
New Cretaceous Lacewings in a Transitional Lineage of Myrmeleontoidea and Their Phylogenetic Implications †


{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Method
2.1. Taxonomy
2.2. Phylogenetic Analysis
3. Results
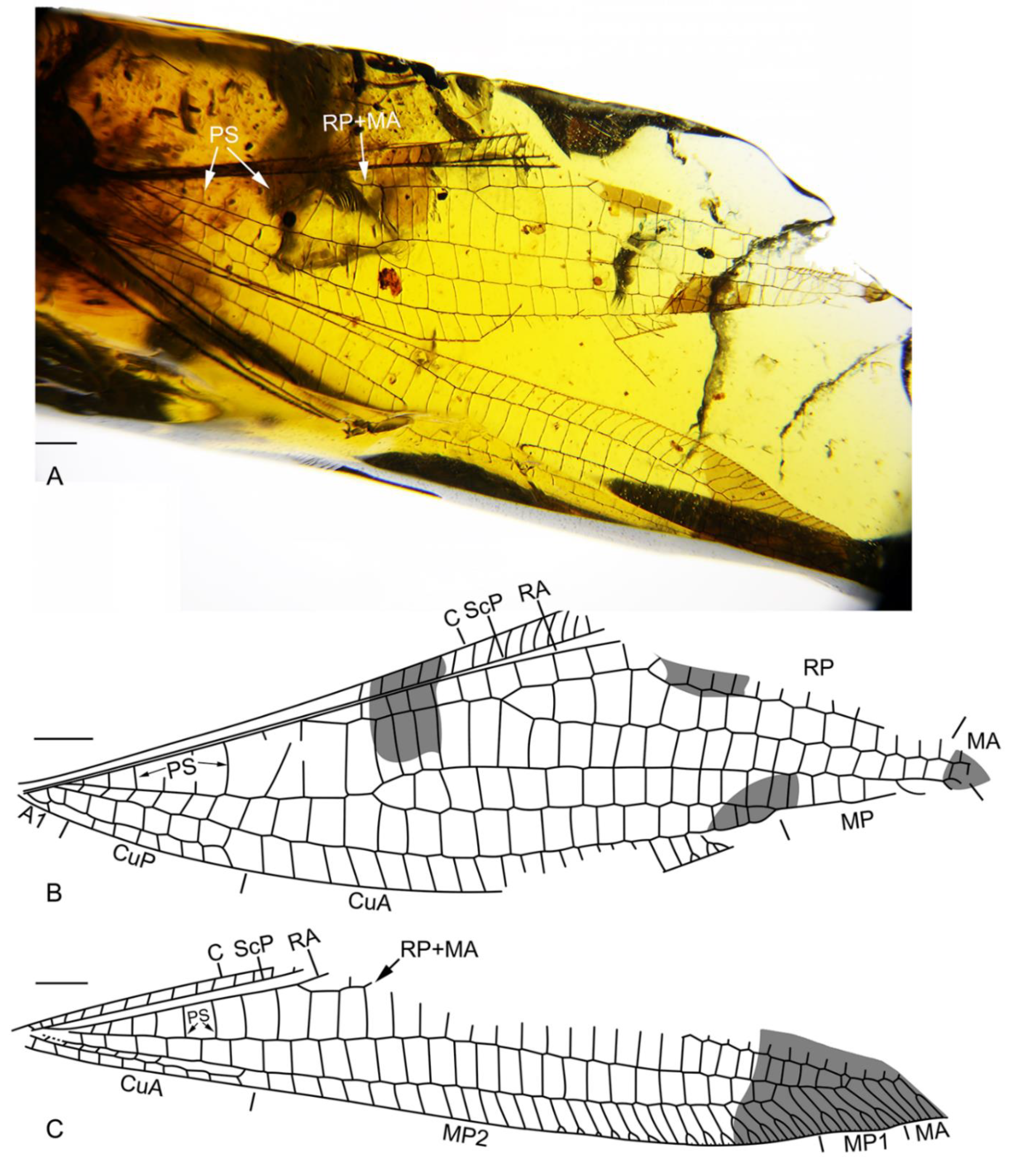
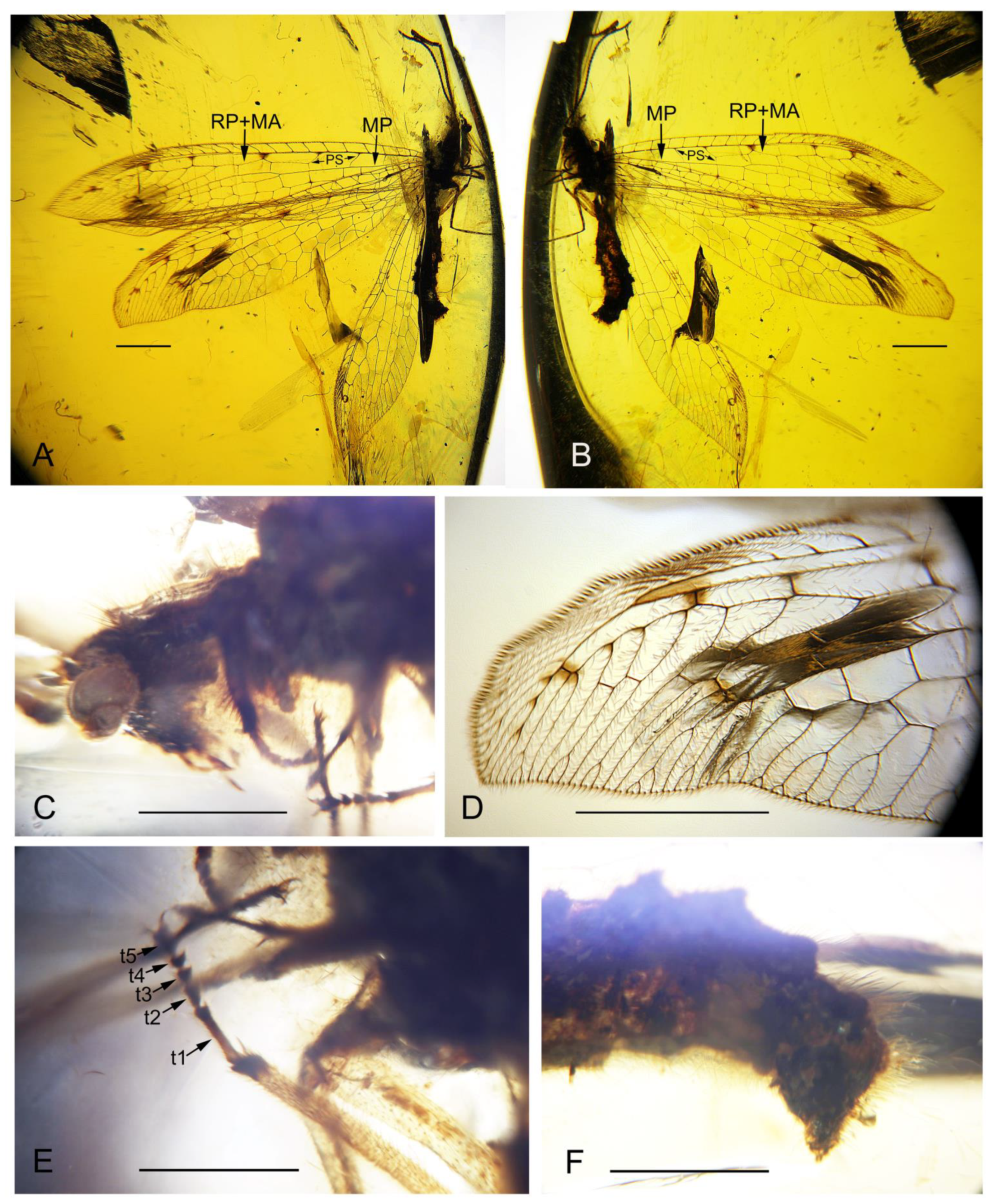
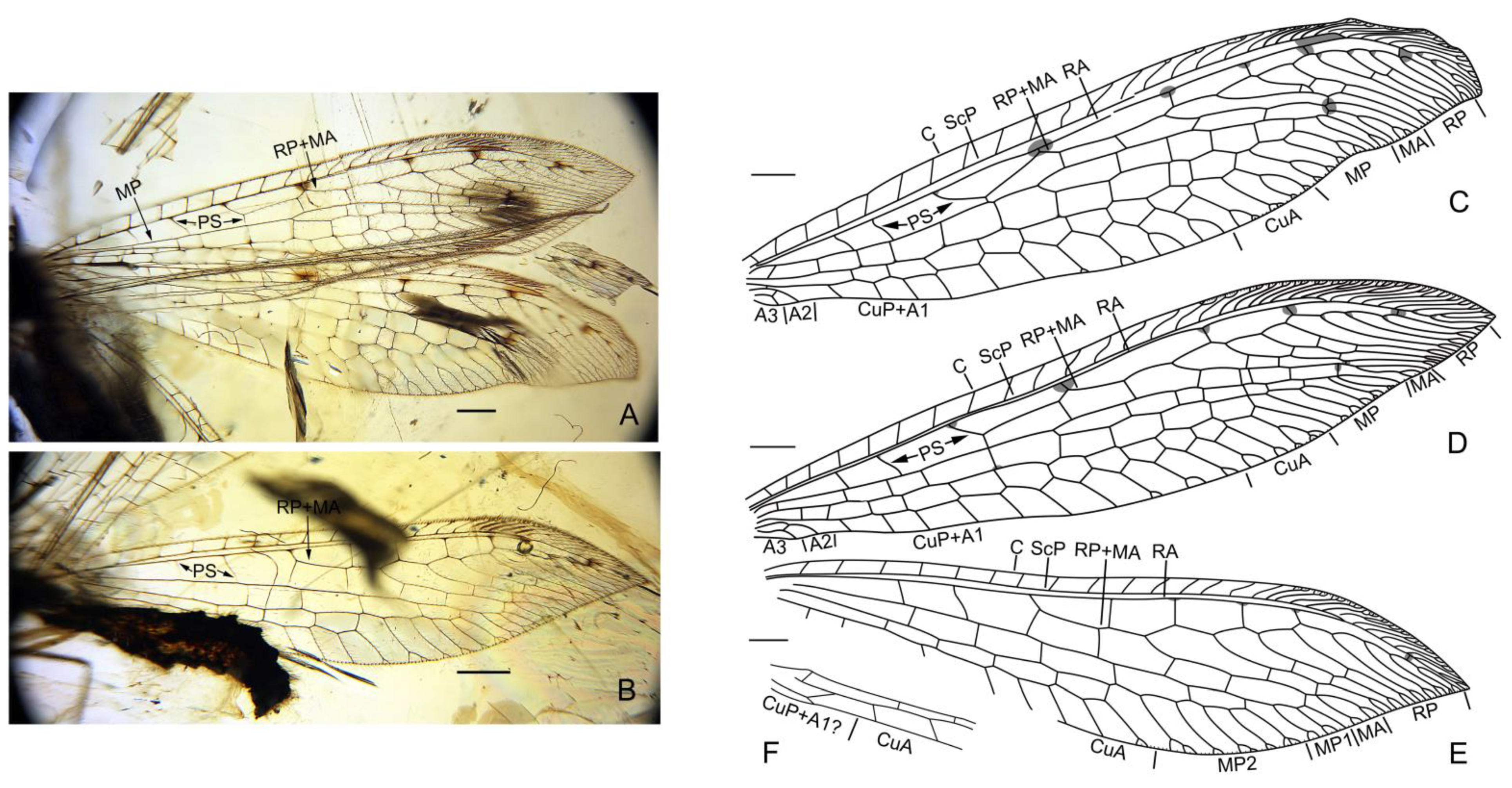
3.1. Systematic Paleontology



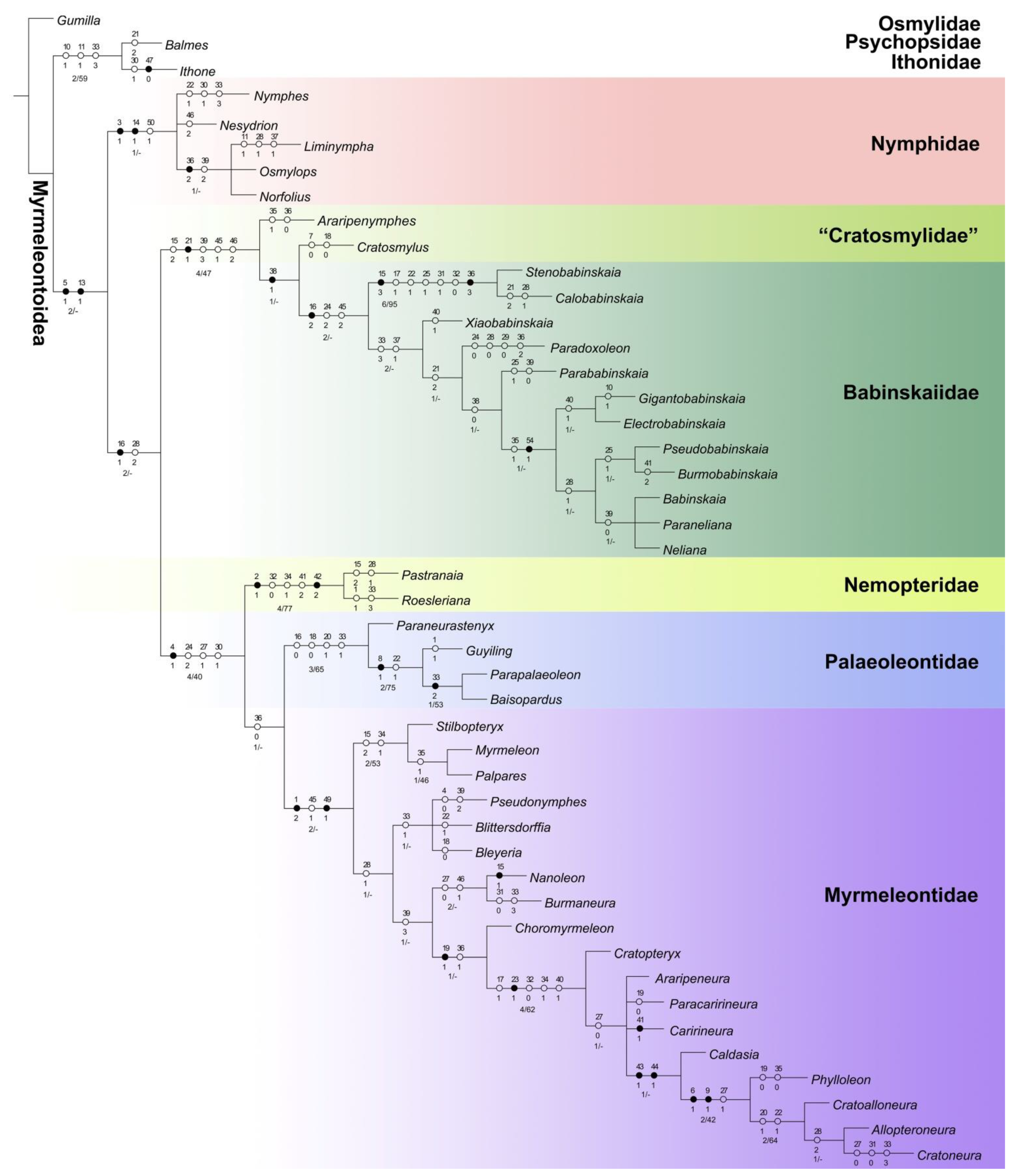
3.2. Phylogenetic Analysis
4. Discussion
4.1. Familial Status of Cratosmylidae
4.2. New Characters of Babinskaiidae
4.3. Biogeographical Consideration
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Engel, M.S.; Winterton, S.L.; Breitkreuz, L.C.V. Phylogeny and evolution of Neuropterida: Where have wings of lace taken us? Annu. Rev. Entomol. 2018, 63, 531–551. [Google Scholar] [CrossRef] [PubMed]
- Machado, R.J.P.; Gillung, J.P.; Winterton, S.L.; Garzón-Orduña, I.J.; Lemmon, A.R.; Lemmon, E.M.; Oswald, J.D. Owlflies are derived antlions: Anchored phylogenomics supports a new phylogeny and classification of Myrmeleontidae (Neuroptera). Syst. Entomol. 2019, 44, 418–450. [Google Scholar] [CrossRef]
- Oswald, J.D. Bibliography of the Neuropterida. Lacewing Digital Library, Research Publication No. 2. Available online: http://lacewing.tamu.edu/Biblio/Main (accessed on 4 March 2022).
- Oswald, J.D. Neuropterida Species of the World. Lacewing Digital Library, Research Publication No. 1. Available online: https://lacewing.tamu.edu/SpeciesCatalog/Main (accessed on 4 March 2022).
- New, T.R. A revision of the Australian Nymphidae (Insecta: Neuroptera). Aust. J. Zool. 1981, 29, 707–750. [Google Scholar] [CrossRef]
- Tjeder, B. Neuroptera-Planipennia. The Lace-wings of Southern Africa. 6. Family Nemopteridae. S. Afr. Anim. Life 1967, 13, 290–501. [Google Scholar]
- Badano, D.; Engel, M.S.; Basso, A.; Wang, B.; Cerretti, P. Diverse Cretaceous larvae reveal the evolutionary and behavioural history of antlions and lacewings. Nat. Commu. 2018, 9, 3257. [Google Scholar] [CrossRef]
- Ren, D.; Guo, Z.G. On the new fossil genera and species of Neuroptera (Insecta) from the Late Jurassic of northeast China. Acta Zootaxon. Sin. 1996, 21, 461–479. [Google Scholar]
- Ren, D.; Engel, M.S. A second antlion from the Mesozoic of northeastern China (Neuroptera: Myrmeleontidae). Alavesia 2008, 2, 183–186. [Google Scholar]
- Winterton, S.L.; Hardy, N.B.; Wiegmann, B.M. On wings of lace: Phylogeny and Bayesian divergence time estimates of Neuropterida (Insecta) based on morphological and molecular data. Syst. Entomol. 2010, 35, 349–378. [Google Scholar] [CrossRef]
- Wang, Y.Y.; Liu, X.Y.; Garzón-Orduña, I.J.; Winterton, S.L.; Yan, Y.; Aspöck, U.; Aspöck, H.; Yang, D. Mitochondrial phylogenomics illuminates the evolutionary history of Neuroptera. Cladistics 2017, 33, 617–636. [Google Scholar] [CrossRef]
- Martins-Neto, R.G.; Vulcano, M.A. Neurópteros (Insecta, Planipennia) da Formação Santana (Cretáceo Inferior), Bacia do Araripe, Nordeste do Brasil: VIII—Descrição de novas taxas de Myrmeleontidae, Ascalaphidae e Nemopteridae. Rev. Uni. Guarulhos Série Ciênc. Biol. Saúde 1997, 2, 64–81. [Google Scholar]
- Menon, F.; Martins-Neto, R.G.; Martill, D.M. A new Lower Cretaceous nymphid (Insecta, Neuroptera, Nymphidae) from the Crato Formation of Brazil. GAEA 2005, 1, 11–15. [Google Scholar]
- Shi, C.F.; Winterton, S.L.; Ren, D. Phylogeny of split-footed lacewings (Neuroptera, Nymphidae), with descriptions of new Cretaceous fossil species from China. Cladistics 2015, 31, 455–490. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.M.; Hu, J.H.; Wang, B.; Zhang, W.W.; Ohl, M.; Liu, X.Y. New antlions (Insecta: Neuroptera: Myrmeleontidae) from the mid-Cretaceous of Myanmar and their phylogenetic implications. J. Syst. Palaeontol. 2019, 17, 995–1012. [Google Scholar] [CrossRef]
- Lu, X.M.; Wang, B.; Yang, S.H.; Liu, X.Y. Early evolution of Nemopteridae illuminated with the first and oldest thread-winged lacewing in Cretaceous amber. Syst. Entomol. 2019, 44, 262–272. [Google Scholar] [CrossRef]
- Lu, X.M.; Liu, X.Y. The Neuropterida from the mid-Cretaceous of Myanmar: A spectacular palaeodiversity bridging the Mesozoic and present faunas. Cret. Res. 2021, 121, 104727. [Google Scholar] [CrossRef]
- Lu, X.M.; Wang, B.; Liu, X.Y. New Cretaceous antlion-like lacewings promote a phylogenetic reappraisal of the extinct myrmeleontoid family Babinskaiidae. Sci. Rep. 2021, 11, 16431. [Google Scholar] [CrossRef]
- Martins-Neto, R.G.; Vulcano, M.A. Neurópteros (Insecta, Planipennia) da Formaçao Santana (Cretáceo Inferior), bacia do Araripe, nordeste do Brasil. II. Superfamília Myrmeleontoidea. Rev. Bras. Entomol. 1989, 33, 367–402. [Google Scholar]
- Martins-Neto, R.G.; Vulcano, M.A. Neurópteros (Insecta, Planipennia) da Formaçao Santana (Cretáceo Inferior), Bacia do Araripe, nordeste do Brasil. IV—Complemento às partes I e II, com descriçao de novos taxa. An. Acad. Bras. Cienc. 1989, 61, 311–318. [Google Scholar]
- Ponomarenko, A.G. Neuroptera (Insecta) from the Lower Cretaceous of Transbaikalia. Paleontol. J. 1992, 1992, 43–50. (In Russian) [Google Scholar]
- Martins Neto, R.G. Neurópteros (Insecta, Planipennia) da Formaçao Santana (Cretáceo Inferior), Bacia do Araripe, Nordeste do Brasil. X—Descriçao de novos táxons (Chrysopidae, Babinskaiidae, Myrmeleontidae, Ascalaphidae e Psychopsidae). Rev. Univ. Guarulhos Série Ciênc. Exat. Technol. 1997, 2, 68–83. [Google Scholar]
- Makarkin, V.N.; Heads, S.W.; Wedmann, S. Taxonomic study of the Cretaceous lacewing family Babinskaiidae (Neuroptera: Myrmeleontoidea: Nymphidoidae), with description of new taxa. Cret. Res. 2017, 78, 149–160. [Google Scholar] [CrossRef]
- Hu, J.H.; Lu, X.M.; Wang, B.; Liu, X.Y. Taxonomic notes on Babinskaiidae from the Cretaceous Burmese amber, with the description of a new species (Insecta, Neuroptera). ZooKeys 2018, 748, 31–46. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.M.; Zhang, W.W.; Liu, X.Y. Discovery of the family Babinskaiidae (Insecta: Neuroptera) in mid-Cretaceous amber from Myanmar. Cret. Res. 2017, 71, 14–23. [Google Scholar] [CrossRef]
- Myskowiak, J.; Escuillié, F.; Nel, A. A new Osmylidae (Insecta, Neuroptera) from the Lower Cretaceous Crato Formation in Brazil. Cret. Res. 2015, 54, 27–33. [Google Scholar] [CrossRef]
- Yu, T.T.; Kelly, R.; Mu, L.; Ross, A.; Kennedy, J.; Broly, P.; Xia, F.Y.; Zhang, H.C.; Wang, B.; Dilcher, D. An ammonite trapped in Burmese amber. Proc. Natl. Acad. Sci. USA 2019, 116, 11345–11350. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shi, G.H.; Grimaldi, D.; Harlow, G.E.; Wang, J. Age constraint on Burmese amber based on U-Pb dating of zircons. Cret. Res. 2012, 37, 155–163. [Google Scholar] [CrossRef]
- Aspöck, H.; Aspöck, U.; Hölzel, H. Die Neuropteren Europas (2 Vols); Goecke and Evers: Krefeld, Germany, 1980; pp. 355, 495. [Google Scholar]
- Martins-Neto, R.G. Remarks on the neuropterofauna (Insecta, Neuroptera) from the Brazilian Cretaceous, with keys for the identification of the known taxa. Acta Geol. Hispan. 2000, 35, 97–118. [Google Scholar]
- Aspöck, U.; Aspöck, H. Phylogenetic relevance of the genital sclerites of Neuropterida (Insecta: Holometabola). Syst. Entomol. 2008, 33, 97–127. [Google Scholar] [CrossRef]
- Goloboff, P.A.; Catalano, S.A. TNT version 1.5, including a full implementation of phylogenetic morphometrics. Cladistics 2016, 3, 221–238. [Google Scholar] [CrossRef]
- Nixon, K.C. WinClada; ver. 1.00.08; Published by the author: Ithaca, NY, USA, 2002. [Google Scholar]
- Myskowiak, J.; Huang, D.Y.; Azar, D.; Cai, C.Y.; Garrouste, R.; Nel, A. New lacewings (Insecta, Neuroptera, Osmylidae, Nymphidae) from the Lower Cretaceous Burmese amber and Crato Formation in Brazil. Cret. Res. 2016, 59, 214–227. [Google Scholar] [CrossRef]
- Makarkin, V.N.; Wedmann, S.; Heads, S.W. A systematic reappraisal of Araripeneuridae (Neuroptera: Myrmeleontoidea), with description of new species from the Lower Cretaceous Crato Formation of Brazil. Cret. Res. 2018, 84, 600–621. [Google Scholar] [CrossRef]
- Winterton, S.L.; Califre Martins, C.; Makarkin, V.N.; Ardila Camacho, J.A.; Wang, Y.J. Lance lacewings of the world (Neuroptera: Archeosmylidae, Osmylidae, Saucrosmylidae): Review of living and fossil genera. Zootaxa 2019, 4581, 1–99. [Google Scholar] [CrossRef] [PubMed]
- Makarkin, V.N.; Staniczek, A.H. A new large-sized genus of Babinskaiidae (Neuroptera: Myrmeleontoidea: Nymphidoidae) from mid-Cretaceous Burmese amber. Cret. Res. 2019, 104, 104196. [Google Scholar] [CrossRef]
- Breitkreuz, L.C.V.; Winterton, S.L.; Engel, M.S. Wing tracheation in Chrysopidae and other Neuropterida (Insecta): A resolution of the confusion about vein fusion. Am. Mus. Novit. 2017, 3890, 1–44. [Google Scholar] [CrossRef] [Green Version]
- Martins-Neto, R.G. Neurópteros (Insecta, Planipennia) da Formaçao Santana (Cretáceo Inferior) Bacia do Araripe, Nordeste do Brasil. V—Aspectos filogenéticos, paleoecológicos, paleobiogeogr ficos e descriçao de novos taxa. An. Acad. Bras. Cienc. 1992, 64, 117–148. [Google Scholar]
- Tjeder, B. Neuroptera-Planipennia. The Lace-wings of Southern Africa. 6. Family Nemopteridae. In South African Animal Life; Hanström, B., Brinck, P., Rudebec, G., Eds.; Swedish Natural Science Research Council: Stockholm, Sweden, 1967; Volume 13, pp. 290–501. [Google Scholar]
- Wang, X.L.; Bao, R. A taxonomic study on the genus Balmes Navás from China (Neuroptera, Psychopsidae). Acta Zootaxon. Sin. 2006, 31, 846–850. [Google Scholar]
- Riek, E.F. The Australian moth-lacewings (Neuroptera: Ithonidae). J. Aust. Entomol. Soc. 1974, 13, 37–54. [Google Scholar] [CrossRef]
- Orfila, R.N. Un nuevo Nemopteridae (Neuropt.) americano con una sinopsis de la familia. R. Soc. Entomol. Arg. 1954, 17, 29–32. [Google Scholar]
- Califre Martins, C.; Ardila-Camacho, A.; Aspöck, U. Neotropical osmylids (Neuroptera, Osmylidae): Three new species of Isostenosmylus Krüger, 1913, new distributional records, redescriptions, checklist and key for the Neotropical species. Zootaxa 2016, 4149, 1–66. [Google Scholar] [CrossRef]

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lu, X.; Xu, C.; Liu, X. New Cretaceous Lacewings in a Transitional Lineage of Myrmeleontoidea and Their Phylogenetic Implications. Insects 2022, 13, 429. https://doi.org/10.3390/insects13050429
Lu X, Xu C, Liu X. New Cretaceous Lacewings in a Transitional Lineage of Myrmeleontoidea and Their Phylogenetic Implications. Insects. 2022; 13(5):429. https://doi.org/10.3390/insects13050429
Chicago/Turabian StyleLu, Xiumei, Chunpeng Xu, and Xingyue Liu. 2022. "New Cretaceous Lacewings in a Transitional Lineage of Myrmeleontoidea and Their Phylogenetic Implications" Insects 13, no. 5: 429. https://doi.org/10.3390/insects13050429