
Characterization of Cold and Heat Tolerance of Bactrocera tau (Walker)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Measuring the SCPs of B. tau at Different Life Stages
2.3. Determining the Tolerance of B. tau to Extreme High and Low Temperatures
2.4. Evaluating the Relationship of Temperature–Time–Mortality in B. tau
2.5. Data Analysis
3. Results
3.1. The SCPs of B. tau at Different Life Stages
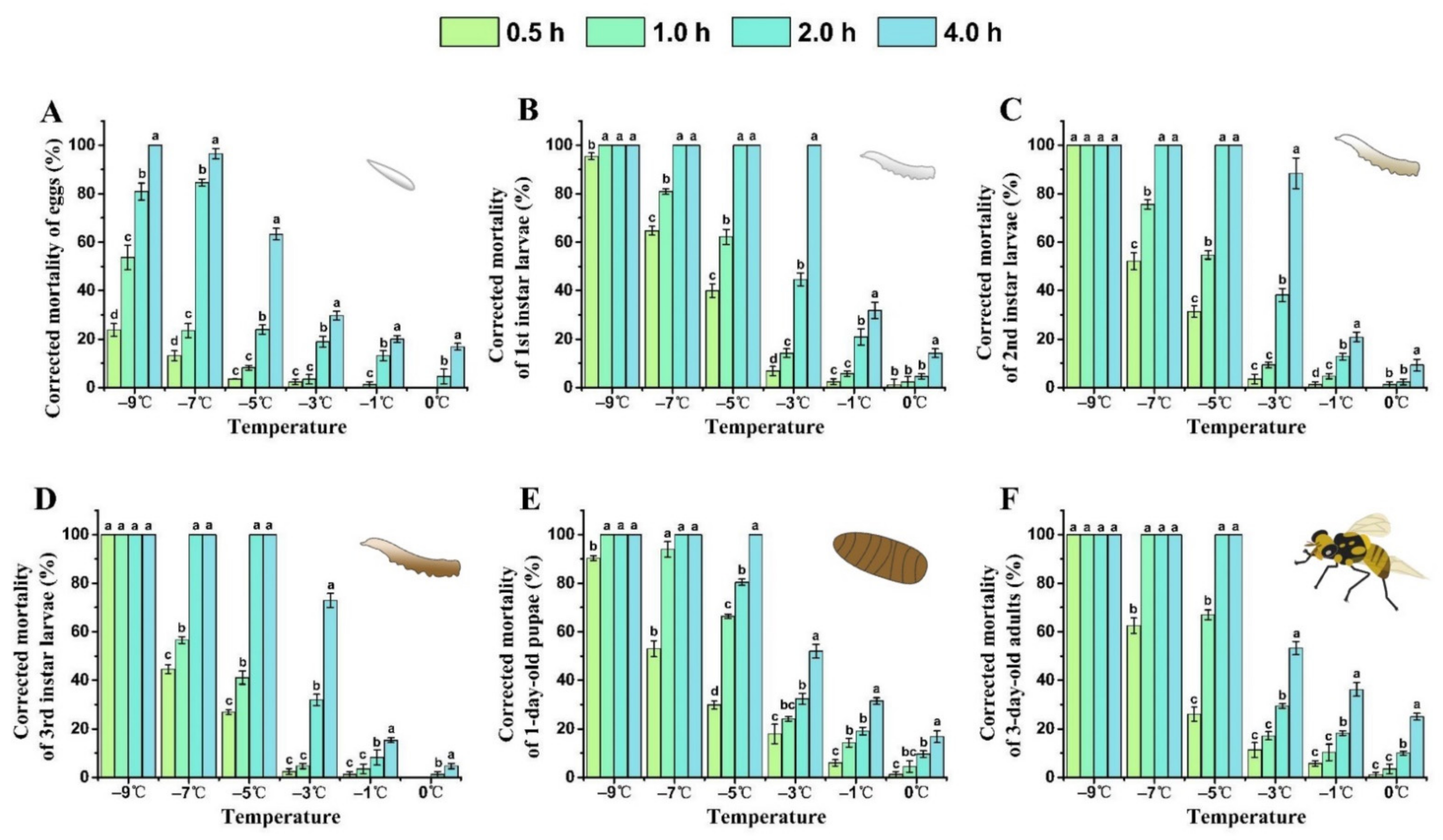
3.2. Cold Tolerance of B. tau at Different Developmental Stages
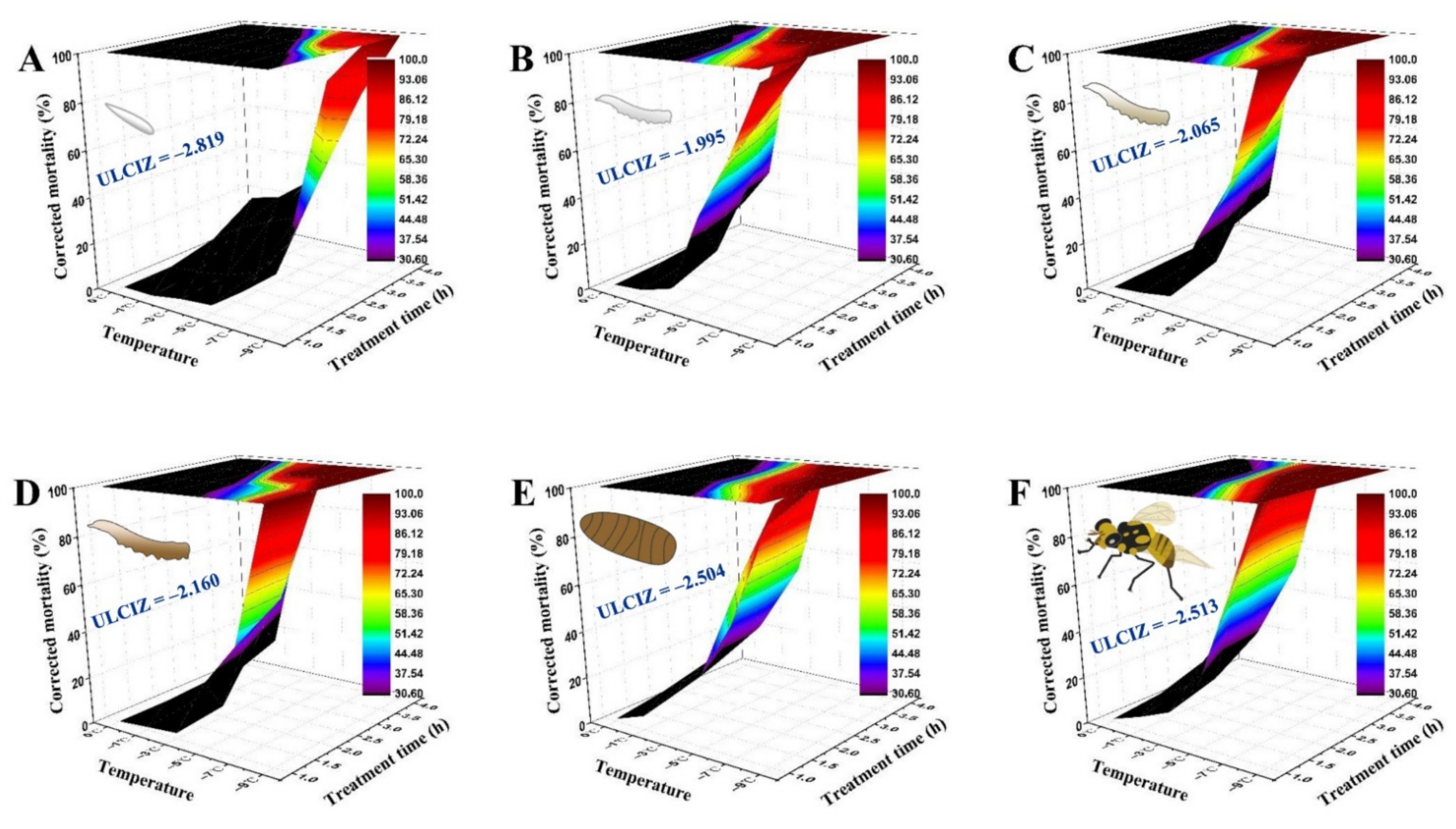
3.3. Interaction between Temperature Exposure Duration and Low Temperature for Mortality of B. tau at Different Developmental Stages
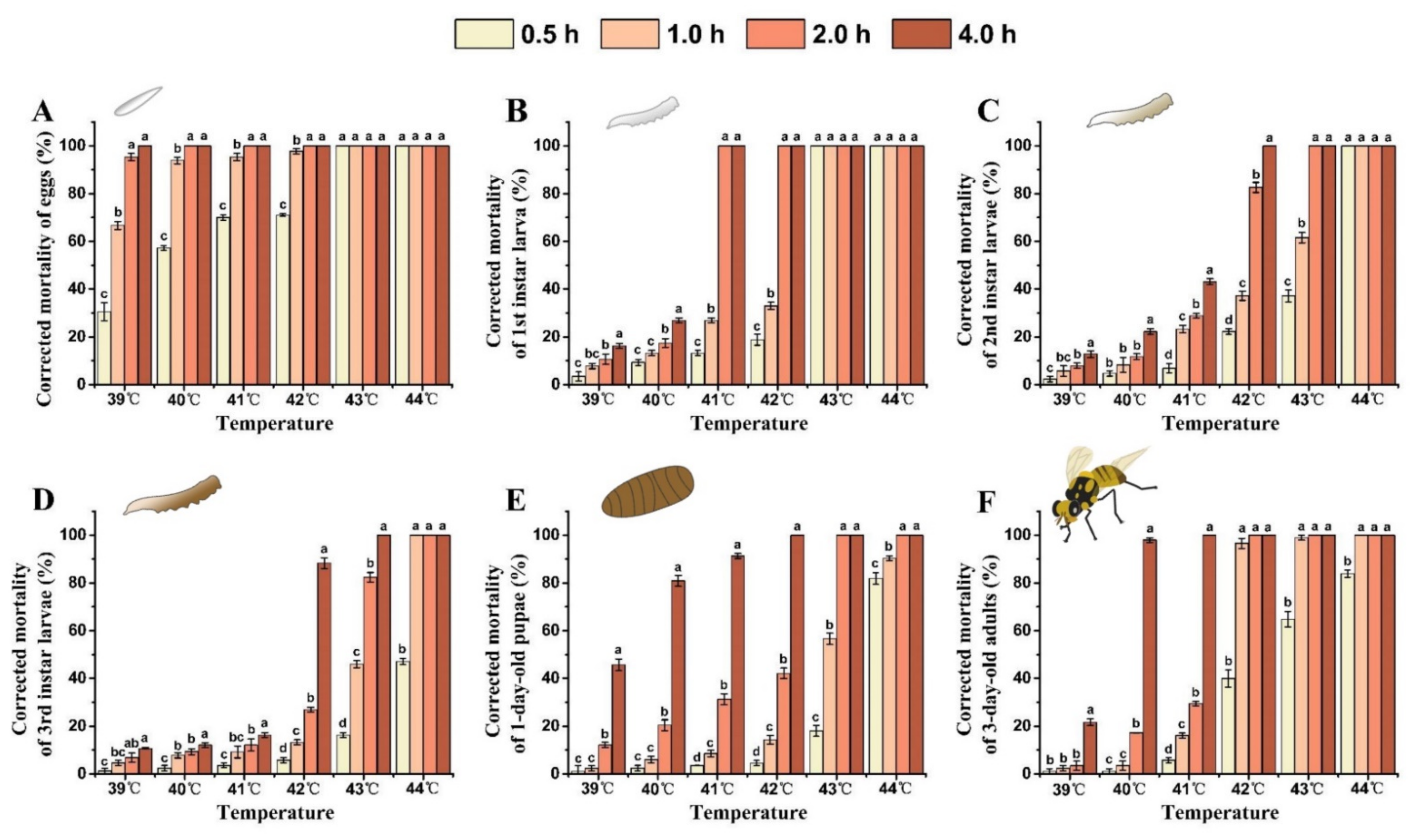
3.4. Heat Tolerance of Different Developmental Stages of B. tau
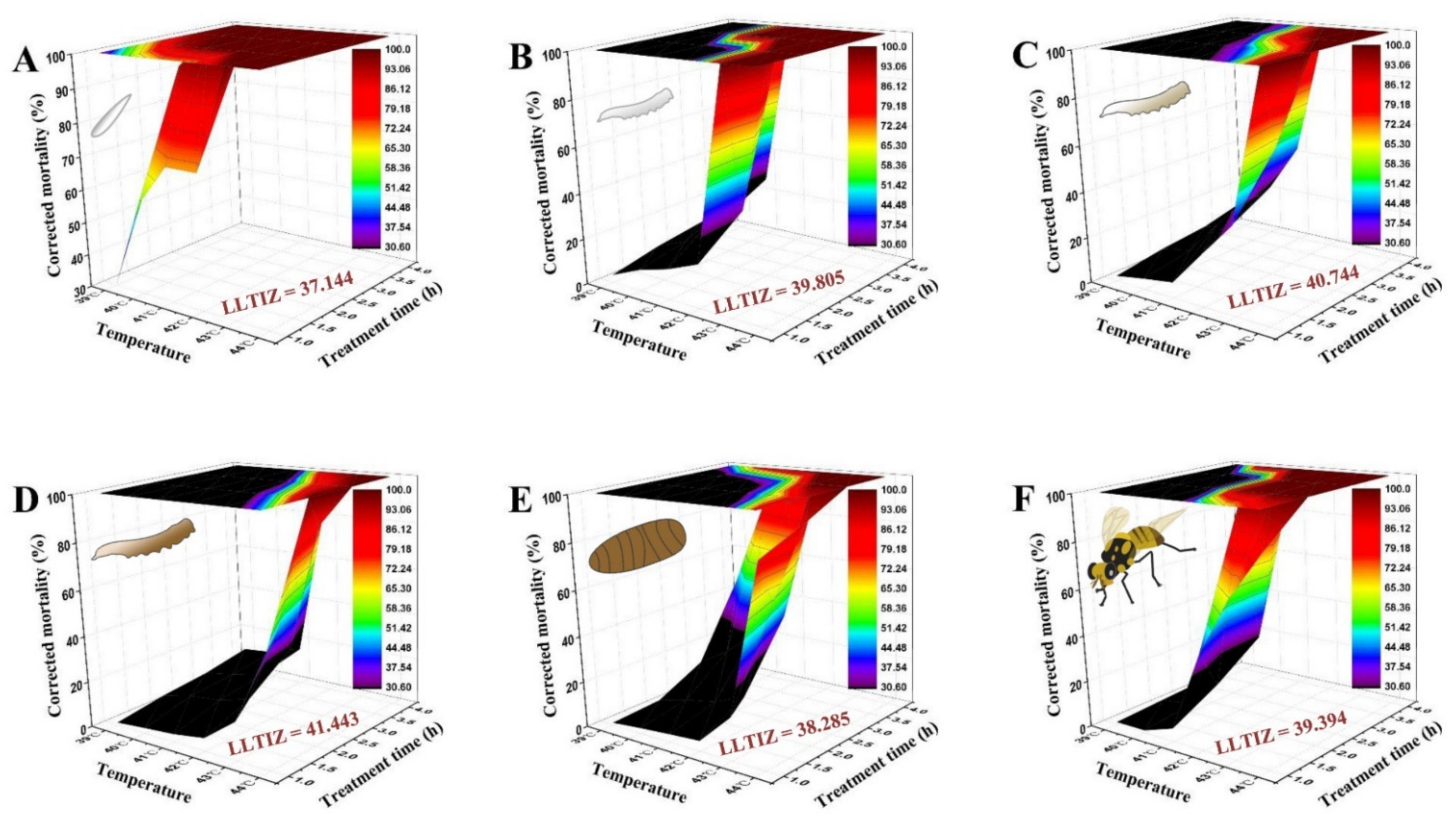
3.5. Interaction between Temperature Exposure Duration and High Temperature for Mortality of B. tau at Different Developmental Stages
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jaleel, W.; Lu, L.; He, Y. Biology, taxonomy, and IPM strategies of Bactrocera tau Walker and complex species (Diptera; Tephritidae) in Asia: A comprehensive review. Environ. Sci. Pollut. R. 2018, 25, 19346–19361. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Gu, X.; Peng, X.; Tao, M.; Peng, L.; Chen, G.; Zhang, X. Effect of short-term low temperature on the growth, development, and reproduction of Bactrocera tau (Diptera: Tephritidae) and Bactrocera cucurbitae. J. Econ. Entomol. 2020, 113, 2141–2149. [Google Scholar] [CrossRef] [PubMed]
- Huang, Y.; Gu, X.; Peng, X.; Tao, M.; Chen, G.; Zhang, X. Effect of short-term high-temperatures on the growth, development and reproduction in the fruit fly, Bactrocera tau (Diptera: Tephritidae). Sci. Rep. 2020, 10, 6418. [Google Scholar] [CrossRef] [PubMed]
- Vasudha, A.; Ahmad, M.A.; Agarwal, M.L. Life history traits and immature stages of Zeugodacus (Zeugodacus) tau (Walker) (Diptera: Tephritidae). J. Entomol. Zool. Stud. 2019, 7, 1149–1153. [Google Scholar]
- An, K.P.; Wu, B.F.; Shen, K.; Zhang, R.J. Charateristic and research progress on control technology of pumpkin fruit fly Bactrocera tau (Walker) (Diptera: Tephritidae). J. Chang. Veg. 2011, 20, 7–13. [Google Scholar]
- Li, X.; Yang, H.; Hu, K.; Wang, J. Temporal dynamics of Bactrocera (Zeugodacus) tau (Diptera: Tephritidae) adults in north Jiangxi, a subtropical area of China revealed by eight years of trapping with cuelure. J. Asia-Pac. Entomol. 2020, 23, 1–6. [Google Scholar] [CrossRef]
- Shi, W.; Kerdelhué, C.; Ye, H. Genetic structure and colonization history of the fruit fly Bactrocera tau (Diptera: Tephritidae) in China and southeast Asia. J. Econ. Entomol. 2014, 107, 1257–1265. [Google Scholar] [CrossRef]
- Khalid, M. Taxonomy of the Bactrocera (Zeugodacus) tau (Tephritidae: Diptera) complex in Asia. Pak. J. Zool. 1999, 31, 219–235. [Google Scholar]
- Hasyim, A.; Muryati, M.; Kogel, D. Population fluctuation of adult males of the fruit fly, Bactrocera tau Walker (Diptera: Tephritidae) in passion fruit orchards in relation to abiotic factors and sanitation. Indones. J. Agr. Sci. 2008, 9, 29–33. [Google Scholar] [CrossRef]
- Ma, C.S.; Ma, G.; Zhao, F. Impact of global warming on cereal aphids. Chin. J. Appl. Entomol. 2014, 51, 1435–1443. [Google Scholar]
- Izadi, H.; Mohammadzadeh, M.; Mehrabian, M. Cold tolerance of the Tribolium castaneum (Coleoptera: Tenebrionidae), under different thermal regimes: Impact of cold acclimation. J. Econ. Entomol. 2019, 112, 1983–1988. [Google Scholar] [CrossRef] [PubMed]
- Enriquez, T.; Colinet, H. Basal tolerance to heat and cold exposure of the spotted wing drosophila, Drosophila suzukii. PeerJ 2017, 5, e3112. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pei, J.; Li, C.; Ren, L.; Zong, S. Factors influencing cold hardiness during overwintering of Streltzoviella insularis (Lepidoptera: Cossidae). J. Econ. Entomol. 2020, 113, 1254–1261. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, J.; Zhang, X.X.; Chang, Y.W.; Du, Y.Z. Differential response of leafminer flies Liriomyza trifolii (Burgess) and Liriomyza sativae (Blanchard) to rapid cold hardening. Insects 2021, 12, 1041. [Google Scholar] [CrossRef]
- Bawa, S.A.; Gregg, P.C.; Del Soccoro, A.P.; Miller, C.; Andrew, N.R. Estimating the differences in critical thermal maximum and metabolic rate of Helicoverpa punctigera (Wallengren) (Lepidoptera: Noctuidae) across life stages. PeerJ 2021, 9, e12479. [Google Scholar] [CrossRef]
- Colinet, H.; Sinclair, B.J.; Vernon, P.; Renault, D. Insects in fluctuating thermal environments. Annu. Rev. Entomol. 2015, 60, 123–140. [Google Scholar] [CrossRef] [Green Version]
- Liu, Z.; Gong, P.; Wu, K.; Wei, W.; Sun, J.; Li, D. Effects of larval host plants on over-wintering preparedness and survival of the cotton bollworm, Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). J. Insect Physiol. 2007, 53, 1016–1026. [Google Scholar] [CrossRef]
- Li, C.; Pei, J.; Li, J.; Liu, X.; Ren, L.; Luo, Y. Overwintering larval cold tolerance of Sirex noctilio (Hymenoptera: Siricidae): Geographic variation in northeast China. Insects 2021, 12, 116. [Google Scholar] [CrossRef]
- Noor-Ul-Ane, M.; Jung, C. Characterization of cold tolerance of immature stages of small hive beetle (SHB) Aethina tumida Murray (Coleoptera: Nitidulidae). Insects 2021, 12, 459. [Google Scholar] [CrossRef]
- Andreadis, S.S.; Athanassiou, C.G. A review of insect cold hardiness and its potential in stored product insect control. Crop Prot. 2017, 91, 93–99. [Google Scholar] [CrossRef]
- Pourani, M.S.; Mahdian, K.; Izadi, H.; Basirat, M.; Sahhafi, S.R. Cold tolerance and supercooling points of two ladybird beetles (Col.: Coccinellidae): Impact of the diet. Cryobiology 2019, 91, 61–68. [Google Scholar] [CrossRef] [PubMed]
- Ditrich, T. Supercooling point is an individually fixed metric of cold tolerance in Pyrrhocoris apterus. J. Therm. Biol. 2018, 74, 208–213. [Google Scholar] [CrossRef] [PubMed]
- Worland, M.R.; Convey, P.; Luke ov, A. Rapid cold hardening: A gut feeling. Cryo Lett. 2000, 21, 315–324. [Google Scholar]
- Terblanche, J.S.; Hoffmann, A.A.; Mitchell, K.A.; Rako, L.; Le Roux, P.C.; Chown, S.L. Ecologically relevant measures of tolerance to potentially lethal temperatures. J. Exp. Biol. 2011, 214, 3713–3725. [Google Scholar] [CrossRef] [Green Version]
- Pieterse, W.; Terblanche, J.S.; Addison, P. Do thermal tolerances and rapid thermal responses contribute to the invasion potential of Bactrocera dorsalis (Diptera: Tephritidae)? J. Insect Physiol. 2017, 98, 1–6. [Google Scholar] [CrossRef] [Green Version]
- Bastola, A.; Davis, J.A. Cold tolerance and supercooling capacity of the redbanded stink bug (Hemiptera: Pentatomidae). Environ. Entomol. 2018, 47, 133–139. [Google Scholar] [CrossRef]
- Jing, X.H.; Kang, L. Research progress in insect cold hardiness. Acta Ecol. Sin. 2002, 22, 2202–2207. [Google Scholar]
- Dou, Q.C.; Liu, X.F.; Yao, W.F.; Ye, H. Tolerance of adult guava fruit fly (Diptera: Tephritidae) to low temperature. Southwest China J. Agric. Sci. 2011, 24, 1771–1774. [Google Scholar]
- Kirk Green, C.; Moore, P.J.; Sial, A.A. Impact of heat stress on development and fertility of Drosophila suzukii Matsumura (Diptera: Drosophilidae). J. Insect Physiol. 2019, 114, 45–52. [Google Scholar] [CrossRef]
- Eben, A.; Reifenrath, M.; Briem, F.; Pink, S.; Vogt, H. Response of Drosophila suzukii (Diptera: Drosophilidae) to extreme heat and dryness. Agric. Forest Entomol. 2018, 20, 113–121. [Google Scholar] [CrossRef] [Green Version]
- Mutamiswa, R.; Machekano, H.; Chidawanyika, F.; Nyamukondiwa, C. Life-stage related responses to combined effects of acclimation temperature and humidity on the thermal tolerance of Chilo partellus (Swinhoe) (Lepidoptera: Crambidae). J. Therm. Biol. 2019, 79, 85–94. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Ma, Z.; Cui, F.; Wang, X.; Guo, W.; Lin, Z.; Yang, P.; Kang, L. Parental phase status affects the cold hardiness of progeny eggs in locusts. Funct. Ecol. 2012, 26, 379–389. [Google Scholar] [CrossRef]
- Mohammadzadeh, M.; Izadi, H. Cooling rate and starvation affect supercooling point and cold tolerance of the khapra beetle, Trogoderma granarium Everts fourth instar larvae (Coleoptera: Dermestidae). J. Therm. Biol. 2018, 71, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Nedved, O.; Hodkova, M.; Brunnhofer, V.; Hodek, I. Simultaneous measurement of low temperatures survival and supercooling in a sample of insects. Cryo Lett. 1995, 16, 108–113. [Google Scholar]
- Hodkova, M.; Hodek, I. Temperature regulation of supercooling and gut nucleation in relation to diapause of Pyrrhocoris apterus (L.) (Heteroptera). Cryobiology 1997, 34, 70–79. [Google Scholar] [CrossRef] [PubMed]
- Ding, J.T.; Adil, S.; Cheng, X.T.; Huang, X.Y.; Amina. Supercooling points and freezing points of Carpomya vesuviana Costa. Acta Agr. Boreali-Occident. Sin. 2014, 23, 163–167. [Google Scholar]
- Huang, N.N.; Dai, P.; Fu, Y.G.; Huang, Q.Y. Measurement of physiological indices of cold tolerance in Bactrocera cucurbitae (Coquillett). Chin. J. Appl. Entomol. 2015, 52, 694–701. [Google Scholar]
- Hou, B.H.; Zhang, R.J. Supercooling capacity of the Oriental fruit fly, Bactrocera dorsalis (Hendel) (Diptera: Tephritidae). Entomol. Sin. 2007, 50, 638–643. [Google Scholar]
- Kost’ál, V.; Slachta, M.; Simek, P. Cryoprotective role of polyols independent of the increase in supercooling capacity in diapausing adults of Pyrrhocoris apterus (Heteroptera: Insecta). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2001, 130, 365–374. [Google Scholar] [CrossRef]
- Worland, M.R.; Leinaas, H.P.; Chown, S.L. Supercooling point frequency distributions in Collembola are affected by moulting. Funct. Ecol. 2006, 20, 323–329. [Google Scholar] [CrossRef]
- Mohammadzadeh, M.; Izadi, H. Cold acclimation of Trogoderma granarium everts is tightly linked to regulation of enzyme activity, energy content, and ion concentration. Front. Physiol. 2018, 9, 1427. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, S.Z.; Li, Q.; Feng, C.H.; Zhang, M.; Jiang, F.; Yang, G.; Luo, L.M. Cold-hardiness of Locusta migratoria tibetensis in each developmental stage. Chin. Bull. Entomol. 2007, 44, 797–798. [Google Scholar]
- Tang, Z.Z.; Wang, J.; Liu, X.R.; Li, Y.Q.; Lv, B.Q.; Peng, Z.Q.; Yu, Y.H.; Liu, M.W. Comparison of cold tolerance in two tropical invasive pests. Plant Quar. 2016, 30, 40–44. [Google Scholar]
- Ditrich, T.; Koštál, V. Comparative analysis of overwintering physiology in nine species of semi-aquatic bugs (Heteroptera: Gerromorpha). Physiol. Entomol. 2011, 36, 261–270. [Google Scholar] [CrossRef]
- Bale, J.S. Insect cold hardiness: A matter of life and death. Eur. J. Entomol. 1996, 93, 369–382. [Google Scholar]
- Parmesan, C.; Yohe, G. A globally coherent fingerprint of climate change impacts across natural systems. Nature 2003, 421, 37–42. [Google Scholar] [CrossRef]
- Cannon, R.J. The implications of predicted climate change for insect pests in the UK, with emphasis on non-indigenous species. Global. Chang. Biol. 1998, 4, 785–796. [Google Scholar] [CrossRef]
- Ragland, G.J.; Kingsolver, J.G. The effect of fluctuating temperatures on ectotherm life-history traits: Comparisons among geographic populations of Wyeomyia smithii. Evol. Ecol. Res. 2008, 10, 29–44. [Google Scholar]
- Rall, B.C.; Vucic-Pestic, O.; Ehnes, R.B.; Emmerson, M.; Brose, U. Temperature, predator-prey interaction strength and population stability. Global. Chang. Biol. 2010, 16, 2145–2157. [Google Scholar] [CrossRef]
- Vargas, R.I.; Walsh, W.A.; Kanehisa, D.; Stark, J.D.; Nishida, T. Comparative demography of three Hawaiian fruit flies (Diptera: Tephritidae) at alternating temperatures. Ann. Entomol. Soc. Am. 2000, 93, 75–81. [Google Scholar] [CrossRef]
- Gibert, P.; Moreteau, B.; Pétavy, G.; Karan, D.; David, J.R. Chill-coma tolerance, a major climatic adaptation among Drosophila species. Evolution 2001, 55, 1063–1068. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Sørensen, J.G.; Loeschcke, V. Adaptation of Drosophila to temperature extremes: Bringing together quantitative and molecular approaches. J. Therm. Biol. 2003, 28, 175–216. [Google Scholar] [CrossRef]
- Mwando, N.L.; Ndlela, S.; Meyhöfer, R.; Subramanian, S.; Mohamed, S.A. Hot water treatment for post-harvest disinfestation of Bactrocera dorsalis (Diptera: Tephritidae) and its effect on cv. tommy atkins mango. Insects 2021, 12, 1070. [Google Scholar] [CrossRef] [PubMed]
- Follett, P.A.; Manoukis, N.C.; Mackey, B. Comparative cold tolerance in Ceratitis capitata and Zeugodacus cucurbitae (Diptera: Tephritidae). J. Econ. Entomol. 2018, 111, 2632–2636. [Google Scholar] [CrossRef]
- Turnock, W.J.; Lamb, R.J.; Bodnaryk, R.P. Effects of cold stress during pupal diapause on the survival and development of Mamestra configurata (Lepidoptera: Noctuidae). Oecologia 1983, 56, 185–192. [Google Scholar] [CrossRef]
- Nedved, O. Modelling the relationship between cold injury and accumulated degree days in terrestial arthropods. Cryo Lett. 1998, 19, 267–274. [Google Scholar]
- Zhang, W.; Chang, X.Q.; Hoffmann, A.; Zhang, S.; Ma, C.S. Impact of hot events at different developmental stages of a moth: The closer to adult stage, the less reproductive output. Sci. Rep. 2015, 5, 10436. [Google Scholar] [CrossRef] [Green Version]
- Zhou, C.Q.; Wu, K.K.; Chen, H.D.; Yang, P.J.; Dowell, R.V. Effect of temperature on the population growth of Bactrocera tau (Walker) (Dipt., Tephritidae). J. Appl. Entomol. 1994, 117, 332–337. [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Developmental Stages | Supercooling Points (SCPs) | |
|---|---|---|
| Mean SCP ± SE (°C) | Range (°C) (Min., Max.) | |
| Egg | −25.82 ± 0.51 | −28.1, −14.8 |
| 1st instar larvae | −7.07 ± 0.61 | −13.0, −1.1 |
| 2nd instar larvae | −12.13 ± 0.93 | −20.3, −4.6 |
| 3rd instar larvae | −13.36 ± 0.87 | −22.0, −4.0 |
| 1-day-old pupae | −9.77 ± 0.49 | −13.2, −3.5 |
| 2-day-old pupae | −11.63 ± 0.78 | −21.0, −2.5 |
| 3-day-old pupae | −16.69 ± 0.66 | −20.9, −3.9 |
| 4-day-old pupae | −18.22 ± 0.70 | −26.4, −11.9 |
| 5-day-old pupae | −18.97 ± 0.88 | −27.1, −8.0 |
| 6-day-old pupae | −22.95 ± 0.34 | −26.1, −18.8 |
| 7-day-old pupae | −19.78 ± 0.60 | −25.4, −2.2 |
| 8-day-old pupae | −18.79 ± 0.47 | −23.7, −12.3 |
| 2–3-day-old female adult | −7.92 ± 0.49 | −13.7, −3.8 |
| 2–3-day-old male adult | −9.99 ± 0.83 | −18.5, −1.8 |
| 8–10-day-old female adult | −14.63 ± 0.78 | −22.0, −4.1 |
| 8–10-day-old male adult | −15.69 ± 0.90 | −25.5, −7.4 |
| 25–30-day-old female adult | −11.35 ± 1.01 | −20.3, −4.4 |
| 25–30-day-old male adult | −12.92 ± 0.72 | −20.7, −0.4 |
| 45–50-day-old female adult | −7.69 ± 0.72 | −23.6, −3.7 |
| 45–50-day-old male adult | −11.01 ± 0.79 | −15.8, −2.5 |
| 65–70-day-old female adult | −8.77 ± 0.76 | −16.2, −2.3 |
| 65–70-day-old male adult | −8.81 ± 0.69 | −19.7, −3.8 |
| 85–90-day-old female adult | −8.36 ± 0.63 | −15.2, −2.8 |
| 85–90-day-old male adult | −9.04 ± 0.87 | −18.6, −3.1 |
| 105–110-day-old female adult | −8.72 ± 0.65 | −16.6, −3.0 |
| 105–110-day-old male adult | −9.78 ± 0.96 | −22.1, −2.3 |
| Life Stages | The Semi-Lethal Temperature (LLT50) (°C) | |||
|---|---|---|---|---|
| 0.5 h | 1 h | 2 h | 4 h | |
| Egg | −16.39 | −13.96 | −4.66 | −2.40 |
| 1st instar larvae | −4.88 | −3.15 | −1.68 | −0.86 |
| 2nd instar larvae | −4.23 | −3.39 | −1.84 | −1.43 |
| 3rd instar larvae | −4.43 | −3.87 | −1.97 | −1.62 |
| 1-day-old pupae | −5.27 | −2.60 | −2.11 | −1.51 |
| 3-day-old adults | −3.69 | −2.46 | −1.80 | −1.44 |
| Life Stages | The Semi-Lethal Temperature (LLT50) (°C) | |||
|---|---|---|---|---|
| 0.5 h | 1 h | 2 h | 4 h | |
| Egg | 39.99 | 37.43 | 31.51 | 27.35 |
| 1st instar larvae | 41.15 | 40.91 | 39.61 | 39.41 |
| 2nd instar larvae | 41.92 | 40.99 | 40.74 | 40.28 |
| 3rd instar larvae | 45.23 | 41.83 | 41.46 | 40.74 |
| 1-day-old pupae | 43.90 | 42.69 | 40.74 | 38.97 |
| 3-day-old adults | 42.63 | 41.06 | 40.48 | 38.22 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Wang, X.; Chen, Z.; Lu, Y. Characterization of Cold and Heat Tolerance of Bactrocera tau (Walker). Insects 2022, 13, 329. https://doi.org/10.3390/insects13040329
Liu H, Wang X, Chen Z, Lu Y. Characterization of Cold and Heat Tolerance of Bactrocera tau (Walker). Insects. 2022; 13(4):329. https://doi.org/10.3390/insects13040329
Chicago/Turabian StyleLiu, Huan, Xiaoyan Wang, Zihan Chen, and Yongyue Lu. 2022. "Characterization of Cold and Heat Tolerance of Bactrocera tau (Walker)" Insects 13, no. 4: 329. https://doi.org/10.3390/insects13040329