
Mechanisms of Trichomes and Terpene Compounds in Indigenous and Commercial Thai Rice Varieties against Brown Planthopper


 ,
, 
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Design
2.2. Materials
2.2.1. Rice Plant
2.2.2. Brown Planthopper Mass Rearing
2.3. Evaluation for Brown Planthopper Resistance
2.4. Morphology Observation Using Scanning Electron Microscopy (SEM)
2.5. VOCs Collection and Identification of Rice Terpenes and Derivatives by the Constitutive Defense and Inducible Defense
2.5.1. VOC Collections for Constitutive and Induced Defense
2.5.2. Extraction and GC-MS Analysis
2.6. Relative Preference of Brown Planthopper between RH and the Other Varieties of Rice Plants
2.7. Statistical Analysis
3. Results
3.1. Screening for Brown Planthopper Resistance
3.2. Scanning Electron Microscopy (SEM) Observation
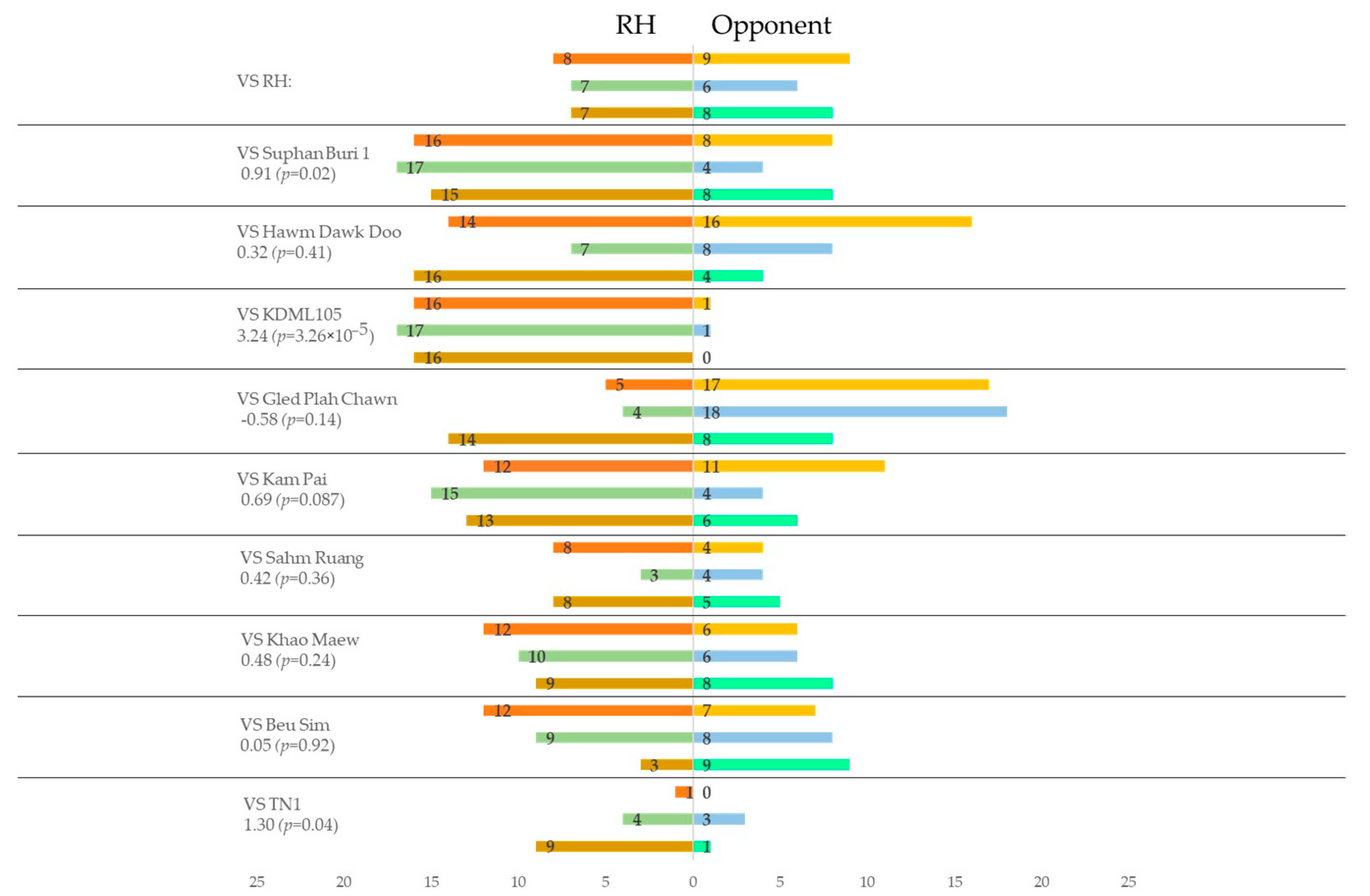
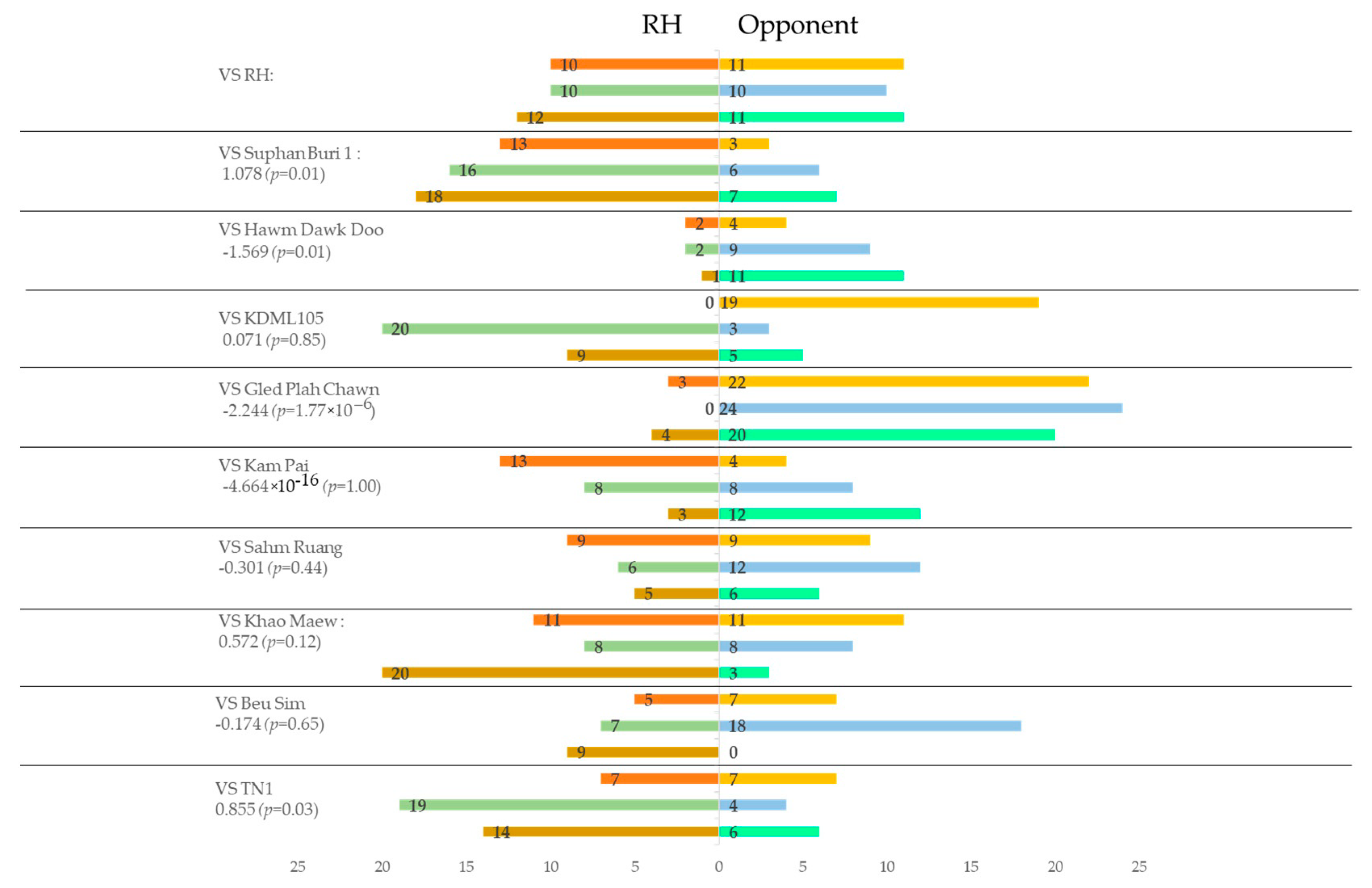
3.3. Preference of Brown Planthopper to VOC Blends from Different Rice Plants
3.4. VOCs 10 Rice Varieties before and during BPH Infestation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jairin, J.; Toojinda, T.; Tragoonrung, S.; Tayapat, S.; Vanavichit, A. Multiple genes determining brown planthopper (Nilaparvata lugens Stal) resistance in backcross introgressed lines of Thai Jasmine rice ‘KDML105’. Sci. Asia 2005, 31, 129–135. [Google Scholar] [CrossRef]
- Rubia-Sanchez, E.; Suzuki, Y.; Miyamoto, K.; Watanabe, T. The potential for compensation of the effects of the brown planthopper Nilaparvata lugens Stal (Homoptera: Delphacidae) feeding on rice. Crop Prot. 1999, 18, 39–45. [Google Scholar] [CrossRef]
- Sōgawa, K. The rice brown planthopper: Feeding physiology and host plant interactions. Annu. Rev. Entomol. 1982, 27, 49–73. [Google Scholar] [CrossRef]
- You, L.-L.; Wu, Y.; Xu, B.; Ding, J.; Ge, L.-Q.; Yang, G.-Q.; Song, Q.-S.; Stanley, D.; Wu, J.-C. Driving pest insect populations: Agricultural chemicals lead to an adaptive syndrome in Nilaparvata lugens Stål (Hemiptera: Delphacidae). Sci. Rep. 2016, 6, 37430. [Google Scholar] [CrossRef] [Green Version]
- Gould, F.; Brown, Z.; Kuzma, J. Wicked evolution: Can we address the sociobiological dilemma of pesticide resistance? Science 2018, 360, 728–732. [Google Scholar] [CrossRef] [Green Version]
- Kole, R.; Roy, K.; Panja, B.; Egambaram, S.; Mondal, T.; Worede, R. Use of pesticides in agriculture and emergence of resistant pests. Indian J. Anim. Health 2020, 58, 53–70. [Google Scholar] [CrossRef]
- Raymond, S.; Nugegoda, D.; Kibria, G. The Effects of Pulse Exposure of Six Agricultural Chemicals (Including four Herbicides) on the Early Life Stages of Selected Native Fish from the Goulburn-Murray River Region; RMIT University: Melbourne, Australia, 2014; 89p. [Google Scholar]
- Ali, S.; Ullah, M.; Sajjad, A.; Shakeel, Q.; Hussain, A. Environmental and health effects of pesticide residues. In Sustainable Agriculture Reviews 48; Inamuddin, I., Ahamed, M.I., Lichtfouse, E., Eds.; Springer: Cham, Switzerland, 2021. [Google Scholar]
- Hanley, M.; Lamont, B.; Fairbanks, M.; Rafferty, C. Plant structural traits and their role in anti-herbivore defence. Plant Ecol. Evol. Syst. 2007, 157–178. [Google Scholar] [CrossRef]
- Joel, F.-H.; Mika, Z.; Søren, B. Plant Defense against Insect Herbivores. Int. J. Mol. Sci. 2013, 14, 10242–10297. [Google Scholar]
- Pegadaraju, V.; Knepper, C.; Reese, J.; Shah, J. Premature leaf senescence modulated by the Arabidopsis PHYTOALEXIN DEFICIENT4 gene is associated with defense against the phloem-feeding green peach aphid. Plant Physiol. 2006, 139, 1927–1934. [Google Scholar] [CrossRef] [Green Version]
- Yan, A.; Pan, J.; An, L.; Gan, Y.; Feng, H. The responses of trichome mutants to enhanced ultraviolet-B radiation in Arabidopsis thaliana. Photochem. Photobiol. 2012, 113, 29–35. [Google Scholar] [CrossRef]
- Bickford, C.P. Ecophysiology of leaf trichomes. Funct. Plant Biol. 2016, 43, 807–814. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hauser, M.T. Molecular basis of natural variation and environmental control of trichome patterning. Front. Plant Sci. 2014, 5, 320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Inomura, K.; Bragg, J.; Follows, M.J. A quantitative analysis of the direct and indirect costs of nitrogen fixation: A model based on Azotobacter vinelandii. ISME J. 2017, 11, 166–175. [Google Scholar] [CrossRef] [PubMed]
- Eisner, T.; Eisner, M.; Hoebeke, E.R. When defense backfires: Detrimental effect of a plant’s protective trichomes on an insect beneficial to the plant. Proc. Natl. Acad. Sci. USA 1998, 95, 4410–4414. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Handley, R.; Ekbom, B.; Ågren, J. Variation in trichome density and resistance against a specialist herbivore in natural populations of Arabidopsis thaliana. Ecol. Entomol. 2005, 30, 284–292. [Google Scholar] [CrossRef]
- Huttunen, P.; Karkkainen, K.; Lee, G.; Rautio, P.; Agren, J. Leaf trichome production and responses to defoliation and drought in Arabidopsis lyrata (Brassicaceae). Ann. Bot. Fenn. 2010, 47, 199–207. [Google Scholar] [CrossRef]
- Levin, D.A. The Role of Trichomes in Plant Defense. Q. Rev. Biol. 1973, 48, 3–15. [Google Scholar] [CrossRef]
- Ascensao, L.; Marques, N.; Pais, M. Peltate Glandular Trichomes of Leonotis leonurus Leaves: Ultrastructure and Histochemical Characterization of Secretions. Int. J. Plant Sci. 1997, 158, 249–258. [Google Scholar] [CrossRef]
- Faculty of Biology. Glandular. Available online: https://mmegias.webs.uvigo.es/02-english/1-vegetal/guiada_v_secretor.php (accessed on 24 February 2022).
- Glas, J.; Schimmel, B.; Alba Cano, J.; Escobar-Bravo, R.; Schuurink, R.; Kant, M. Plant Glandular Trichomes as Targets for Breeding or Engineering of Resistance to Herbivores. Int. J. Mol. Sci. 2012, 13, 17077–17103. [Google Scholar] [CrossRef] [Green Version]
- Schilmiller, A.L.; Last, R.L.; Pichersky, E. Harnessing plant trichome biochemistry for the production of useful compounds. Plant J. 2008, 54, 702–711. [Google Scholar] [CrossRef] [Green Version]
- Huchelmann, A.; Boutry, M.; Hachez, C. Plant glandular trichomes: Natural cell factories of high biotechnological interest. Plant Physiol. 2017, 175, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pitija, K.; Kamolsukyunyong, W.; Vanavichi, A.; Sookwong, P.; Mahatheeranont, S. Monoterpenoid Allelochemicals in Resistance Rice Varieties against Brown Planthoppers, Nilaparvata Lugens (Stål). J. Adv. Agric. Technol. 2014, 1, 82. [Google Scholar] [CrossRef]
- Ågren, J.; Schemske, D. The cost of defense against herbivores: An experimental study of trichome production in Brassica rapa. Am. Nat. 1993, 141, 338–350. [Google Scholar] [CrossRef] [PubMed]
- Lam, W.K.F.; Pedigo, L. Effect of trichome density on soybean pod feeding by adult bean leaf beetles (Coleoptera: Chrysomelidae). J. Econ. Entomol. 2002, 94, 1459–1463. [Google Scholar] [CrossRef]
- Roberts, J.; Foster, J. Effect of Leaf Pubescence in Wheat on the Bird Cherry Oat Aphid (Homoptera: Aphidae). J. Econ. Entomol. 1983, 76, 1320–1322. [Google Scholar] [CrossRef]
- Bleeker, P.M.; Diergaarde, P.J.; Ament, K.; Schütz, S.; Johne, B.; Dijkink, J.; Hiemstra, H.; de Gelder, R.; de Both, M.T.J.; Sabelis, M.W.; et al. Tomato-produced 7-epizingiberene and R-curcumene act as repellents to whiteflies. Phytochemistry 2011, 72, 68–73. [Google Scholar] [CrossRef]
- Wang, Y.; Li, H.; Si, Y.; Zhang, H.; Guo, H.; Miao, X. Microarray analysis of broad-spectrum resistance derived from an indica cultivars Rathu heenati. Planta 2012, 235, 829–840. [Google Scholar] [CrossRef]
- Li, Y.; Huang, Y.-F.; Huang, S.-H.; Kuang, Y.-H.; Tung, C.-W.; Liao, C.-T.; Chuang, W.-P. Genomic and phenotypic evaluation of rice susceptible check TN1 collected in Taiwan. Bot. Stud. 2019, 60, 19. [Google Scholar] [CrossRef] [Green Version]
- Poonsin, R.; Parinthawong, N. Investigation of rice blast resistant genes in Thai elite rice varieties (Oryza sativa L.) for improvement of broad-spectrum blast disease resistance variety. Int. J. Agric. Technol. 2020, 16, 109–118. [Google Scholar]
- Rice Knowledge Bank. Standard Evaluation System for Rice (SES). 2002, p. 20. Available online: http://www.knowledgebank.irri.org/images/docs/rice-standard-evaluation-system.pdf (accessed on 25 February 2022).
- IRRI. Standard Evaluation System for Rice; International Rice Research Institute: Manila, Philippines, 1980. [Google Scholar]
- Litsinger, J.; Litsinger, J.A. Crop loss assessment in rice. In Rice Insects: Management Strategies; Heinrichs, E.A., Miller, T.A., Eds.; Springer: New York, NY, USA, 1991; pp. 1–65. [Google Scholar]
- Kamolsukyunyong, W.; Ruengphayak, S.; Chumwong, P.; Kusumawati, L.; Chaichoompu, E.; Jamboonsri, W.; Saensuk, C.; Phoonsiri, K.; Toojinda, T.; Vanavichit, A. Identification of spontaneous mutation for broad-spectrum brown planthopper resistance in a large, long-term fast neutron mutagenized rice population. Rice 2019, 12, 16. [Google Scholar] [CrossRef] [PubMed]
- Song, H.; Wang, X.; Zhou, X.; Zhang, K.; Chen, X.; Liu, J.; Han, J.; Wang, A. The roles of different types of trichomes in tomato resistance to cold, drought, whiteflies, and botrytis. Agronomy 2020, 10, 411. [Google Scholar]
- Zorrilla, C.; Schabow, J.; Chernov, V.; Palta, J. CAX1 vacuolar antiporter overexpression in potato results in calcium deficiency in leaves and tubers by sequestering calcium as calcium oxalate. Crop Sci. 2018, 59, 176–189. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; Qiang, Z.; Xia, Q.; Zhang, W.; Zhang, G.; Gu, D. Effects of herbivore-induced rice volatiles on the host selection behavior of brown planthopper, Nilaparvata lugens. Chin. Sci. Bull. 2002, 47, 1355–1360. [Google Scholar] [CrossRef]
- Giunti, G.; Palmeri, V.; Algeri, G.; Campolo, O. VOC emissions influence intra- and interspecific interactions among stored-product Coleoptera in paddy rice. Sci. Rep. 2018, 8, 2052. [Google Scholar] [CrossRef] [Green Version]
- Vijayaraghavendra, R.; Lakshmi, K.; Shanker, C.; Seetalam, M.; Jagadeeshwar, R.; Chennamadhavuni, D. Olfactory response of insect predators of rice brown plant hopper (Nilaparvata lugens (Stal.)) to flower volatiles. J. Entomol. Zool. Stud. 2019, 7, 1095–1099. [Google Scholar]
- R Core Team. R: A language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria; Available online: https://www.R-project.org/ (accessed on 5 April 2022).
- Ogle, D.H.; Doll, J.C.; Wheeler, P.; Dinno, A. FSA: Fisheries Stock Analysis. R Package Version 0.9.3. Available online: https://github.com/fishR-Core-Team/FSA (accessed on 5 April 2022).
- Felipe, d.M.; Muhammad, Y. agricolae: Statistical Procedures for Agricultural Research. R Package Version 1.4. Available online: https://myaseen208.github.io/agricolae/https://cran.r-project.org/package=agricolae (accessed on 5 April 2022).
- Jairin, J.; Teangdeerith, S.; Leelagud, P.; Phengrat, K.; Vanavichit, A.; Toojinda, T. Physical mapping of Bph3, a brown planthopper resistance locus in rice. Maejo Int. J. Sci. Technol. 2007, 1, 166–177. [Google Scholar]
- Velusamy, R.; Kumar, M.; Edward, Y. Mechanisms of resistance to the brown planthopper Nilaparvata lugens in wild rice (Oryza spp.) cultivars. Entomol. Exp. Appl. 2011, 74, 245–251. [Google Scholar] [CrossRef]
- Lakshminarayana, A.; Khush, G. New genes for resistance to brown planthopper in rice. Crop Sci. 1976, 17, 96–100. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Su, C.; Wang, C.; Zhai, H.; Wan, J. Mapping of a major resistance gene to the brown planthopper in the rice cultivar Rathu Heenati. Breed. Sci. 2005, 55, 391–396. [Google Scholar] [CrossRef] [Green Version]
- Jairin, J.; Phengrat, K.; Teangdeerith, S.; Vanavichit, A.; Toojinda, T. Mapping of a broad-spectrum brown planthopper resistance gene, Bph3, on rice chromosome 6. Mol. Breed. 2007, 19, 35–44. [Google Scholar] [CrossRef]
- Kamolsukyunyong, W.; Sukhaket, W.; Ruanjaichon, V.; Toojinda, T.; Vanavichit, A. Single-feature polymorphism mapping of isogenic rice lines identifies the influence of terpene synthase on brown planthopper feeding preferences. Rice 2013, 6, 18. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deen, R.; Kandu, R.; Padmavathi, G.; Viraktamath, B.; Ram, T. Mapping of brown planthopper [Nilaparvata lugens (Stål)] resistance gene (bph5) in rice (Oryza sativa L.). Euphytica 2017, 213, 35. [Google Scholar] [CrossRef]
- Vanavichit, A.; Kamolsukyeunyong, W.; Siangliw, M.; Siangliw, J.L.; Traprab, S.; Ruengphayak, S.; Chaichoompu, E.; Saensuk, C.; Phuvanartnarubal, E.; Toojinda, T.; et al. Thai Hom Mali Rice: Origin and Breeding for Subsistence Rainfed Lowland Rice System. Rice 2018, 11, 20. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xing, Z.; Liu, Y.; Cai, W.; Huang, X.; Wu, S.; Lei, Z. Efficiency of Trichome-Based Plant Defense in Phaseolus vulgaris Depends on Insect Behavior, Plant Ontogeny, and Structure. Front. Plant Sci. 2017, 8, 2006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, G.; Klinkhamer, P.G.L.; Escobar-Bravo, R.; Leiss, K.A. Type VI glandular trichome density and their derived volatiles are differently induced by jasmonic acid in developing and fully developed tomato leaves: Implications for thrips resistance. Plant Sci. 2018, 276, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Andama, J.; Mujiono, K.; Hojo, Y.; Shinya, T.; Galis, I. Non-glandular silicified trichomes are essential for rice defense against chewing herbivores. Plant Cell Environ. 2020, 43, 22019–22032. [Google Scholar] [CrossRef]
- Sandhu, R.; Sarao, P. Evaluation of antixenosis resistance in wild rice accessions against brown planthopper, Nilaparvata lugens (Stål). Int. J. Trop. Insect Sci. 2021, 41, 65–73. [Google Scholar] [CrossRef]
- Maluf, W.; Inoue, I.; Ferreira, R.; Gomes, L.; Castro, E.; Cardoso, M. Higher glandular trichome density in tomato leaflets and repellence to spider mites. Pesquisa Agropecuaria Brasileira 2007, 42, 1227–1235. [Google Scholar] [CrossRef] [Green Version]
- Schuurink, R.; Tissier, A. Glandular trichomes: Micro-organs with model status? New Phytol. 2019, 225, 2251–2266. [Google Scholar] [CrossRef] [Green Version]
- Youn, Y. Electroantennogram Responses of Nilaparvata lugens (Homoptera: Delphacidae) to Plant Volatile Compounds. J. Econ. Entomol. 2002, 95, 269–277. [Google Scholar] [CrossRef] [PubMed]
- Xiao, Y.; Wang, Q.; Erb, M.; Turlings, T.; Ge, L.; Hu, L.; Li, J.; Han, X.; Zhang, T.; Lu, J.; et al. Specific herbivore-induced volatiles defend plants and determine insect community composition in the field. Ecol. lett. 2012, 15, 1130–1139. [Google Scholar] [CrossRef] [PubMed]
- Kamolsukyunyong, W.; Sukhaket, W.; Pitija, K.; Thorngkham, P.; Mahatheeranont, S.; Toojinda, T.; Vanavichit, A.; Borrelli, M.; Marone, D. Rice Sesquiterpene Plays Important Roles in Antixenosis against Brown Planthopper in Rice. Plants 2021, 10, 1049. [Google Scholar] [CrossRef] [PubMed]




{kind=link}
{kind=link}
{kind=link}
| Variety | Varieties Description | Originated | Resistance to BPH | Reference |
|---|---|---|---|---|
| Beu Sim | Indigenous rice variety | Thailand | Data not found | - |
| Gled Plah Chawn | Indigenous rice variety | Thailand | Data not found | - |
| Hawm Dawk Doo | Indigenous rice variety | Thailand | Data not found | - |
| Kam Pai | Indigenous rice variety | Thailand | Data not found | - |
| Khao Maew | Indigenous rice variety | Thailand | Data not found | - |
| Sahm Ruang | Indigenous rice variety | Thailand | Data not found | - |
| Rathu Heenati (RH) | Resistant donor variety in the conventional breeding program | Sri Lanka | Resistant | [30] |
| Taichung Native 1 (TN1) | Standard susceptible variety | Taiwan | Susceptible | [31] |
| Khao Dawk Mali 105 (KDML105) | Commercial variety | Thailand | Susceptible | [32] |
| Suphan Buri 1 | Commercial variety | Thailand | Resistant | [32] |
| Varieties | Injury Score by SES (0–9) | BPH Damage AUC | Evaluated BPH Resistance | ||
|---|---|---|---|---|---|
| 7 DAI | 9 DAI | 11 DAI | |||
| RH | 0.0 (0–0) a | 0.2 (0–1) a | 6.0 (3–9) ab | 3.2 (1.5–4.5) a | HR |
| Suphan Buri 1 | 0.0 (0–0) ab | 2.0 (1–3) ac | 6.7 (5–9) a | 5.3 (3.5–7.5) ac | R |
| Hawm Dawk Doo | 0.3 (0–1) ab | 4.2 (0–7) abc | 9.0 (9–9) c | 8.8 (4.5–11.5) abc | I |
| KDML105 | 0.7 (0–3) ab | 5.3 (5–7) bc | 8.7 (7–9) c | 10.0 (8.5–12.0) bc | I |
| Gled Plah Chawn | 1.3 (0–5) ab | 5.0 (3–7) bc | 9.0 (9–9) c | 10.1 (7.5–14.0) abc | I |
| Kam Pai | 2.3 (0–5) ab | 5.0 (1–7) bc | 9.0 (9–9) c | 10.6 (5.5–14.0) b | S |
| Sahm Ruang | 2.2 (0–5) ab | 5.3 (5–7) bc | 8.3 (5–9) bc | 10.6 (8.0–14.0) b | S |
| Khao Maew | 3.2 (0–7) ab | 5.3 (3–7) bc | 9.0 (9–9) c | 11.4 (7.5–15.0) b | S |
| Beu Sim | 1.8 (0–3) ab | 6.3 (5–7) b | 9.0 (9–9) c | 11.8 (9.5–13.0) b | S |
| TN1 | 3.8 (0–7) b | 5.7 (1–9) b | 9.0 (9–9) c | 12.1 (5.5–17.0) b | S |
| Mean | 1.0 | 4.6 | 8.5 | 9.4 | |
| p-value | 0.006 | 0.0007 | 0.0002 | 0.0004 | |
| Rice Varieties | Glandular Trichome | Non-Glandular Trichome | |||
|---|---|---|---|---|---|
| Prickle | Micro | Macro | Papillae | ||
| RH | 35.10 ± 0.49 b | 36.00 ± 1.80 d | 16.80 ± 0.75 c | 247.67 ± 12.44 b | 15.80 ± 0.57 bc |
| Suphan Buri 1 | 37.93 ± 3.66 ab | 42.77 ± 3.84 cd | 16.67 ± 2.50 c | 170.33 ± 17.46 c | 15.60 ± 0.92 bc |
| Hom Dawk Doo | 44.83 ± 2.23 a | 74.47 ± 4.37 a | 0.00 ± 0.00 d | 249.33 ± 13.78 b | 19.00 ± 0.25 a |
| KDML105 | 38.03 ± 1.42 ab | 0.00 ± 0.00 e | 0.00 ± 0.00 d | 157.33 ± 20.85 cd | 18.07 ± 1.01 ab |
| Gled Plah Chawn | 40.23 ± 4.36 ab | 55.10 ± 7.95 b | 25.53 ± 1.58 a | 125.33 ± 0.88 cd | 14.50 ± 0.75 c |
| Kam Pai | 42.37 ± 0.27 ab | 45.83 ± 1.67 bcd | 21.40 ± 2.63 b | 110.80 ± 5.88 d | 14.83 ± 2.09 c |
| Sahm Ruang | 40.23 ± 2.14 ab | 0.00 ± 0.00 e | 23.03 ± 0.22 ab | 229.33 ± 7.31 b | 15.33 ± 0.60 bc |
| Khao Maew | 40.20 ± 0.87 ab | 0.00 ± 0.00 e | 0.00 ± 0.00 d | 342.67 ± 34.23 a | 15.00 ± 0.31 c |
| Beu Sim | 40.87 ± 1.91 ab | 48.47 ± 4.60 bc | 22.73 ± 0.22 ab | 162.33 ± 9.96 cd | 16.00 ± 0.26 bc |
| TN1 | 37.90 ± 1.65 ab | 43.23 ± 4.43 cd | 0.00 ± 0.00 d | 108.13 ± 19.45 d | 14.53 ± 0.52 c |
| Rice Varieties | All Trichomes | Glandular Trichome | Non-glandular Trichome | |||
|---|---|---|---|---|---|---|
| Prickle | Micro | Macro | Papillae | |||
| RH | 11.42 ± 0.58 c | 0.33 ± 0.38 e | 0.58 ± 0.14 b | 0.42 ± 0.14 bc | 0.17 ± 0.14 e | 9.92 ± 0.52 c |
| Suphan Buri 1 | 10.92 ± 1.04 cd | 1.75 ± 0.43 c | 1.58 ± 1.26 a | 0.83 ± 0.29 a | 0.42 ± 0.14 de | 6.33 ± 0.88 cde |
| Hom Dawk Doo | 15.67 ± 3.25 b | 2.50 ± 0.50 b | 0.17 ± 0.14 b | 0.00 ± 0.00 d | 2.58 ± 0.95 a | 10.42 ± 3.75 b |
| KDML105 | 8.42 ± 0.58 cde | 0.67 ± 0.14 de | 0.00 ± 0.00 b | 0.00 ± 0.00 d | 1.75 ± 0.25 b | 6.00 ± 0.50 de |
| Gled Plah Chawn | 5.42 ± 1.28 e | 0.83 ± 0.14 de | 0.17 ± 0.14 b | 0.67 ± 0.29 ab | 0.17 ± 0.29 e | 3.58 ± 0.95 e |
| Kam Pai | 7.75 ± 1.98 de | 0.58 ± 0.14 de | 0.17 ± 0.29 b | 0.92 ± 0.38 a | 0.92 ± 0.29 cde | 5.17 ± 2.57 de |
| Sahm Ruang | 11.50 ± 1.64 c | 2.58 ± 0.63 b | 0.00 ± 0.00 b | 0.17 ± 0.14 cd | 0.67 ± 0.29 cde | 8.08 ± 1.04 bcd |
| Khao Maew | 21.33 ± 2.01 a | 3.67 ± 0.38 a | 0.00 ± 0.00 b | 0.00 ± 0.00 d | 2.50 ± 0.25 a | 15.17 ± 2.27 a |
| Beu Sim | 8.25 ± 1.15 cde | 1.25 ± 0.43 cd | 0.25 ± 0.00 b | 0.25 ± 0.25 cd | 1.08 ± 0.52 bcd | 5.42 ± 1.59 de |
| TN1 | 9.33 ± 3.26 cd | 1.75 ± 0.66 c | 0.08 ± 0.14 b | 0.00 ± 0.00 d | 1.33 ± 0.29 bc | 6.17 ± 3.06 de |
| Length | Density | |
|---|---|---|
| All trichomes | NA | −0.15 (p = 0.42) |
| Grandular trichome | 0.05 (p = 0.80) | 0.27 (p = 0.15) |
| Prickle trichome | 0.05 (p = 0.79) | −0.43 (p = 0.016) |
| Micro trichome | −0.10 (p = 0.60) | −0.33 (p = 0.08) |
| Macro trichome | −0.32 (p = 0.08) | 0.44 (p = 0.016) |
| Papillae | −0.10 (p = 0.60) | −0.20 (p = 0.27) |
| Compounds | RH | Suphan Buri 1 | Hawm Dawk Doo | KDML105 | Gled Plah Chawn | Kam Pai | Sahm Ruang | Khao Maew | Beu Sim | TN1 | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| w/o | w | w/o | w | w/o | w | w/o | w | w/o | w | w/o | w | w/o | w | w/o | w | w/o | w | w/o | w | ||
| Monoterpenoids | Total | 0 | 0 | 2 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 2 | 0 |
| Bicyclo[3.1.0]hex-2-ene,4-methyl-1-(1-methylethyl)- | √ | ||||||||||||||||||||
| D-Limonene | √ | ||||||||||||||||||||
| α-Phellandrene | √ | √ | |||||||||||||||||||
| β-Myrcene | √ | √ | |||||||||||||||||||
| Sesquiterpenoids | Total | 2 | 1 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 0 | 0 | 1 | 1 | 1 | 0 | 0 | 1 | 0 |
| (S,1Z,6Z)-8-Isopropyl-1-methyl-5-methylenecyclodeca-1,6-diene | √ | ||||||||||||||||||||
| Aromandendrene | √ | ||||||||||||||||||||
| Caryophyllene | √ | √ | |||||||||||||||||||
| α-Cubebene | √ | √ | √ | ||||||||||||||||||
| β-Caryophyllen | √ | ||||||||||||||||||||
| β-Curcumene | √ | ||||||||||||||||||||
| β-Sesquiphellandrene | √ | ||||||||||||||||||||
| Norsesquiterpenes | Total | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 |
| Geijerene | √ | ||||||||||||||||||||
| Carbonyl compounds | Total | 3 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 0 | 0 | 0 | 2 |
| 1-Hepten-3-one | √ | ||||||||||||||||||||
| 1-Penten-3-one | √ | ||||||||||||||||||||
| 4′-Ethylpropiophenone | √ | √ | |||||||||||||||||||
| Ethanone,1-(4-ethylphenyl)- | √ | √ | |||||||||||||||||||
| Terpene | Total | 12 | 2 | 3 | 1 | 2 | 0 | 3 | 1 | 11 | 2 | 5 | 0 | 3 | 2 | 3 | 3 | 2 | 3 | 5 | 2 |
| 1-Methyl-3-(1′-methylcyclopropyl)cyclopentene | √ | ||||||||||||||||||||
| 1-Octene,2-methyl- | √ | ||||||||||||||||||||
| 1,3-Bis(cyclopentyl)-1-cyclopentanone | √ | ||||||||||||||||||||
| 1,3-Cyclohexadiene,5,6-dimethyl- | √ | ||||||||||||||||||||
| 1,3,4,6-Hexanetetrone,1-(4-methylphenyl)-6-phenyl- | √ | ||||||||||||||||||||
| 1,5-Heptadiene,(E)- | √ | ||||||||||||||||||||
| 1,5-Heptadiene,(Z)- | √ | ||||||||||||||||||||
| 1,5-Heptadiene,3-methyl-,(E)- | √ | ||||||||||||||||||||
| 1H-Indene,1-hexadecyl-2,3-dihydro- | √ | ||||||||||||||||||||
| 2,5-Cyclohexadien-1-one,4,4′-(1,2-ethanediylidene)bis[2,6-bis(1,1-dimethylethyl)- | √ | ||||||||||||||||||||
| 3-Ethyl-3-hexene | √ | ||||||||||||||||||||
| 3-Pentanone,2,2,4,4-tetramethyl- | √ | ||||||||||||||||||||
| 3-Undecene,5-methyl- | √ | ||||||||||||||||||||
| 4-(2′,4′,4′-trimethyl-yciclo[4.1.0]hept-2′-en-3′-yl)-3-buten-2-one | √ | ||||||||||||||||||||
| 4-Undecene,6-methyl- | √ | ||||||||||||||||||||
| 5-Hepten-3-one,5-ethyl-4-methyl- | √ | √ | |||||||||||||||||||
| Benzene,(2,2-dimethylpropyl)- | √ | ||||||||||||||||||||
| Benzene,[(cyclohex-1-en-1-yl)methyl]- | √ | ||||||||||||||||||||
| Benzene,1,2-diethyl- | √ | ||||||||||||||||||||
| Benzene,1,3-diethyl | √ | ||||||||||||||||||||
| Benzene,1,4-diethyl- | √ | √ | √ | √ | |||||||||||||||||
| Bicyclo[5.2.0]nonane,2-methylene-4,8,8-trimethyl-4-vinyl- | √ | ||||||||||||||||||||
| cis-Muurola-4(15),5-diene | √ | √ | |||||||||||||||||||
| Cyclobutane,1,2-bis(1-methylethenyl)-,trans- | √ | ||||||||||||||||||||
| Cyclobutane,1,2-dipropenyl- | √ | ||||||||||||||||||||
| Cyclobutane,1,3-diisopropenyl-,trans | √ | √ | √ | √ | √ | √ | √ | √ | √ | ||||||||||||
| Cyclobutanone,2,3,3,4-tetramethyl- | √ | ||||||||||||||||||||
| Cyclohexene,1-(1-propynyl)- | √ | ||||||||||||||||||||
| Cyclopentanone,2,2,5-trimethyl- | √ | ||||||||||||||||||||
| Cyclopropane,1-(2-methylbutyl)-1-(1-methylpropyl)- | √ | ||||||||||||||||||||
| Decane,3,8-dimethyl- | √ | ||||||||||||||||||||
| Hexane,3-methyl-4-methylene- | √ | ||||||||||||||||||||
| Hexane,3,4-bis(1,1-dimethylethyl)-2,2,5,5-tetramethyl- | √ | √ | √ | √ | |||||||||||||||||
| Naphthalene | √ | √ | √ | √ | √ | √ | √ | √ | √ | √ | |||||||||||
| Naphthalene,1,2,3,4,4a,7-hexahydro-1,6-dimethyl-4-(1-methylethyl)- | √ | √ | |||||||||||||||||||
| Octadecane,2,2,4,15,17,17-hexamethyl-7,12-bis(3,5,5-trimethylhexyl)- | √ | ||||||||||||||||||||
| Santolinatriene | √ | ||||||||||||||||||||
| Tridecane,2,2,4,10,12,12-hexamethyl-7-(3,5,5-trimethylhexyl)- | √ | √ | |||||||||||||||||||
| Sum total | 17 | 3 | 6 | 1 | 2 | 0 | 3 | 1 | 12 | 5 | 6 | 0 | 3 | 3 | 4 | 5 | 2 | 3 | 8 | 4 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Khetnon, P.; Busarakam, K.; Sukhaket, W.; Niwaspragrit, C.; Kamolsukyeunyong, W.; Kamata, N.; Sanguansub, S. Mechanisms of Trichomes and Terpene Compounds in Indigenous and Commercial Thai Rice Varieties against Brown Planthopper. Insects 2022, 13, 427. https://doi.org/10.3390/insects13050427
Khetnon P, Busarakam K, Sukhaket W, Niwaspragrit C, Kamolsukyeunyong W, Kamata N, Sanguansub S. Mechanisms of Trichomes and Terpene Compounds in Indigenous and Commercial Thai Rice Varieties against Brown Planthopper. Insects. 2022; 13(5):427. https://doi.org/10.3390/insects13050427
Chicago/Turabian StyleKhetnon, Phawini, Kanungnid Busarakam, Wissarut Sukhaket, Cholticha Niwaspragrit, Wintai Kamolsukyeunyong, Naoto Kamata, and Sunisa Sanguansub. 2022. "Mechanisms of Trichomes and Terpene Compounds in Indigenous and Commercial Thai Rice Varieties against Brown Planthopper" Insects 13, no. 5: 427. https://doi.org/10.3390/insects13050427