
Midgut and Head Transcriptomic Analysis of Silkworms Reveals the Physiological Effects of Artificial Diets


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation of ML and AD
2.2. Rearing of Silkworms
2.3. RNA Extraction, cDNA Library Preparation and Sequencing
2.4. Transcriptome Data Quality Controls and Alignment
2.5. Bioinformatic and Computational Analysis
2.6. RNA Reverse Transcription and qRT-PCR of DEGs
3. Results
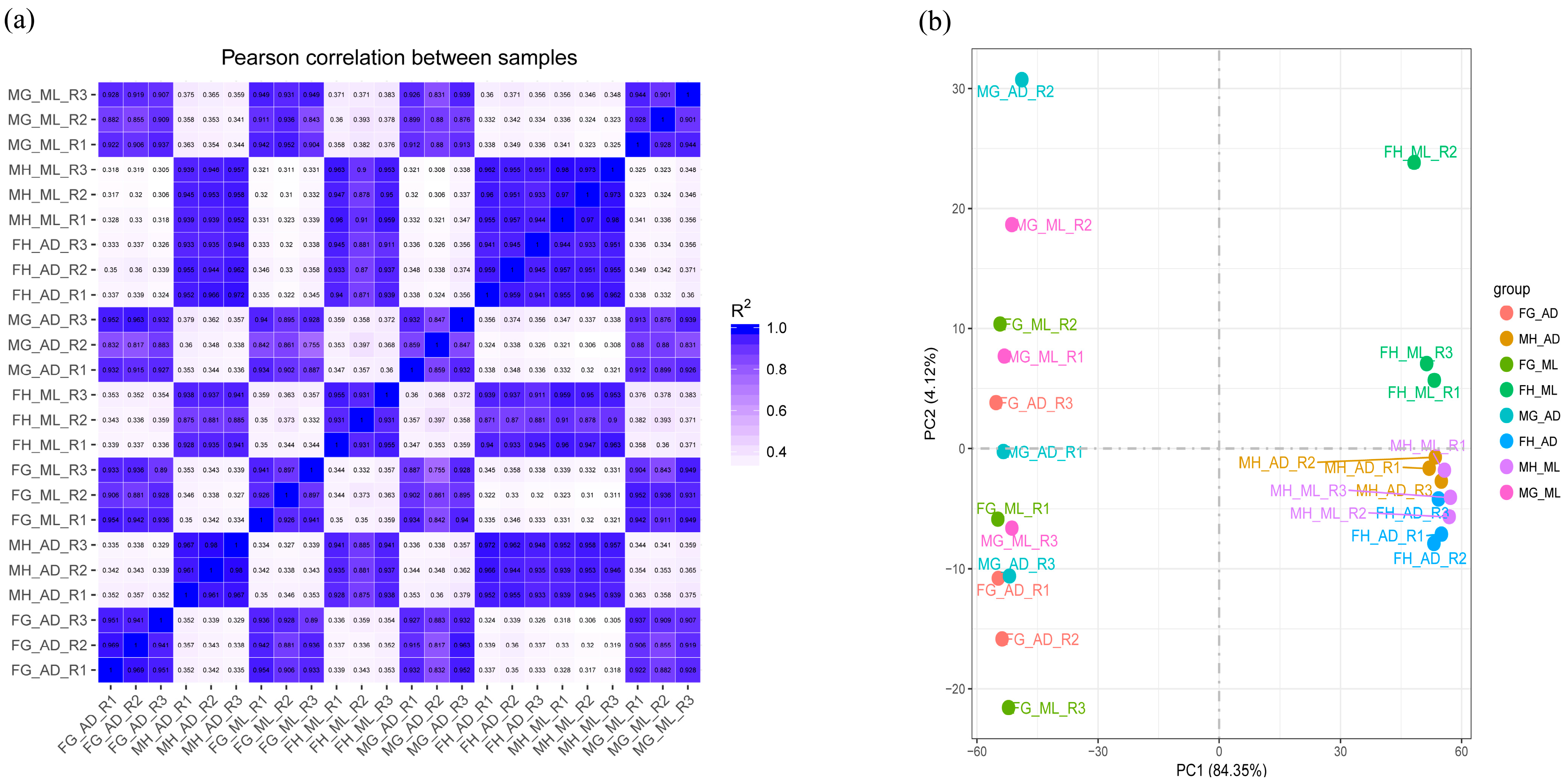
3.1. Phenotypic Observations and Sequencing Data
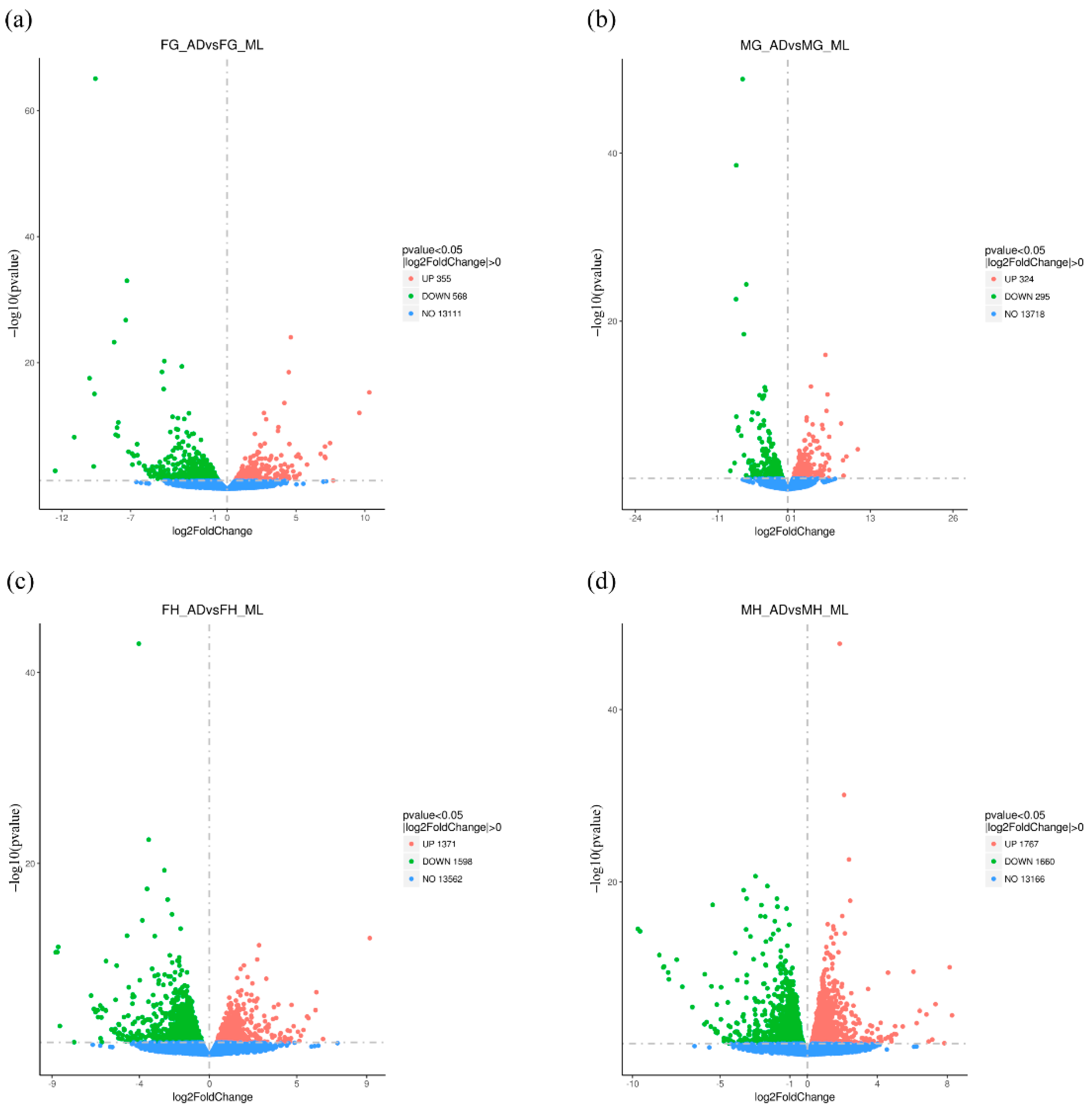
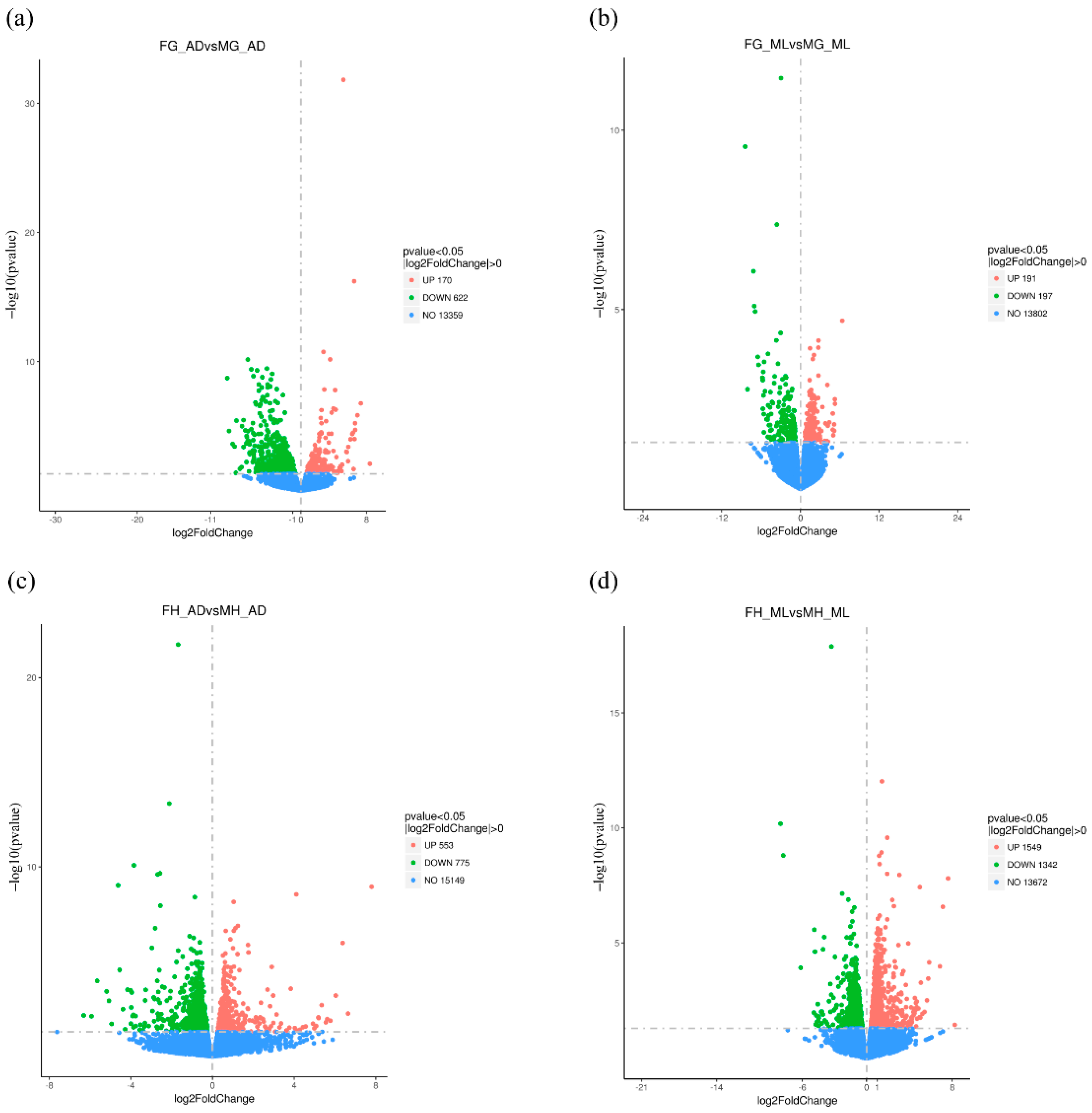
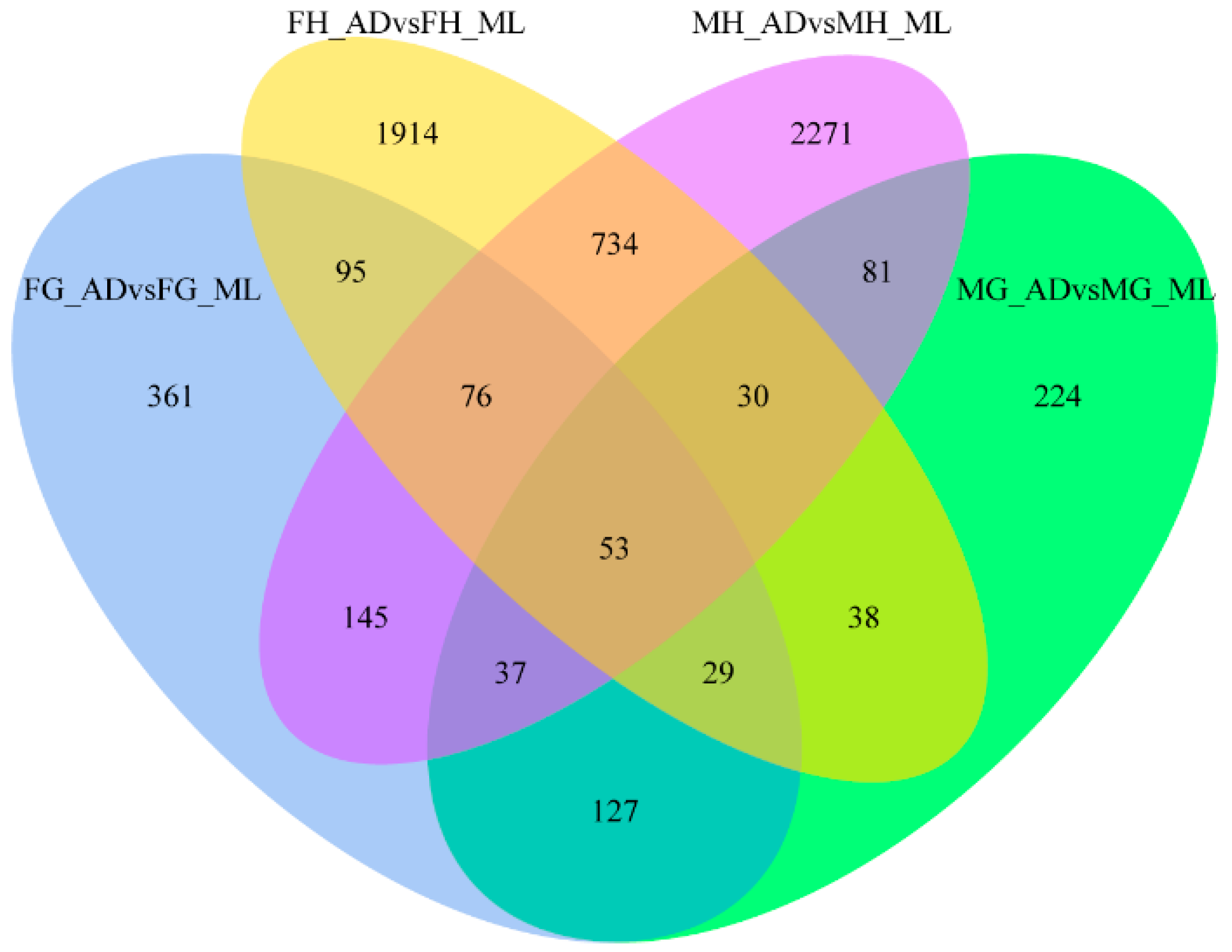
3.2. Global Differential Gene Expression Changes
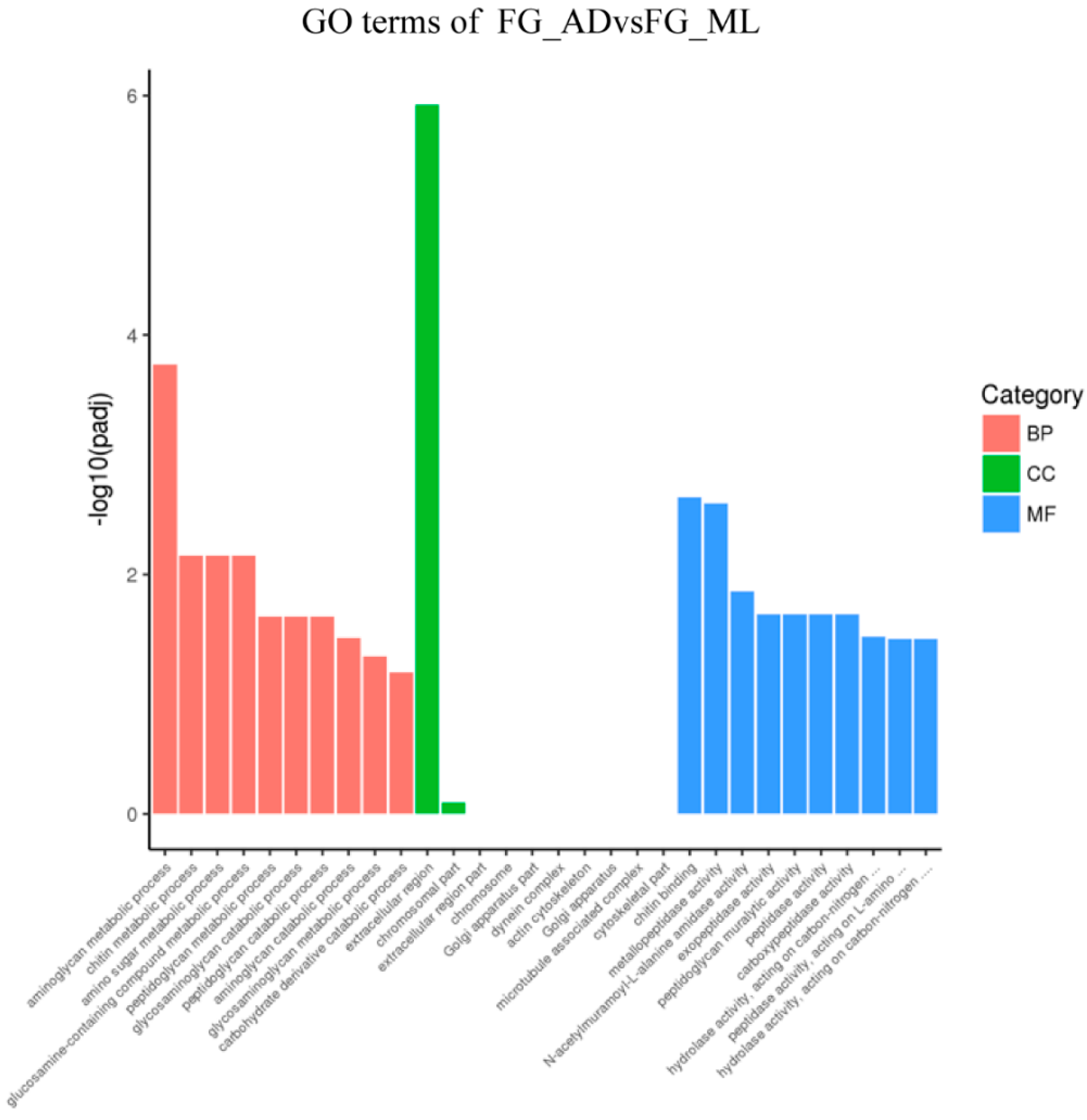
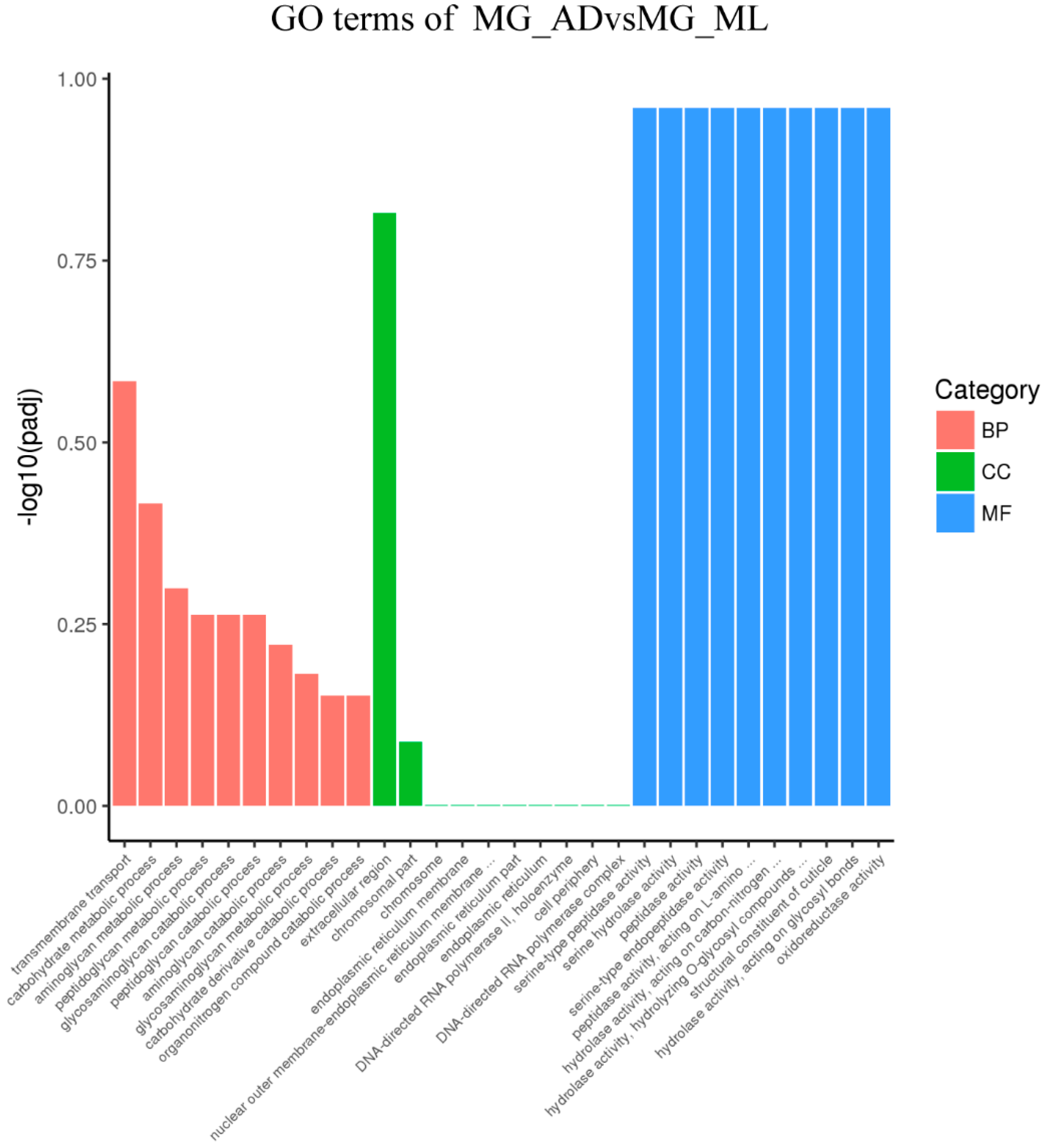
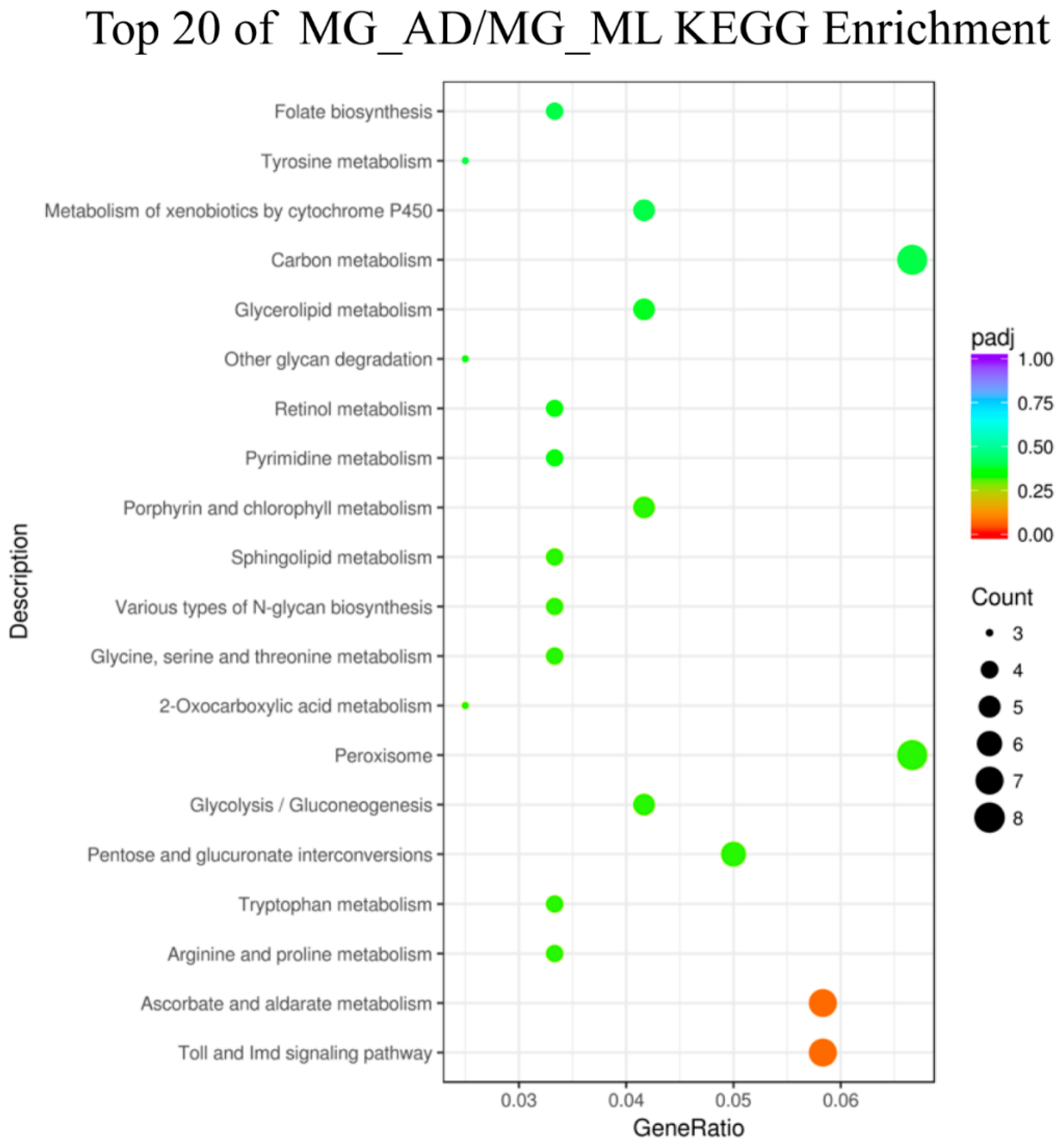
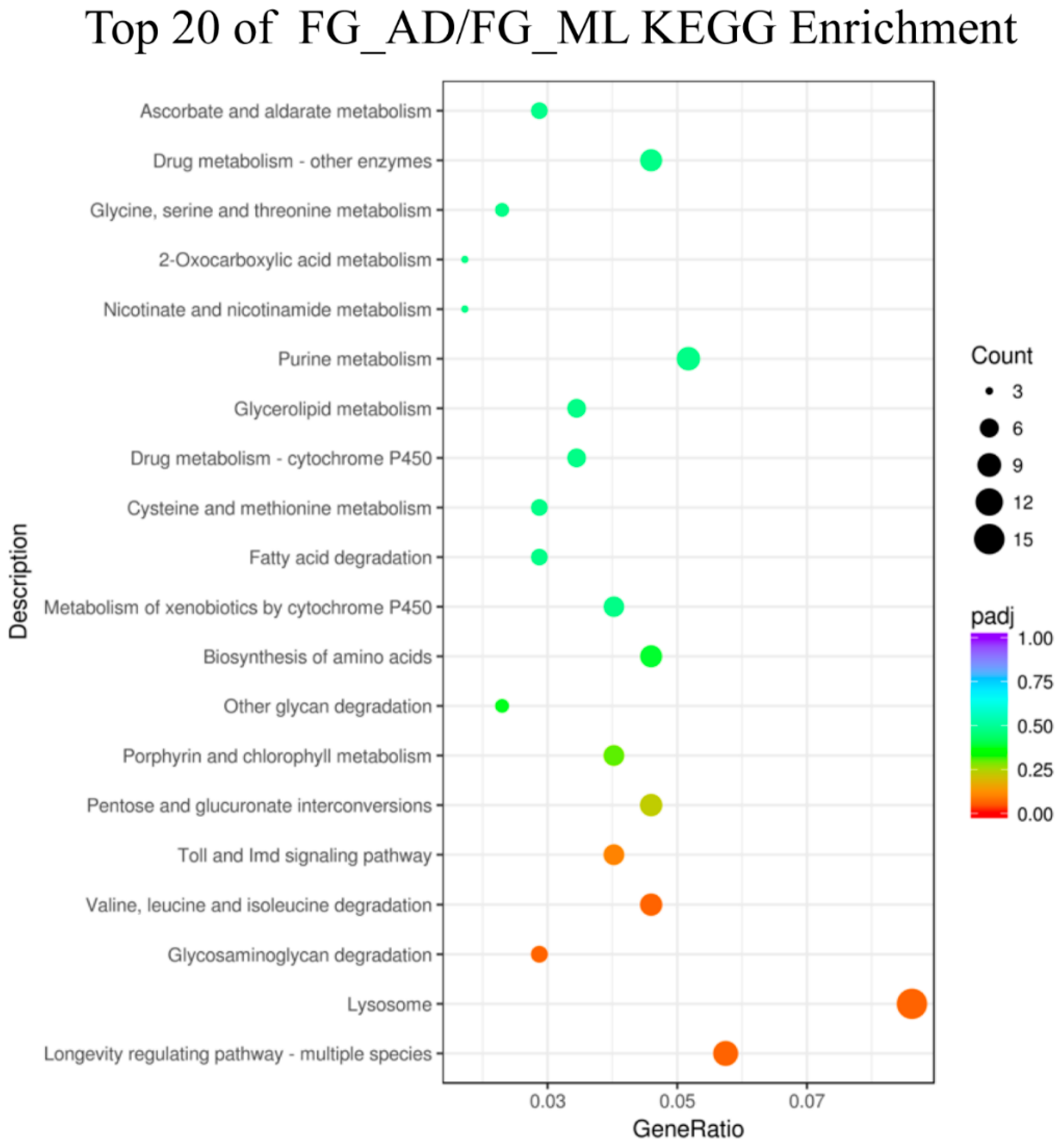
3.3. Genes Associated with Digestion and Absorption of Nutrients
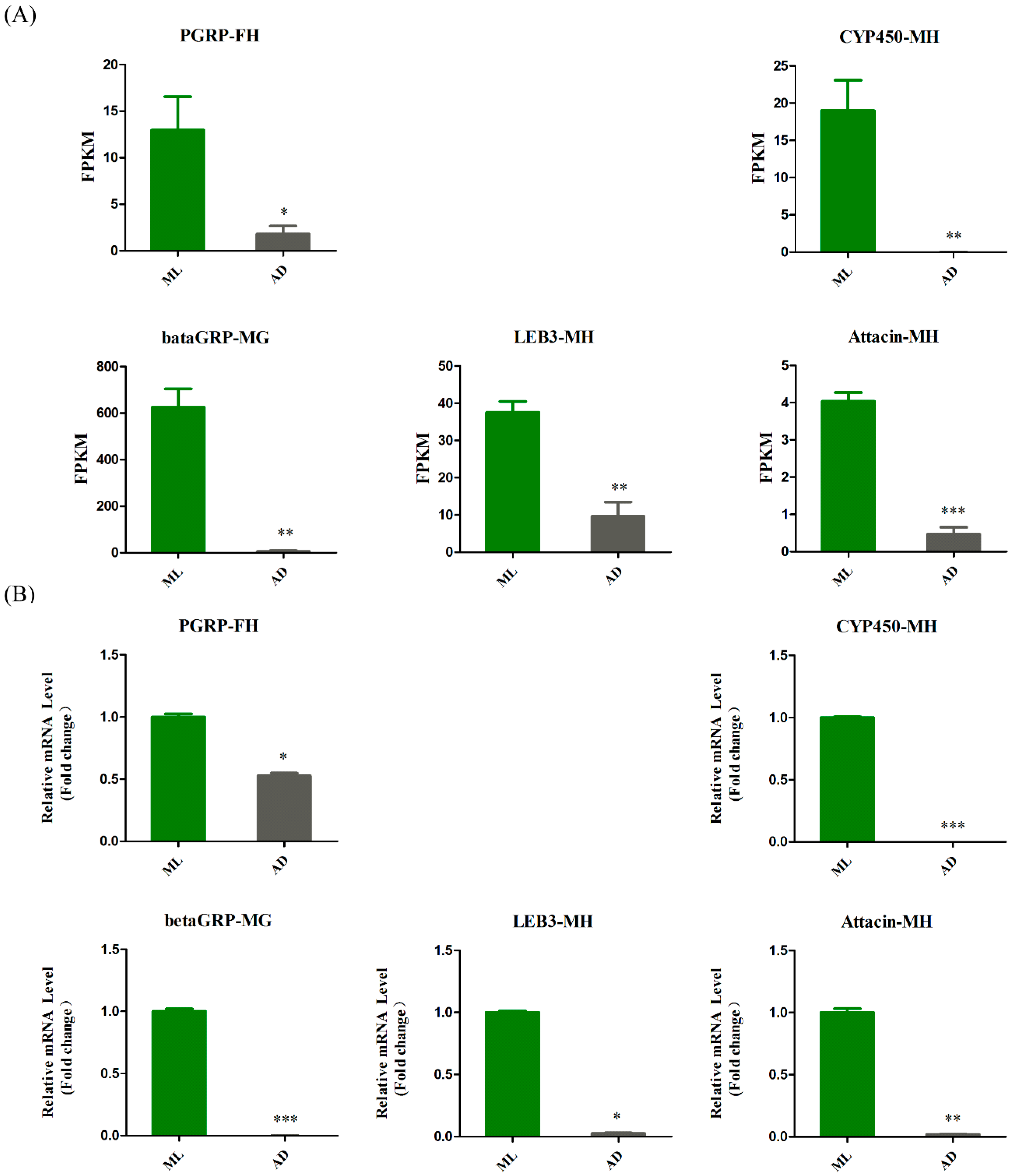
3.4. Genes Associated with Innate Immunity
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Xia, Q.; Guo, Y.; Zhang, Z.; Li, D.; Xuan, Z.; Li, Z.; Dai, F.; Li, Y.; Cheng, D.; Li, R.; et al. Complete Resequencing of 40 Genomes Reveals Domestication Events and Genes in Silkworm (Bombyx). Science 2009, 326, 433–436. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wu, X.; He, K.; Velickovic, T.C.; Liu, Z. Nutritional, functional, and allergenic properties of silkworm pupae. Food Sci. Nutr. 2021, 9, 4655–4665. [Google Scholar] [CrossRef] [PubMed]
- Hamamura, Y.; Matsuura, K.; Nishida, J.; Hayashiya, K.; Naito, K.I. Food selection by silkworm larvae. Nature 1962, 194, 754–755. [Google Scholar] [CrossRef]
- Kumar, H.; Priya, Y.S.; Kumar, M.; Elangovan, V. Effect of Different Mulberry Varieties and Seasons on Growth and Economic Traits of Bivoltine Silkworm (Bombyx mori). J. Entomol. 2013, 10, 147–155. [Google Scholar] [CrossRef] [Green Version]
- Cappellozza, L.; Cappellozza, S.; Saviane, A.; Sbrenna, G. Artificial diet rearing system for the silkworm Bombyx mori (Lepidoptera: Bombycidae): Effect of vitamin C deprivation on larval growth and cocoon production. Appl. Entomol. Zool. 2005, 40, 405–412. [Google Scholar] [CrossRef] [Green Version]
- Zhou, Z.H.; Yang, H.J.; Chen, M.; Lou, C.F.; Zhang, Y.Z.; Chen, K.P.; Wang, Y.; Yu, M.L.; Yu, F.; Li, J.Y.; et al. Comparative proteomic analysis between the domesticated silkworm (Bombyx mori) reared on fresh mulberry leaves and on artificial diet. J. Proteome Res. 2008, 7, 5103–5111. [Google Scholar] [CrossRef]
- Hegedus, D.; Erlandson, M.; Gillott, C.; Toprak, U. New insights into peritrophic matrix synthesis, architecture, and function. Annu. Rev. Entomol. 2009, 54, 285–302. [Google Scholar] [CrossRef]
- Wu, K.; Yang, B.; Huang, W.; Dobens, L.; Song, H.; Ling, E. Gut immunity in Lepidopteran insects. Dev. Comp. Immunol. 2016, 64, 65–74. [Google Scholar] [CrossRef]
- Weiss, B.L.; Savage, A.F.; Griffith, B.C.; Wu, Y.; Aksoy, S. The peritrophic matrix mediates differential infection outcomes in the tsetse fly gut following challenge with commensal, pathogenic, and parasitic microbes. J. Immunol. 2014, 193, 773–782. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huang, F.; Wu, X. Brain Neurotransmitter Modulation by Gut Microbiota in Anxiety and Depression. Front. Cell Dev. Biol. 2021, 9, 649103. [Google Scholar] [CrossRef]
- Needham, B.D.; Funabashi, M.; Adame, M.D.; Wang, Z.; Boktor, J.C.; Haney, J.; Wu, W.L.; Rabut, C.; Ladinsky, M.S.; Hwang, S.J.; et al. A gut-derived metabolite alters brain activity and anxiety behaviour in mice. Nature 2022, 602, 647–653. [Google Scholar] [CrossRef] [PubMed]
- Konala, N.; Abburi, P.; Bovilla, V.R.; Mamillapalli, A. The effect of bovine milk on the growth of Bombyx mori. J. Insect. Sci. 2013, 13, 7. [Google Scholar] [CrossRef] [PubMed]
- Hamamura, Y.; Kuwata, K.; Masuda, H. Effect of gallic acid on growth of the silkworm larvae Bombyx mori L. Nature 1966, 212, 1386–1387. [Google Scholar] [CrossRef]
- Horie, Y.; Nakasone, S. Effects of levels of fatty acids and carbohydrates in a diet on biosynthesis of fatty acids in larvae of silkworm, Bombyx mori. J. Insect Physiol. 1971, 17, 1441–1450. [Google Scholar] [CrossRef]
- Weizheng, C.; Shengxiang, Z.; Qingxin, L.; Yanwen, W.; Hongli, W.; Xunli, L.; Zhimei, M. An overview on research and commercialization of artificial diet for silkworm (Bombyx mori) in China. Sci. Seric. 2016, 42, 3–15. [Google Scholar]
- Qin, D.; Wang, G.; Dong, Z.; Xia, Q.; Zhao, P. Comparative Fecal Metabolomes of Silkworms Being Fed Mulberry Leaf and Artificial Diet. Insects 2020, 11, 851. [Google Scholar] [CrossRef]
- Lamberti, C.; Gai, F.; Cirrincione, S.; Giribaldi, M.; Purrotti, M.; Manfredi, M.; Marengo, E.; Sicuro, B.; Saviane, A.; Cappellozza, S.; et al. Investigation of the protein profile of silkworm (Bombyx mori) pupae reared on a well-calibrated artificial diet compared to mulberry leaf diet. PeerJ. 2019, 7, e6723. [Google Scholar] [CrossRef] [Green Version]
- Feng, L.C.; Sheng, W.D. Anatomy and Physiology of Silkworm, 1st ed.; Higher Education Press: Beijing, China, 2015; pp. 244–246. [Google Scholar]
- Qin, L.; Shi, H.; Xia, H.; Chen, L.; Yao, Q.; Chen, K. Comparative proteomic analysis of midgut proteins from male and female Bombyx mori (Lepidoptera: Bombycidae). J. Insect. Sci. 2014, 14, 226. [Google Scholar] [CrossRef] [Green Version]
- Anders, S.; Huber, W. Differential expression analysis for sequence count data. Genome Biol. 2010, 11, R106. [Google Scholar] [CrossRef] [Green Version]
- Robinson, M.D.; McCarthy, D.J.; Smyth, G.K. edgeR: A Bioconductor package for differential expression analysis of digital gene expression data. Bioinformatics 2010, 26, 139–140. [Google Scholar] [CrossRef] [Green Version]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Dziarski, R.; Gupta, D. A Balancing Act: PGRPs Preserve and Protect. Cell Host Microbe 2018, 23, 149–151. [Google Scholar] [CrossRef] [Green Version]
- Takehana, A.; Yano, T.; Mita, S.; Kotani, A.; Oshima, Y.; Kurata, S. Peptidoglycan recognition protein (PGRP)-LE and PGRP-LC act synergistically in Drosophila immunity. Embo J. 2004, 23, 4690–4700. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Charroux, B.; Capo, F.; Kurz, C.L.; Peslier, S.; Chaduli, D.; Viallat-Lieutaud, A.; Royet, J. Cytosolic and Secreted Peptidoglycan-Degrading Enzymes in Drosophila Respectively Control Local and Systemic Immune Responses to Microbiota. Cell Host Microbe 2018, 23, 215–228.e214. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaidman-Rémy, A.; Hervé, M.; Poidevin, M.; Pili-Floury, S.; Kim, M.S.; Blanot, D.; Oh, B.H.; Ueda, R.; Mengin-Lecreulx, D.; Lemaitre, B. The Drosophila amidase PGRP-LB modulates the immune response to bacterial infection. Immunity 2006, 24, 463–473. [Google Scholar] [CrossRef] [Green Version]
- Yoshida, H.; Kinoshita, K.; Ashida, M. Purification of a peptidoglycan recognition protein from hemolymph of the silkworm, Bombyx mori. J. Biol. Chem. 1996, 271, 13854–13860. [Google Scholar] [CrossRef] [Green Version]
- De Diego-Diaz, B.; Treu, L.; Campanaro, S.; da Silva Duarte, V.; Saviane, A.; Cappellozza, S.; Squartini, A. Genome Sequence of Enterococcus mundtii EM01, Isolated from Bombyx mori Midgut and Responsible for Flacherie Disease in Silkworms Reared on an Artificial Diet. Genome Announc. 2018, 6, e01495-17. [Google Scholar] [CrossRef] [Green Version]
- Nwibo, D.D.; Matsumoto, Y.; Sekimizu, K. Identification and methods for prevention of Enterococcus mundtii infection in silkworm larvae, Bombyx mori, reared on artificial diet. Drug Discov. Ther. 2015, 9, 184–190. [Google Scholar] [CrossRef] [Green Version]
- Liu, N.; Zhang, L. Identification of two new cytochrome P450 genes and their 5’-flanking regions from the housefly, Musca domestica. Insect. Biochem. Mol. Biol. 2002, 32, 755–764. [Google Scholar] [CrossRef]
- Gu, Z.Y.; Sun, S.S.; Wang, Y.H.; Wang, B.B.; Xie, Y.; Ma, L.; Wang, J.M.; Shen, W.D.; Li, B. Transcriptional characteristics of gene expression in the midgut of domestic silkworms (Bombyx mori) exposed to phoxim. Pestic. Biochem. Physiol. 2013, 105, 36–43. [Google Scholar] [CrossRef] [PubMed]
- Bartling, M.T.; Thumecke, S.; Russert, J.H.; Vilcinskas, A.; Lee, K.Z. Exposure to low doses of pesticides induces an immune response and the production of nitric oxide in honeybees. Sci. Rep. 2021, 11, 6819. [Google Scholar] [CrossRef]
- Ochiai, M.; Ashida, M. A pattern-recognition protein for beta-1,3-glucan. The binding domain and the cDNA cloning of beta-1,3-glucan recognition protein from the silkworm, Bombyx mori. J. Biol. Chem. 2000, 275, 4995–5002. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Takahasi, K.; Ochiai, M.; Horiuchi, M.; Kumeta, H.; Ogura, K.; Ashida, M.; Inagaki, F. Solution structure of the silkworm betaGRP/GNBP3 N-terminal domain reveals the mechanism for beta-1,3-glucan-specific recognition. Proc. Natl. Acad. Sci. USA 2009, 106, 11679–11684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Mao, X.; Xu, X.; Li, Z.; Yang, J.; Liu, Z. Molecular cloning and characterization of the β-1,3-glucan recognition protein in Anatolica polita. Gene 2019, 697, 144–151. [Google Scholar] [CrossRef]
- Rayaprolu, S.; Wang, Y.; Kanost, M.R.; Hartson, S.; Jiang, H. Functional analysis of four processing products from multiple precursors encoded by a lebocin-related gene from Manduca sexta. Dev. Comp. Immunol. 2010, 34, 638–647. [Google Scholar] [CrossRef] [Green Version]
- Lü, D.; Hou, C.; Qin, G.; Gao, K.; Chen, T.; Guo, X. Molecular Cloning, Bioinformatic Analysis, and Expression of Bombyx mori Lebocin 5 Gene Related to Beauveria bassiana Infection. Biomed Res. Int. 2017, 2017, 9390803. [Google Scholar] [CrossRef] [Green Version]
- Chowdhury, S.; Taniai, K.; Hara, S.; Kadono-Okuda, K.; Kato, Y.; Yamamoto, M.; Xu, J.; Choi, S.K.; Debnath, N.C.; Choi, H.K.; et al. cDNA cloning and gene expression of lebocin, a novel member of antibacterial peptides from the silkworm, Bombyx mori. Biochem. Biophys Res. Commun. 1995, 214, 271–278. [Google Scholar] [CrossRef]
- Zhang, J.; Li, Q.; Wei, G.; Wang, L.; Qian, C.; Sun, Y.; Tian, J.; Zhu, B.; Liu, C. Identification and function of a lebocin-like gene from the Chinese oak silkworm, Antheraea pernyi. J. Invertebr. Pathol. 2019, 166, 107207. [Google Scholar] [CrossRef]
- Furukawa, S.; Taniai, K.; Ishibashi, J.; Hara, S.; Shono, T.; Yamakawa, M. A novel member of lebocin gene family from the silkworm, Bombyx mori. Biochem. Biophys Res. Commun. 1997, 238, 769–774. [Google Scholar] [CrossRef]
- Mai, T.; Chen, S.; Lin, X.; Zhang, X.; Zou, X.; Feng, Q.; Zheng, S. 20-hydroxyecdysone positively regulates the transcription of the antimicrobial peptide, lebocin, via BmEts and BmBR-C Z4 in the midgut of Bombyx mori during metamorphosis. Dev. Comp. Immunol. 2017, 74, 10–18. [Google Scholar] [CrossRef] [PubMed]
- Tanaka, H.; Sagisaka, A.; Fujita, K.; Furukawa, S.; Ishibashi, J.; Yamakawa, M. BmEts upregulates promoter activity of lebocin in Bombyx mori. Insect. Biochem. Mol. Biol. 2012, 42, 474–481. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Zhang, J.; Sun, Y.; Wang, L.; Qian, C.; Wei, G.; Zhu, B.; Liu, C. Immunological Function of the Antibacterial Peptide Attacin-Like in the Chinese Oak Silkworm, Antheraea pernyi. Protein Pept. Lett. 2020, 27, 953–961. [Google Scholar] [CrossRef] [PubMed]
- Taniai, K.; Ishii, T.; Sugiyama, M.; Miyanoshita, A.; Yamakawa, M. Nucleotide sequence of 5’-upstream region and expression of a silkworm gene encoding a new member of the attacin family. Biochem. Biophys Res. Commun. 1996, 220, 594–599. [Google Scholar] [CrossRef]
- Kishimoto, K.; Fujimoto, S.; Matsumoto, K.; Yamano, Y.; Morishima, I. Protein purification, cDNA cloning and gene expression of attacin, an antibacterial protein, from eri-silkworm, Samia cynthia ricini. Insect. Biochem. Mol. Biol. 2002, 32, 881–887. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Category | ML | AD | ||||||
|---|---|---|---|---|---|---|---|---|
| Parameter | FG | FH | MG | MH | FG | FH | MG | MH |
| raw reads | 48,376,351 | 45,283,683 | 44,425,225 | 46,127,387 | 46,232,769 | 40,543,023 | 42,918,998 | 52,525,801 |
| clean reads | 47,594,289 | 44,735,653 | 43,798,904 | 45,544,796 | 45,407,637 | 40,039,759 | 42,469,478 | 51,984,274 |
| clean bases (bp) | 7.14 (G) | 6.71 (G) | 6.57 (G) | 6.83 (G) | 6.81 (G) | 6.01 (G) | 6.37 (G) | 7.8 (G) |
| clean reads/raw reads | 0.9838 | 0.9879 | 0.9859 | 0.9874 | 0.9822 | 0.9876 | 0.9895 | 0.9897 |
| Q20 (%) | 96.09 | 95.74 | 95.98 | 95.97 | 95.92 | 95.91 | 95.84 | 96.26 |
| Q30 (%) | 89.98 | 89.22 | 89.71 | 89.68 | 89.64 | 89.56 | 89.53 | 90.33 |
| GC (%) | 47.83 | 44.77 | 48.17 | 45.71 | 46.91 | 44.28 | 47.02 | 44.9 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, J.; Chen, C.; Zha, X. Midgut and Head Transcriptomic Analysis of Silkworms Reveals the Physiological Effects of Artificial Diets. Insects 2022, 13, 291. https://doi.org/10.3390/insects13030291
Li J, Chen C, Zha X. Midgut and Head Transcriptomic Analysis of Silkworms Reveals the Physiological Effects of Artificial Diets. Insects. 2022; 13(3):291. https://doi.org/10.3390/insects13030291
Chicago/Turabian StyleLi, Juan, Chunbing Chen, and Xingfu Zha. 2022. "Midgut and Head Transcriptomic Analysis of Silkworms Reveals the Physiological Effects of Artificial Diets" Insects 13, no. 3: 291. https://doi.org/10.3390/insects13030291