
Behavioural Responses of Male Aedes albopictus to Different Volatile Chemical Compounds
 ,
,  ,
, 
 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects
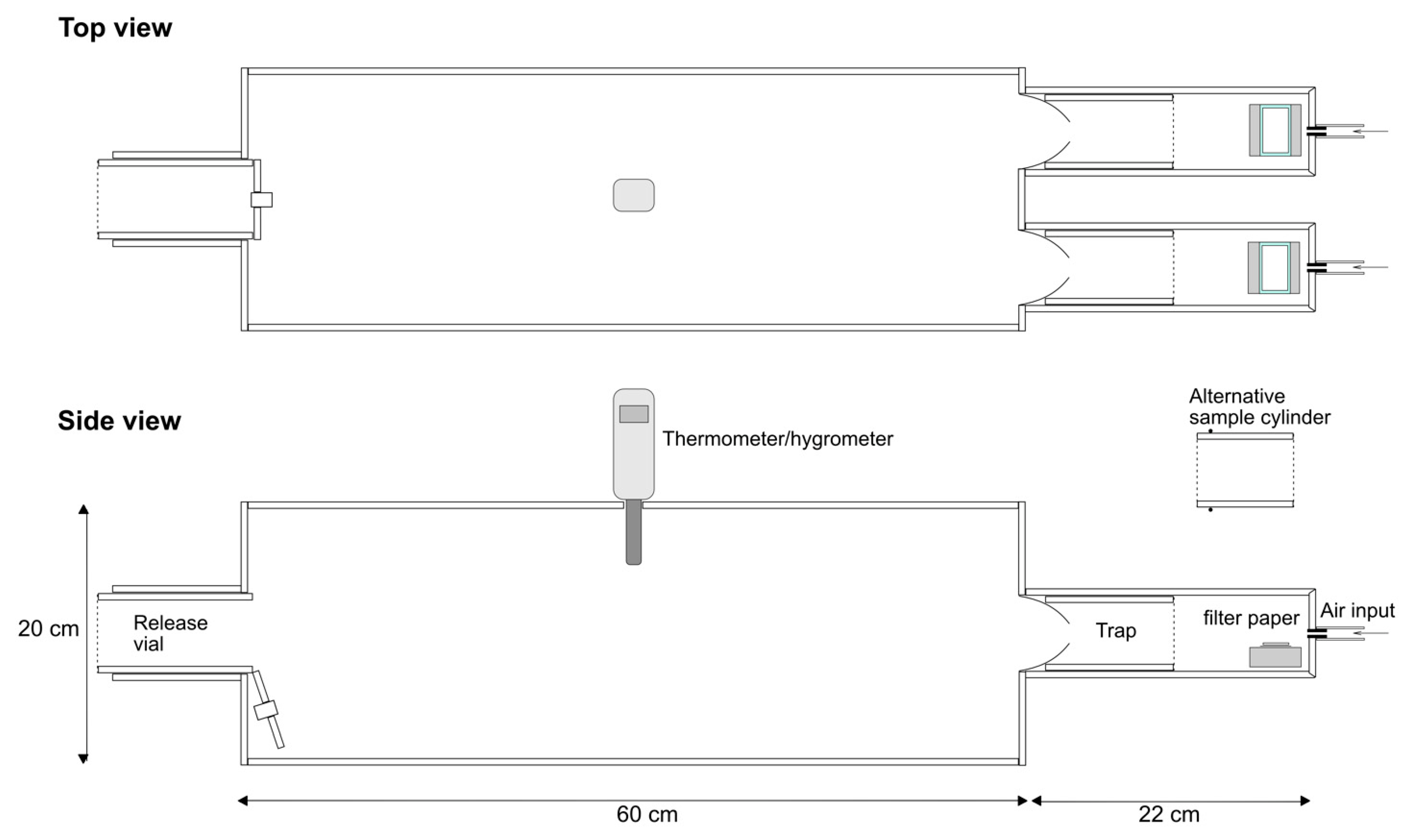
2.2. Olfactometer
2.3. Olfactometer Validation
2.4. Chemical Compounds and Preparation
3. Results
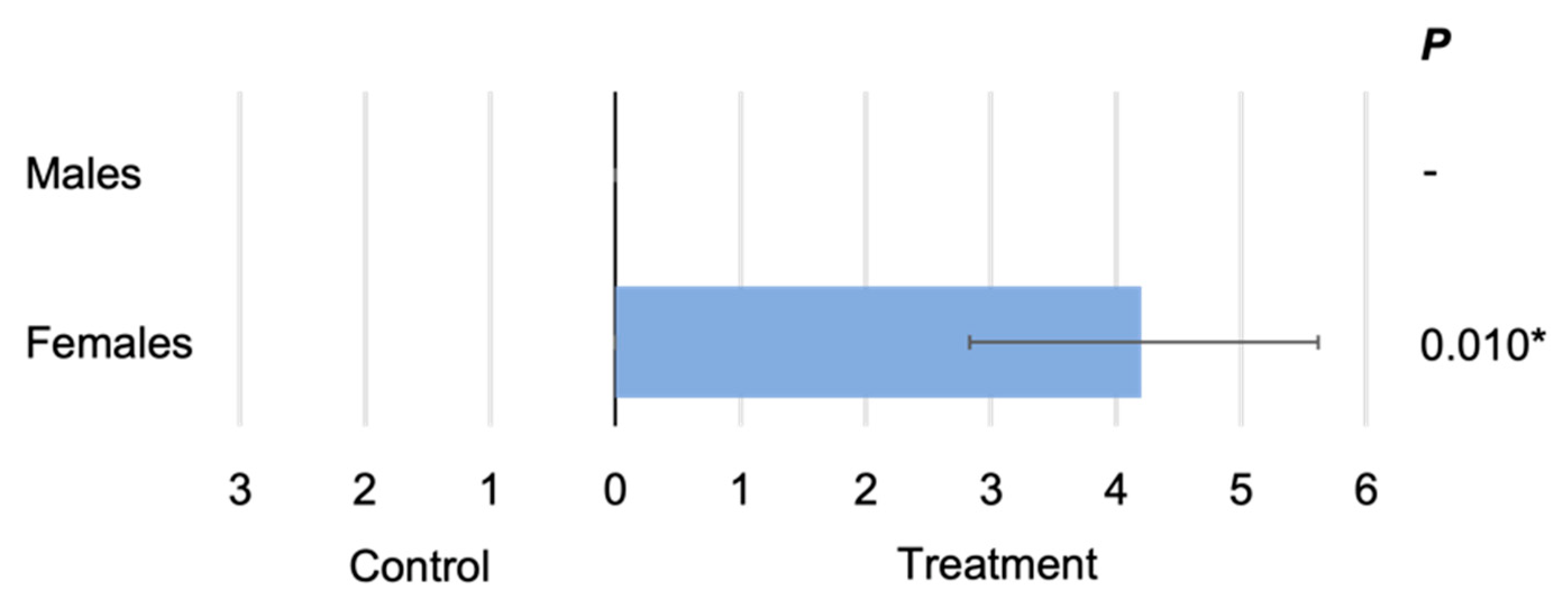
3.1. Olfactometer Validation
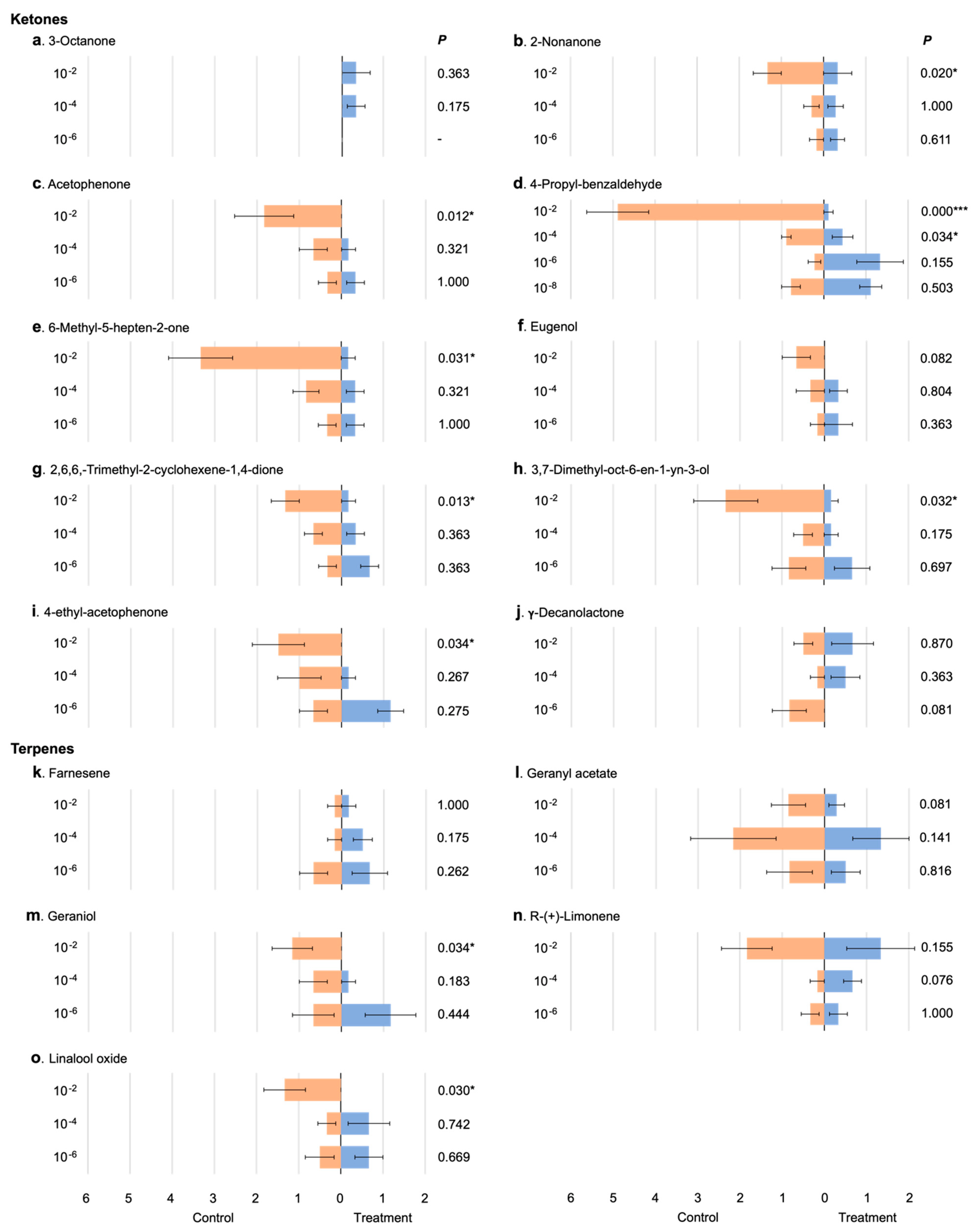
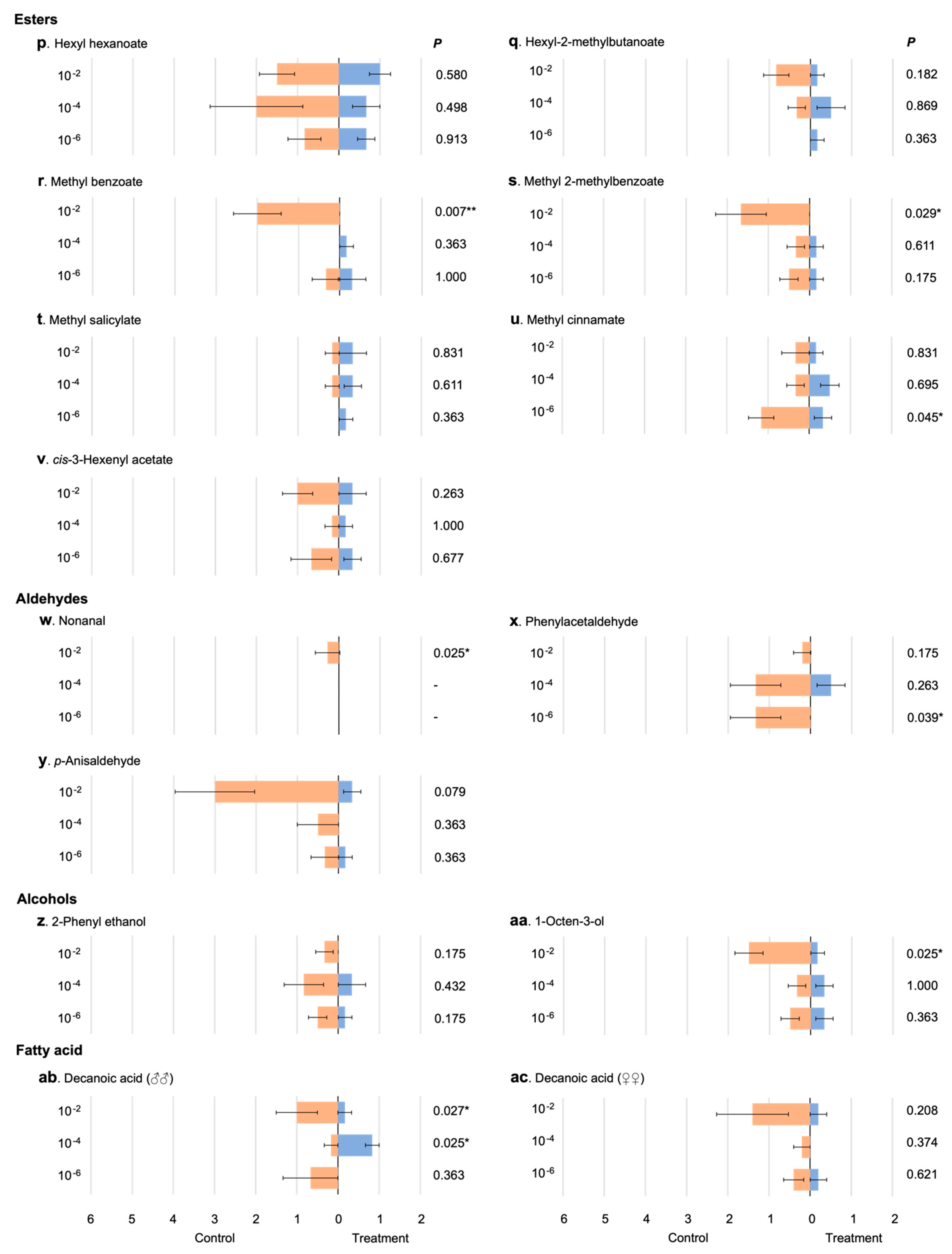
3.2. Response of Male Mosquitoes to Chemical Compounds
4. Discussion
4.1. Animal-Related Volatiles
4.2. Plant-Related Volatiles
4.3. Putative Pheromones
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pitts, R.J.; Mozūraitis, R.; Gauvin-Bialecki, A.; Lempérière, G. The roles of kairomones, synomones and pheromones in the chemically-mediated behaviour of male mosquitoes. Acta Trop. 2014, 132, S26–S34. [Google Scholar] [CrossRef]
- Lahondère, C.; Vinauger, C.; Okubo, R.P.; Wolff, G.H.; Chan, J.K.; Akbari, O.S.; Riffell, J.A. The olfactory basis of orchid pollination by mosquitoes. Proc. Natl. Acad. Sci. USA 2019, 117, 708–716. [Google Scholar] [CrossRef] [Green Version]
- Peach, D.A.H.; Gries, G. Nectar thieves or invited pollinators? A case study of tansy flowers and common house mosquitoes. Arthropod-Plant Interact. 2016, 10, 497–506. [Google Scholar] [CrossRef]
- Afify, A.; Potter, C.J. Insect repellents mediate species-specific olfactory behaviours in mosquitoes. Malar. J. 2020, 19, 127. [Google Scholar] [CrossRef] [Green Version]
- Otienoburu, P.E.; Ebrahimi, B.; Phelan, P.L.; Foster, W.A. Analysis and optimization of a synthetic Milkweed floral attractant for mosquitoes. J. Chem. Ecol. 2012, 38, 873–881. [Google Scholar] [CrossRef] [Green Version]
- Jhumur, U.S.; Dötterl, S.; Jürgens, A. Electrophysiological and behavioural responses of mosquitoes to volatiles of Silene otites (Caryophyllaceae). Arthropod-Plant Interact. 2007, 1, 245–254. [Google Scholar] [CrossRef]
- Gillies, M.T. The role of carbon dioxide in host-finding by mosquitoes (Diptera: Culicidae): A review. Bull. Entomol. Res. 1980, 70, 525–532. [Google Scholar] [CrossRef] [Green Version]
- Gubler, D.J.; Bhattacharya, N.C. Swarming and mating of Aedes (S.) albopictus in nature. Mosq. News 1972, 32, 219–223. [Google Scholar]
- Hartberg, W.K. Observations on the mating behaviour of Aedes aegypti in nature. Bull. World Health Organ. 1971, 45, 847–850. [Google Scholar]
- Wooding, M.; Naudé, Y.; Rohwer, E.; Bouwer, M. Controlling mosquitoes with semiochemicals: A review. Parasites Vectors 2020, 13, 80. [Google Scholar] [CrossRef]
- Nyasembe, V.O.; Torto, B. Volatile phytochemicals as mosquito semiochemicals. Phytochem. Lett. 2013, 8, 196–201. [Google Scholar] [CrossRef] [Green Version]
- Fawaz, E.Y.; Allan, S.A.; Bernier, U.; Obenauer, P.J.; Diclaro, J.W. Swarming mechanisms in the yellow fever mosquito: Aggregation pheromones are involved in the mating behavior of Aedes aegypti. J. Vector Ecol. 2014, 39, 347–354. [Google Scholar] [CrossRef] [PubMed]
- Hao, H.; Sun, J.; Dai, J. Dose-dependent behavioral response of the mosquito Aedes albopictus to floral odorous compounds. J. Insect Sci. 2013, 13, 127. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dormont, L.; Mulatier, M.; Carrasco, D.; Cohuet, A. Mosquito attractants. J. Chem. Ecol. 2021, 47, 351–393. [Google Scholar] [CrossRef]
- Wang, G.; Carey, A.F.; Carlson, J.R.; Zwiebel, L.J. Molecular basis of odor coding in the malaria vector mosquito Anopheles gambiae. Proc. Natl. Acad. Sci. USA 2010, 107, 4418–4423. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McBride, C.S.; Baier, F.; Omondi, A.B.; Spitzer, S.A.; Lutomiah, J.; Sang, R.; Ignell, R.; Vosshall, L.B. Evolution of mosquito preference for humans linked to an odorant receptor. Nature 2014, 515, 222–227. [Google Scholar] [CrossRef] [PubMed]
- Kline, D.L. Olfactory responses and field attraction of mosquitoes to volatiles from Limburger cheese and human foot odor. J. Vector Ecol. 1998, 23, 186–194. [Google Scholar] [PubMed]
- Qui, Y.; Smallegange, R.; Smid, H.; van Loon, J.; Galimard, A.; Posthumus, M.; van Beek, T.; Takken, W. GC-EAG analysis of human odours that attract the malaria mosquito Anopheles gambiae sensu stricto. Proc. Neth. Entomol. Soc. 2004, 15, 59–64. [Google Scholar]
- Verhulst, N.O.; Qiu, Y.T.; Beijleveld, H.; Maliepaard, C.; Knights, D.; Schulz, S.; Berg-Lyons, D.; Lauber, C.L.; Verduijn, W.; Haasnoot, G.W.; et al. Composition of human skin microbiota affects attractiveness to malaria mosquitoes. PLoS ONE 2011, 6, e28991. [Google Scholar] [CrossRef] [PubMed]
- Sokal, R.R.; Rohlf, F.J. Biometry, 2nd ed.; W. H. Freeman and Company: San Francisco, CA, USA, 1981. [Google Scholar]
- Kim, S.; Chen, J.; Cheng, T.; Gindulyte, A.; He, J.; He, S.; Li, Q.; Shoemaker, B.A.; Thiessen, P.A.; Yu, B.; et al. PubChem in 2021: New data content and improved web interfaces. Nucleic Acids Res. 2021, 49, D1388–D1395. [Google Scholar] [CrossRef] [PubMed]
- The Good Scents Company Information System. Available online: https://www.thegoodscentscompany.com/ (accessed on 1 October 2021).
- Skinner, W.A.; Tong, H.C.; Maibach, H.I.; Skidmore, D. Human skin-surface lipid fatty acids—Mosquito repellents. Experientia 1970, 26, 728–730. [Google Scholar] [CrossRef] [PubMed]
- Kaczmarek, A.; Wrońska, A.K.; Boguś, M.I.; Kazek, M.; Gliniewicz, A.; Mikulak, E.; Matławska, M. The type of blood used to feed Aedes aegypti females affects their cuticular and internal free fatty acid (FFA) profiles. PLoS ONE 2021, 16, e0251100. [Google Scholar] [CrossRef]
- Ali, A.; Cantrell, C.L.; Bernier, U.R.; Duke, S.O.; Schneider, J.C.; Agramonte, N.M.; Khan, I. Aedes aegypti (Diptera: Culicidae) biting deterrence: Structure-activity relationship of saturated and unsaturated fatty acids. J. Med. Entomol. 2012, 49, 1370–1378. [Google Scholar] [CrossRef] [PubMed]
- Maw, M.G. Capric acid as a larvicide and an oviposition stimulant for mosquitoes. Nature 1970, 227, 1154–1155. [Google Scholar] [CrossRef]
- Ikeshoji, T.; Saito, K.; Yano, A. Bacterial production of the ovipositional attractants for mosquitoes on fatty acid substrates. Appl. Entomol. Zool. 1975, 10, 239–242. [Google Scholar] [CrossRef] [Green Version]
- Owino, E.A.; Sang, R.; Sole, C.L.; Pirk, C.; Mbogo, C.; Torto, B. An improved odor bait for monitoring populations of Aedes aegypti-vectors of dengue and chikungunya viruses in Kenya. Parasites Vectors 2015, 8, 253. [Google Scholar] [CrossRef] [Green Version]
- Syed, Z. Chemical ecology and olfaction in arthropod vectors of diseases. Curr. Opin. Insect Sci. 2015, 10, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Logan, J.G.; Stanczyk, N.M.; Hassanali, A.; Kemei, J.; Santana, A.E.G.; Ribeiro, K.A.L.; Pickett, J.A.; Mordue, A.J. Arm-in-cage testing of natural human-derived mosquito repellents. Malar. J. 2010, 9, 239. [Google Scholar] [CrossRef] [Green Version]
- Cator, L. Malaria altering host attractiveness and mosquito feeding. Trends Parasitol. 2017, 33, 338–339. [Google Scholar] [CrossRef] [PubMed]
- Robinson, A.; Busula, A.O.; Voets, M.A.; Beshir, K.B.; Caulfield, J.C.; Powers, S.J.; Verhulst, N.O.; Winskill, P.; Muwanguzi, J.; Birkett, M.A.; et al. Plasmodium-associated changes in human odor attract mosquitoes. Proc. Natl. Acad. Sci. USA 2018, 115, E4209–E4218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hall, D.R.; Beevor, P.S.; Cork, A.; Nesbitt, B.F.; Vale, G.A. 1-Octen-3-ol. A potent olfactory stimulant and attractant for tsetse isolated from cattle odours. Int. J. Trop. Insect Sci. 1984, 5, 335–339. [Google Scholar] [CrossRef]
- Cork, A.; Park, K.C. Identification of electrophysiologically-active compounds for the malaria mosquito, Anopheles gambiae, in human sweat extracts. Med. Vet. Entomol. 1996, 10, 269–276. [Google Scholar] [CrossRef] [PubMed]
- Kline, D.L.; Allan, S.A.; Bernier, U.R.; Welch, C.H. Evaluation of the enantiomers of 1-octen-3-ol and 1-octyn-3-ol as attractants for mosquitoes associated with a freshwater swamp in Florida, U.S.A. Med. Vet. Entomol. 2007, 21, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Xie, L.; Yang, W.; Liu, H.; Liu, T.; Xie, Y.; Lin, F.; Zhou, G.; Zhou, X.; Wu, K.; Gu, J.; et al. Enhancing attraction of the vector mosquito Aedes albopictus by using a novel synthetic odorant blend. Parasites Vectors 2019, 12, 243. [Google Scholar] [CrossRef]
- Xu, P.; Zhu, F.; Buss, G.K.; Leal, W.S. 1-Octen-3-ol—the attractant that repels. F1000Research 2015, 4, 156. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Logan, J.G.; Birkett, M.A.; Clark, S.J.; Powers, S.; Seal, N.J.; Wadhams, L.J.; Mordue, A.J.; Pickett, J.A. Identification of human-derived volatile chemicals that interfere with attraction of Aedes aegypti mosquitoes. J. Chem. Ecol. 2008, 34, 308–322. [Google Scholar] [CrossRef]
- Suh, E.; Choe, D.-H.; Saveer, A.M.; Zwiebel, L.J. Suboptimal larval habitats modulate oviposition of the malaria vector mosquito Anopheles coluzzii. PLoS ONE 2016, 11, e0149800. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jhumur, U.S.; Dötterl, S.; Jürgens, A. Floral odors of Silene otites: Their variability and attractiveness to mosquitoes. J. Chem. Ecol. 2007, 34, 14–25. [Google Scholar] [CrossRef]
- Brantjes, N.B.M.; Leemans, J.A.A.M. Silene otites (caryophyllaceae) pollinated by nocturnal lepidoptera and mosquitoes. Acta Bot. Neerl. 1976, 25, 281–295. [Google Scholar] [CrossRef]
- Su, C.-Y.; Menuz, K.; Carlson, J.R. Olfactory perception: Receptors, cells, and circuits. Cell 2009, 139, 45–59. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fikrig, K.; Johnson, B.J.; Fish, D.; Ritchie, S.A. Assessment of synthetic floral-based attractants and sugar baits to capture male and female Aedes aegypti (Diptera: Culicidae). Parasites Vectors 2017, 10, 32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Von Oppen, S.; Masuh, H.; Licastro, S.; Zerba, E.; Gonzalez-Audino, P. A floral-derived attractant for Aedes aegypti mosquitoes. Entomol. Exp. Appl. 2015, 155, 184–192. [Google Scholar] [CrossRef]
- Morton, F.; Travis, B.; Linduska, J. Results of Screening Tests with Materials Evaluated as Insecticides, Miticides, and Repellents at the Orlando Laboratory. Available online: http://ufdc.ufl.edu/AA00025211/00001 (accessed on 5 May 2021).
- Finch, S. Volatile plant chemicals and their effect on host plant finding by the cabbage root fly (Delia brassicae). Entomol. Exp. Appl. 1978, 24, 350–359. [Google Scholar] [CrossRef]
- Dekker, T.; Ignell, R.; Ghebru, M.; Glinwood, R.; Hopkins, R. Identification of mosquito repellent odours from Ocimum forskolei. Parasites Vectors 2011, 4, 183. [Google Scholar] [CrossRef] [Green Version]
- Aisien, M.; Imasuen, A.; Wagbatsoma, V.; Ayinde, B. Preliminary evaluation of the repellent activity of some plant essential oils against Simulium damnosum s.l., the vector of human onchocerciasis. Int. J. Trop. Insect Sci. 2004, 24, 196–199. [Google Scholar] [CrossRef]
- Mwangi, E.N.; Hassanali, A.; Essuman, S.; Myandat, E.; Moreka, L.; Kimondo, M. Repellent and acaricidal properties of Ocimum suave against Rhipicephalus appendiculatus ticks. Exp. Appl. Acarol. 1995, 19, 11–18. [Google Scholar] [CrossRef] [Green Version]
- Nyasembe, V.O.; Teal, P.E.; Mukabana, W.R.; Tumlinson, J.H.; Torto, B. Behavioural response of the malaria vector Anopheles gambiae to host plant volatiles and synthetic blends. Parasites Vectors 2012, 5, 234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nyasembe, V.O.; Tchouassi, D.P.; Mbogo, C.M.; Sole, C.L.; Pirk, C.W.W.; Torto, B. Linalool oxide: Generalist plant based lure for mosquito disease vectors. Parasites Vectors 2015, 8, 581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Omolo, M.O.; Okinyo, D.; Ndiege, I.O.; Lwande, W.; Hassanali, A. Repellency of essential oils of some Kenyan plants against Anopheles gambiae. Phytochemistry 2004, 65, 2797–2802. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Portilla-Pulido, J.S.; Morales, R.M.C.; Rodríguez, M.A.B.; Duque, J.E.; Mendez-Sanchez, S. Design of a repellent against Aedes aegypti (Diptera: Culicidae) using in silico simulations with AaegOBP1 protein. J. Med. Entomol. 2019, 57, 463–476. [Google Scholar] [CrossRef] [PubMed]
- Kline, D.L.; Bernier, U.R.; Posey, K.H.; Barnard, D.R. Olfactometric evaluation of spatial repellents for Aedes aegypti. J. Med. Entomol. 2003, 40, 463–467. [Google Scholar] [CrossRef] [Green Version]
- Díaz-Santiz, E.; Rojas, J.C.; Casas-Martínez, M.; Cruz-López, L.; Malo, E.A. Rat volatiles as an attractant source for the Asian tiger mosquito, Aedes albopictus. Sci. Rep. 2020, 10, 168–175. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bruce, T.J.; Pickett, J.A. Perception of plant volatile blends by herbivorous insects–Finding the right mix. Phytochemistry 2011, 72, 1605–1611. [Google Scholar] [CrossRef] [PubMed]
- Johnson, B.J.; Ritchie, S.A. The Siren’s Song: Exploitation of female flight tones to passively capture male Aedes aegypti (Diptera: Culicidae). J. Med. Entomol. 2015, 53, 245–248. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compound | Sigma-Aldrich Product No. | Purity 1 | Family | EC No. 2 | Origin: Perceived Odour 3 |
|---|---|---|---|---|---|
| 3-Octanone | 136913 | >98% | Ketone | 203-423-0 | Fungal: fresh, herbal, lavender, mushroom |
| 2-Nonanone | 108731 | >99% | Ketone | 212-480-0 | Animal: fragrant, fruit, green, hot milk |
| Acetophenone | 42163 | NS | Ketone | 202-708-7 | Plant: almond, floral, sweet, cherry |
| 4-Propyl-benzaldehyde | 562882 | 95% | Ketone | 249-221-6 | - |
| 6-Methyl-5-hepten-2-one (sulcatone) | M48805 | >98% | Ketone | 203-816-7 | Animal: citrus, green, musty |
| Eugenol | E51791 | 99% | Ketone | 202-589-1 | Plant: clove |
| 2,6,6-Trimethyl-2-cyclohexene-1,4-dione | 14239 | ≥98% | Ketone | 214-406-2 | Pheromone: sweet, leaf, floral, tobacco |
| 3,7-Dimethyl-oct-6-en-1-yn-3-ol (Dehydrolinalool) | CDS009913 | ≤100% | Ketone | 249-482-6 | Plant: mould |
| 4′-Ethyl-acetophenone | 226750 | 97% | Ketone | 213-326-5 | Pheromone: floral, hawthorn |
| γ-Decanolactone | D804 | 98% | Ketone | 211-892-8 | Plant: fat, fruit, lactone, peach |
| Farnesene, mix of isomers | W383902 | NS | Terpene | 207-948-6 | Plant: green apple |
| Geranyl acetate | 173495 | ≥97% | Terpene | 203-341-5 | Plant: floral |
| Geraniol | 163333 | 98% | Terpene | 203-377-1 | Plant: geranium, lemon peel, passion fruit, peach, rose |
| R-(+)-limonene | 183164 | 97% | Terpene | 227-813-5 | Plant: citrus |
| Linalool oxide | 62141 | ≥97% | Terpene | 262-038-6 | Plant: floral, herbal |
| Hexyl hexanoate | W257206 | ≥97% | Ester | 228-952-4 | Plant: apple peel, peach, plum |
| Hexyl-2-methylbutanoate | W349909 | ≥95% | Ester | 233-106-2 | Plant: sweet, green apple |
| Methyl benzoate | M29908 | 99% | Ester | 202-259-7 | Plant: phenolic, almond, floral |
| Methyl 2-methyl benzoate | 259985 | 99% | Ester | 201-932-2 | Plant: floral, orange flower |
| Methyl salicylate | M6752 | ≥99% | Ester | 204-317-7 | Plant: liniments, hospital |
| Methyl cinnamate | 96410 | ≥99% | Ester | 203-093-8 | Plant: balsamic, cinnamon |
| cis-3-Hexenyl acetate | W317101 | ≥98% | Ester | 222-960-1 | Plant: banana, candy, floral, green |
| Nonanal | W278220 | 97% | Aldehyde | 204-688-5 | Animal/Plant: fat, rose, orange |
| Phenylacetaldehyde | W287407 | ≥95% | Aldehyde | 204-574-5 | Plant: honey, sweet, rose, green |
| p-Anisaldehyde | A88107 | 98% | Aldehyde | 204-602-6 | Plant: floral, aniseed |
| 2-Phenyl ethanol | 77861 | ≥99% | Alcohol | 200-456-2 | Plant: floral, honey, rose |
| 1-Octen-3-ol | O5284 | 98% | Alcohol | 222-226-0 | Animal: mushroom, earthy |
| Decanoic acid | W236403 | ≥98% | Fatty acid | 206-376-4 | Animal: goat, fat, grass, dust |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Carraretto, D.; Soresinetti, L.; Rossi, I.; Malacrida, A.R.; Gasperi, G.; Gomulski, L.M. Behavioural Responses of Male Aedes albopictus to Different Volatile Chemical Compounds. Insects 2022, 13, 290. https://doi.org/10.3390/insects13030290
Carraretto D, Soresinetti L, Rossi I, Malacrida AR, Gasperi G, Gomulski LM. Behavioural Responses of Male Aedes albopictus to Different Volatile Chemical Compounds. Insects. 2022; 13(3):290. https://doi.org/10.3390/insects13030290
Chicago/Turabian StyleCarraretto, Davide, Laura Soresinetti, Irene Rossi, Anna R. Malacrida, Giuliano Gasperi, and Ludvik M. Gomulski. 2022. "Behavioural Responses of Male Aedes albopictus to Different Volatile Chemical Compounds" Insects 13, no. 3: 290. https://doi.org/10.3390/insects13030290