossRef]
- Zheng, W.; Yang, D.; Wang, J.; Song, Q.; Gilbert, L.I.; Zhao, X. Hsc70 binds to ultraspiracle resulting in the upregulation of 20-hydroxyecdsone-responsive genes in Helicoverpa armigera. Mol. Cell. Endocrinol. 2010, 315, 282–291. [Google Scholar] [CrossRef]
- Denlinger, D.L. Regulation of diapause. Annu. Rev. Entomol. 2002, 47, 93–122. [Google Scholar] [CrossRef]
- MacRae, T.H. Gene expression, metabolic regulation and stress tolerance during diapause. Cell. Mol. Life Sci. 2010, 67, 2405–2424. [Google Scholar] [CrossRef]
- Goto, S.G.; Yoshida, K.M.; Kimura, M.T. Accumulation of Hsp70 mRNA under environmental stresses in diapausing and nondiapausing adults of Drosophila triauraria. J. Insect Physiol. 1998, 44, 1009–1015. [Google Scholar] [CrossRef]
- Aruda, A.M.; Baumgartner, M.F.; Reitzel, A.M.; Tarrant, A.M. Heat shock protein expression during stress and diapause in the marine copepod Calanus finmarchicus. J. Insect Physiol. 2011, 57, 665–675. [Google Scholar] [CrossRef] [Green Version]
- Sonoda, S.; Fukumoto, K.; Izumi, Y.; Yoshida, H.; Tsumuki, H. Cloning of heat shock protein genes (hsp90 and hsc70) and their expression during larval diapause and cold tolerance acquisition in the rice stem borer, Chilo suppressalis Walker. Arch. Insect Biochem. 2006, 63, 36–47. [Google Scholar] [CrossRef]
- Hayward, S.; Pavlides, S.C.; Tammariello, S.P.; Rinehart, J.P.; Denlinger, D.L. Temporal expression patterns of diapause-associated genes in flesh fly pupae from the onset of diapause through post-diapause quiescence. J. Insect Physiol. 2005, 51, 631–640. [Google Scholar] [CrossRef]
- Xiao, H.; Li, F.; Wei, X.; Xue, F. A comparison of photoperiodic control of diapause between aestivation and hibernation in the cabbage butterfly Pieris melete. J. Insect Physiol. 2008, 54, 755–764. [Google Scholar] [CrossRef]
- Xiao, H.J.; Wu, S.H.; He, H.M.; Chen, C.; Xue, F.S. Role of natural day-length and temperature in determination of summer and winter diapause in Pieris melete (Lepidoptera: Pieridae). Bull. Entomol. Res. 2012, 102, 267–273. [Google Scholar] [CrossRef]
- Xiao, H.; Wu, S.; Chen, C.; Xue, F. Optimal low temperature and chilling period for both summer and winter diapause development in Pieris melete: Based on a similar mechanism. PLoS ONE 2013, 8, e56404. [Google Scholar] [CrossRef]
- Wu, Y.K.; Zou, C.; Fu, D.M.; Zhang, W.N.; Xiao, H.J. Molecular characterization of three Hsp90 from Pieris and expression patterns in response to cold and thermal stress in summer and winter diapause of Pieris melete. Insect Sci. 2018, 25, 273–283. [Google Scholar] [CrossRef]
- Jiang, T.; Zhu, Y.; Peng, Y.; Zhang, W.; Xiao, H. Universal and differential transcriptional regulatory pathways involved in the preparation of summer and winter diapauses in Pieris melete. Bull. Entomol. Res. 2021, 111, 371–378. [Google Scholar] [CrossRef]
- Sørensen, J.G.; Giribets, M.P.; Tarrío, R.; Rodríguez-Trelles, F.; Schou, M.F.; Loeschcke, V. Expression of thermal tolerance genes in two Drosophila species with different acclimation capacities. J. Therm. Biol. 2019, 84, 200–207. [Google Scholar] [CrossRef]
- Rinehart, J.P.; Li, A.; Yocum, G.D.; Robich, R.M.; Hayward, S.A.; Denlinger, D.L. Up-regulation of heat shock proteins is essential for cold survival during insect diapause. Proc. Natl. Acad. Sci. USA 2007, 104, 11130–11137. [Google Scholar] [CrossRef] [Green Version]
- Yocum, G.D. Differential expression of two HSP70 transcripts in response to cold shock, thermoperiod, and adult diapause in the Colorado potato beetle. J. Insect Physiol. 2001, 47, 1139–1145. [Google Scholar] [CrossRef]
- Colinet, H.; Lee, S.F.; Hoffmann, A. Temporal expression of heat shock genes during cold stress and recovery from chill coma in adult Drosophila melanogaster. FEBS J. 2010, 277, 174–185. [Google Scholar] [CrossRef]
- Chen, B.; Kayukawa, T.; Monteiro, A.; Ishikawa, Y. Cloning and characterization of the HSP70 gene, and its expression in response to diapauses and thermal stress in the onion maggot, Delia antiqua. BMB Rep. 2006, 39, 749–758. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, L.J.; Wang, K.F.; Jing, Y.P.; Zhuang, H.M.; Wu, G. Identification of heat shock protein genes hsp70s and hsc70 and their associated mRNA expression under heat stress in insecticide-resistant and susceptible diamondback moth, Plutella xylostella (Lepidoptera: Plutellidae). Eur. J. Entomol. 2015, 112, 215. [Google Scholar] [CrossRef]
- Qin, W.; Tyshenko, M.G.; Wu, B.S.; Walker, V.K.; Robertson, R.M. Cloning and characterization of a member of the hsp70 gene family from Locusta migratoria, a highly thermotolerant insect. Cell Stress Chaperon. 2003, 8, 144. [Google Scholar] [CrossRef] [Green Version]
- Sun, Y.; Zhao, J.; Sheng, Y.; Xiao, Y.F.; Zhang, Y.J.; Bai, L.X.; Tan, Y.; Xiao, L.B.; Xu, G.C. Identification of heat shock cognate protein 70 gene (Alhsc70) of Apolygus lucorum and its expression in response to different temperature and pesticide stresses. Insect Sci. 2016, 23, 37–49. [Google Scholar] [CrossRef]
- Fang, S.; Zhang, Q.; Zhang, Y.; Zhang, G.; Zhang, Z.; Yu, Q. Heat Shock Protein 70 Family in Response to Multiple Abiotic Stresses in the Silkworm. Insects 2021, 12, 928. [Google Scholar] [CrossRef]
- Sørensen, J.G.; Michalak, P.; Justesen, J.; Loeschcke, V. Expression of the heat-shock protein HSP70 in Drosophila buzzatii lines selected for thermal resistance. Hereditas 1999, 131, 155–164. [Google Scholar] [CrossRef]
- Rank, N.E.; Bruce, D.A.; McMillan, D.M.; Barclay, C.; Dahlhoff, E.P. Phosphoglucose isomerase genotype affects running speed and heat shock protein expression after exposure to extreme temperatures in a montane willow beetle. J. Exp. Biol. 2007, 210, 750–764. [Google Scholar] [CrossRef] [Green Version]
- Lu, K.; Chen, X.; Liu, W.; Zhang, Z.; Wang, Y.; You, K.; Li, Y.; Zhang, R.; Zhou, Q. Characterization of heat shock protein 70 transcript from Nilaparvata lugens (Stål): Its response to temperature and insecticide stresses. Pestic. Biochem. Phys. 2017, 142, 102–110. [Google Scholar] [CrossRef]
- Jing, Q.; Peng, G.; Zhang, X.; LU, M.; DU, Y. Characterization of two novel heat shock protein 70s and their transcriptional expression patterns in response to thermal stress in adult of Frankliniella occidentalis (Thysanoptera: Thripidae). J. Integr. Agric. 2018, 17, 1023–1031. [Google Scholar]
- Wang, X.R.; Wang, C.; Ban, F.X.; Zhu, D.T.; Liu, S.S.; Wang, X.W. Genome-wide identification and characterization of HSP gene superfamily in whitefly (Bemisia tabaci) and expression profiling analysis under temperature stress. Insect Sci. 2019, 26, 44–57. [Google Scholar] [CrossRef] [Green Version]
- Silbermann, R.; Tatar, M. Reproductive costs of heat shock protein in transgenic Drosophila melanogaster. Evolution 2000, 54, 2038–2045. [Google Scholar] [CrossRef]
- Wang, H.; Dong, S.; Li, K.; Hu, C.; Ye, G. A heat shock cognate 70 gene in the endoparasitoid, Pteromalus puparum, and its expression in relation to thermal stress. BMB Rep. 2008, 41, 388–393. [Google Scholar] [CrossRef] [Green Version]
- Tian, C.; Li, Y.; Wu, Y.; Chu, W.; Liu, H. Sustaining induced heat shock protein 70 confers biological thermotolerance in a high-temperature adapted predatory mite Neoseiulus barkeri (Hughes). Pest Manag. Sci. 2021, 77, 939–948. [Google Scholar] [CrossRef]
- Podrabsky, J.E.; Lopez, J.P.; Fan, T.W.; Higashi, R.; Somero, G.N. Extreme anoxia tolerance in embryos of the annual killifish Austrofundulus limnaeus: Insights from a metabolomics analysis. J. Exp. Biol. 2007, 210, 2253–2266. [Google Scholar] [CrossRef] [Green Version]
- Cheng, W.; Li, D.; Wang, Y.; Liu, Y.; Zhu-Salzman, K. Cloning of heat shock protein genes (hsp70, hsc70 and hsp90) and their expression in response to larval diapause and thermal stress in the wheat blossom midge, Sitodiplosis mosellana. J. Insect Physiol. 2016, 95, 66–77. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, J.; Miano, F.N.; Jiang, T.; Peng, Y.; Zhang, W.; Xiao, H. Characterization of Three Heat Shock Protein Genes in Pieris melete and Their Expression Patterns in Response to Temperature Stress and Pupal Diapause. Insects 2022, 13, 430. https://doi.org/10.3390/insects13050430
Zhang J, Miano FN, Jiang T, Peng Y, Zhang W, Xiao H. Characterization of Three Heat Shock Protein Genes in Pieris melete and Their Expression Patterns in Response to Temperature Stress and Pupal Diapause. Insects. 2022; 13(5):430. https://doi.org/10.3390/insects13050430
Chicago/Turabian StyleZhang, Jing, Falak Naz Miano, Ting Jiang, Yingchuan Peng, Wanna Zhang, and Haijun Xiao. 2022. "Characterization of Three Heat Shock Protein Genes in Pieris melete and Their Expression Patterns in Response to Temperature Stress and Pupal Diapause" Insects 13, no. 5: 430. https://doi.org/10.3390/insects13050430