
Integrative Insight into Relationships between Florivorous Thrips Haplothrips leucanthemi and H. niger (Insecta, Thysanoptera)


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
3. Results
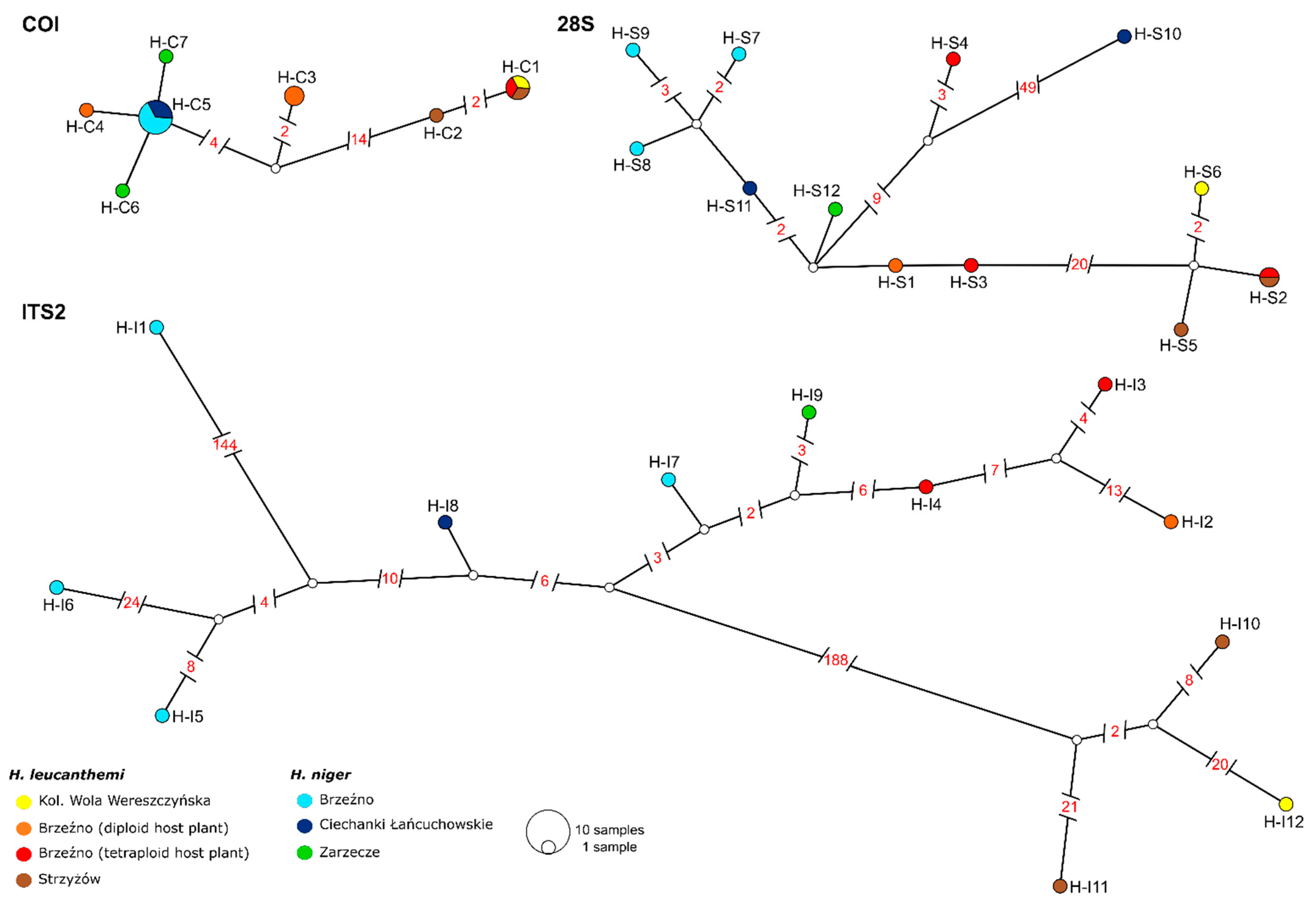
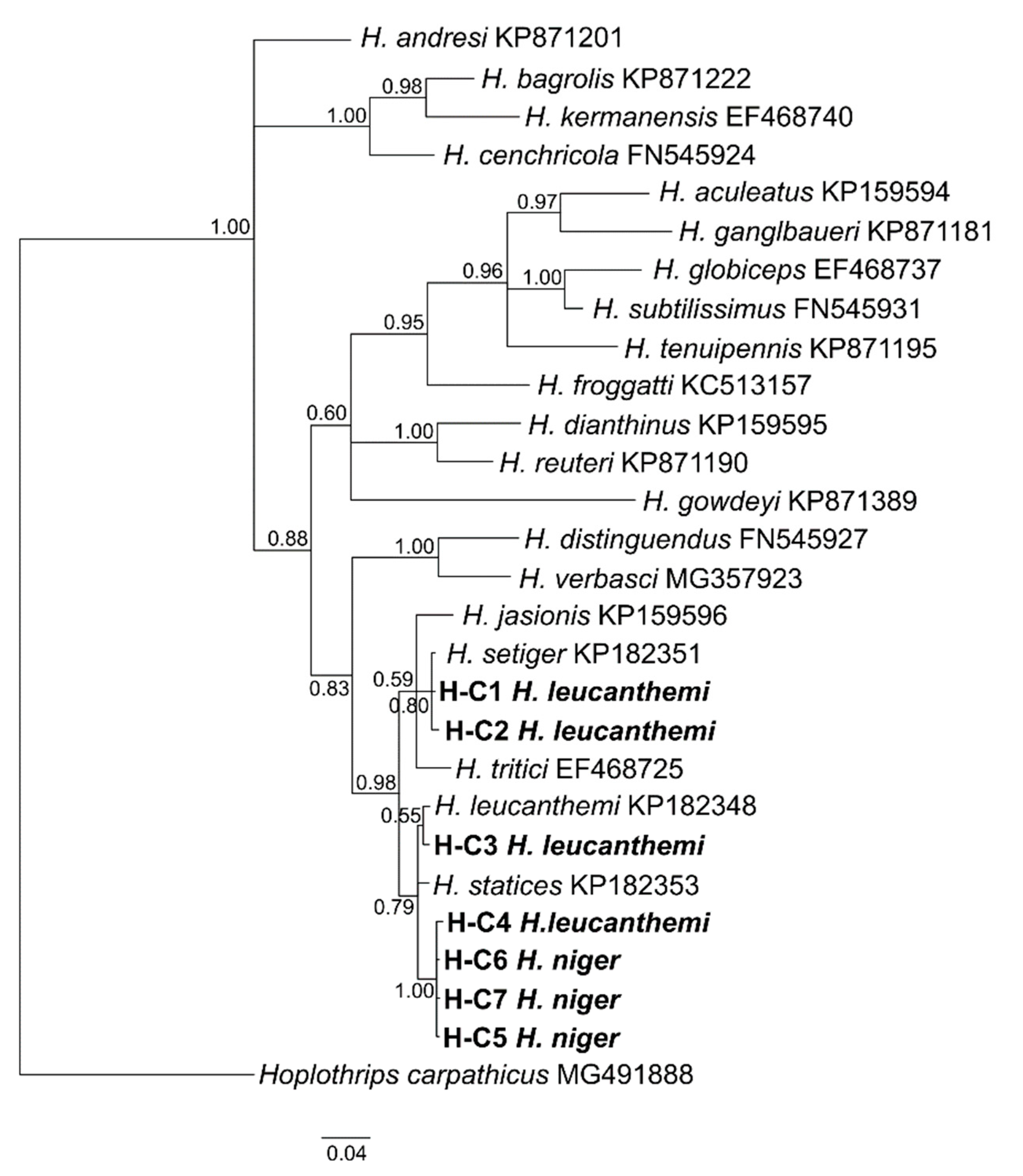
3.1. Phylogenetic Analyses
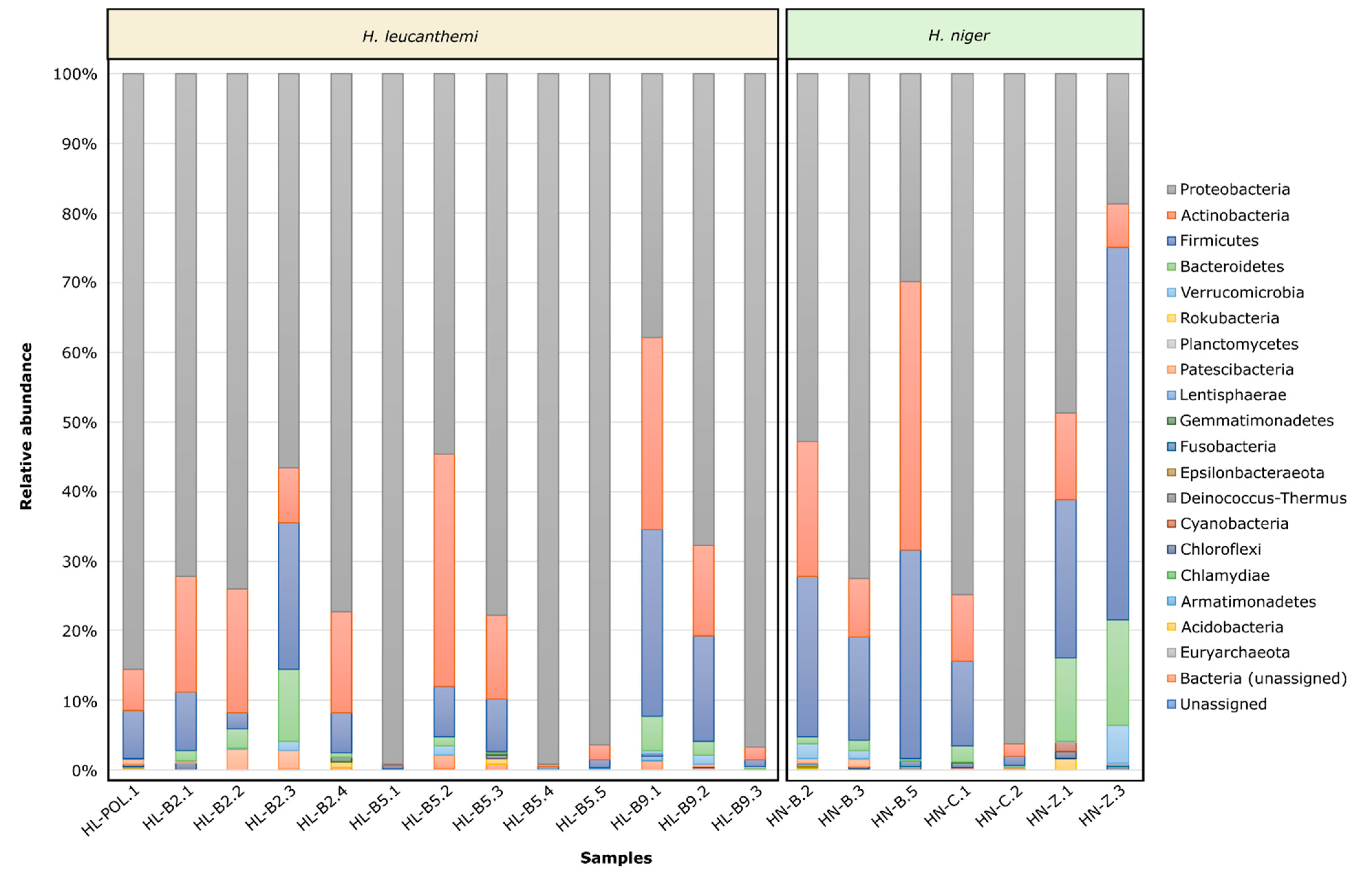
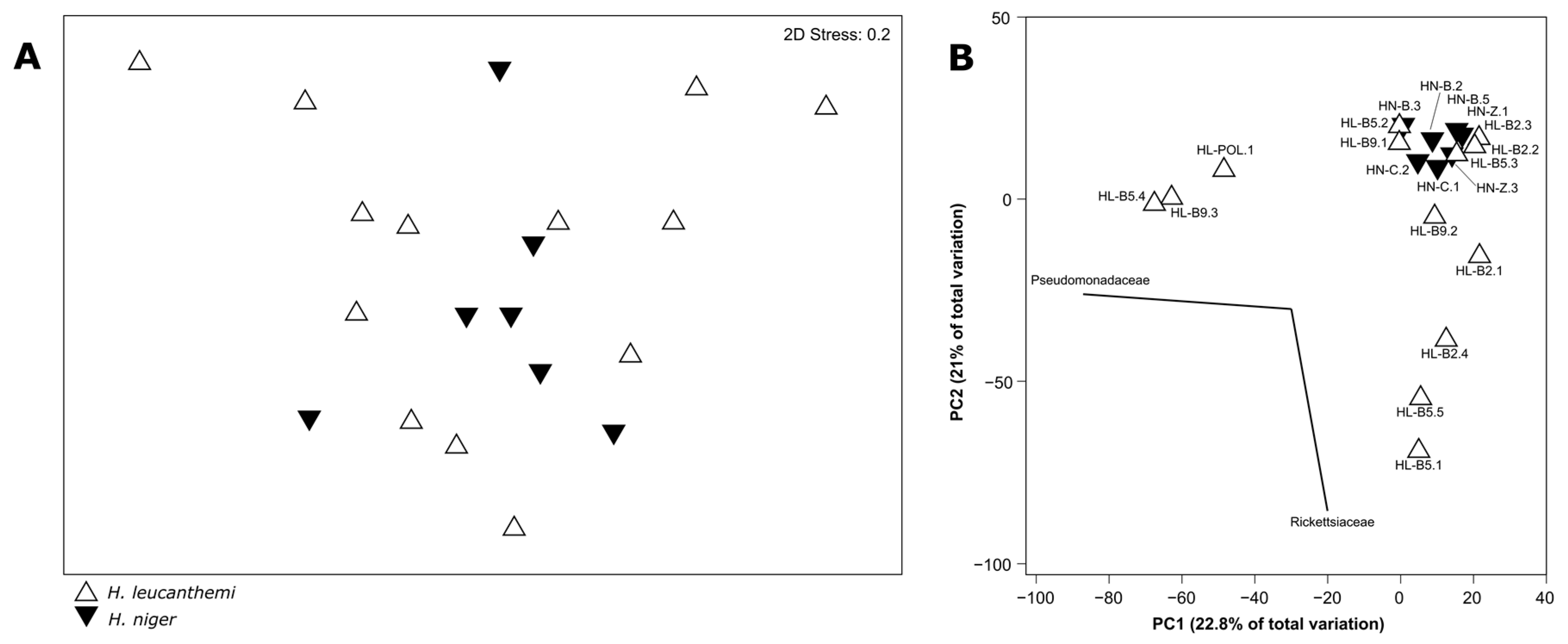
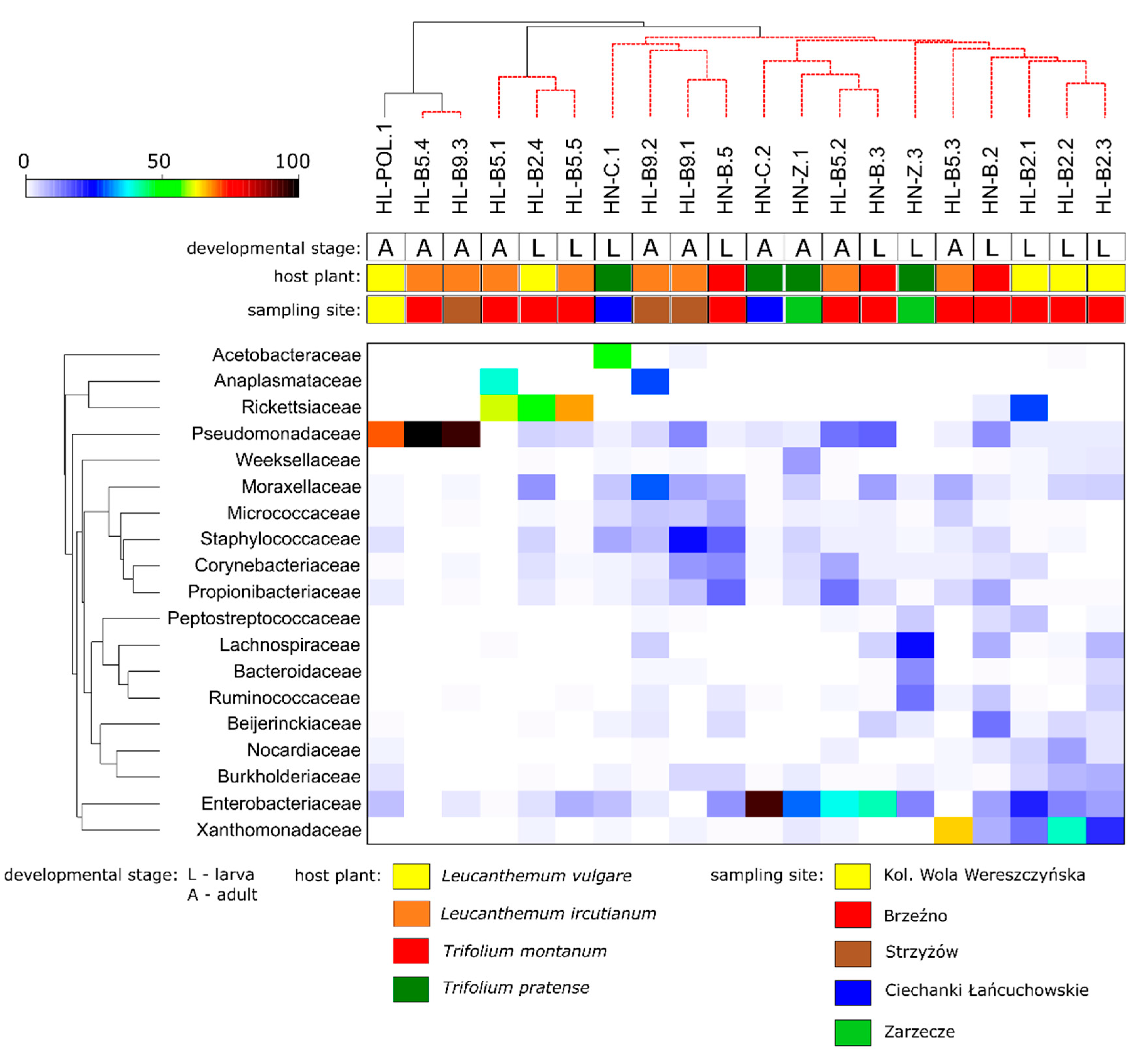
3.2. Microbiota Profiling
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A
- Sample Collection for Morphometric Study
- Morphological Comparison

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Locality/Coordinates | Host Plant | Developmental Stage | Date and Author of Collected Materials |
|---|---|---|---|---|
| Haplothrips leucanthemi | Rowokół near. Smołdzino N 54.66 E 17.21 | Chrysanthemum leucanthemum | Second larval instar | 19.07.2000 I. Zawirska |
| Urszulin N 51.40 E 23.20 | 07.06.2010 K. Stanisławek | |||
| Zawoja N 49.67 E 19.57 | 19.07.2010 A. Dembicka | |||
| Zastawie N 51.39 E 23.27 | 15.06.2010 K. Stanisławek | |||
| Białka (near Łęczna) N 51.54 E 23.01 | 23.05.2011 K. Stanisławek | |||
| Ciechanki Łańcuchowskie N 51.28 E 22.95 | 23.05.2011 K. Stanisławek | |||
| Jaszczów Kolonia N 51.21 E 22.95 | 23.05.2011 K. Stanisławek | |||
| Werbkowice N 50.75 E 23.78 | 30.05.2011 M. Kucharczyk | |||
| Udrycze (near Zamość) N 50.80 E 23.28 | 20.05.2011 K. Stanisławek | |||
| Wojtkowa (Bieszczady Mts.) N 49.57 E 22.55 | 13.08.2011 K. Stanisławek | |||
| Dziewięcierz N 50.31 E 23.42 | 25.06.2011 K. Stanisławek | |||
| Mała Rawka (Bieszczady Mts.) N 49.11 E 22.57 | 04.08.2011 H. Kucharczyk | |||
| Haplothrips niger | Ursynów (Warszawa) N 52.15 E 21.03 | Trifolium pratense | Second larval instar | 22.10.1984 I. Zawirska |
| Warszawa, Huta N 52.30 E 20.91 | 27.06.1995 I. Zawirska | |||
| Jakubowice Konińskie N 51.31 E 22.55 | 13.06.2010 K. Stanisławek | |||
| Lublin N 51.25 E 22.57 | 25.05.2011 20.06.2011 22.06.2011 K. Stanisławek | |||
| Białka (near Łęczna) N 51.54 E 23.01 | 23.05.2011 K. Stanisławek | |||
| Rudnik (near Lublin) N 51.28 E 22.60 | 20.06.2011 K. Stanisławek | |||
| Ciechanki Łańcuchowskie N 51.28 E 22.95 | 23.05.2011 K. Stanisławek | |||
| Polanka Horyniecka (Roztocze) N 51.25 E 22.57 | 25.06.2011 K. Stanisławek | |||
| Bandrów Narodowy (Bieszczady Mts.) N 49.39 E 22.70 | 13.08.2011 K. Stanisławek | |||
| Muczne (Bieszczady Mts.) N 49.13 E 22.74 | Trifolium repens | Second larval instar | 14.08.2011 K. Stanisławek |
| Character | H. leucanthemi | H. niger |
|---|---|---|
| Body length | 1200–1780 (1422) | 1317–1933 (1636) |
| Head—length/width | 190–219 (206.5)/110–153 (133.0) | 185–211 (197.0)/109–134 (120.5) |
| Head—length/width proportion | 1.5–1.9 (1.65) | 1.4–1.8 (1.63) |
| Head—length of dorsal setae D1, D2 | D1 37.5–51.0 (43.2) D2 14.0–22.5 (17.1) | D1 36.0–51.0 (44.2) D2 12.0–20.0 (16.9) |
| Length of antennae | 206–250 (223.5) | 214–272 (248.0) |
| Prothorax—length of dorsal setae D1, D4, D5, D6 | D1 22.0–32.0 (27.2) D4 28.0–43.0 (35.2) D5 53.0–69.0 (60.5) D6 60.5–77.0 (68.7) | D1 24.0–38.0 (31.3) D4 30.0–44.0 (37.5) D5 51.0–74.0 (65.3) D6 61.5–85.0 (73.3) |
| Mesothrax—length of dorsal setae D1, D2, D3, D5, D6 | D1 26.0–38.0 (31.8) D2 16.0–22.5 (18.4) D3 19.0–30.0 (22.8) D5 55.0–77.0 (64.0) D6 35.0–50.0 (42.1) | D1 29.0–41.0 (35.6) D2 17.0–26.0 (22.5) D3 18.0–29.0 (25.7) D5 62.0–81.0 (72.5) D6 36.0–49.0 (42.7) |
| Metathorax—length of dorsal setae D1, D2, D3, D5, D6 | D1 27.0–40.5 (33.6) D2 16.0–24.0 (19.8) D3 17.5–37.5 (27.1) D5 59.0–75.0 (66.6) D6 32.5–52.0 (41.8) | D1 31.0–44.0 (38.1) D2 18.0–25.0 (21.7) D3 19.0–35.0 (27.3) D5 64.0–80.0 (73.2) D6 38.0–50.0 (44.7) |
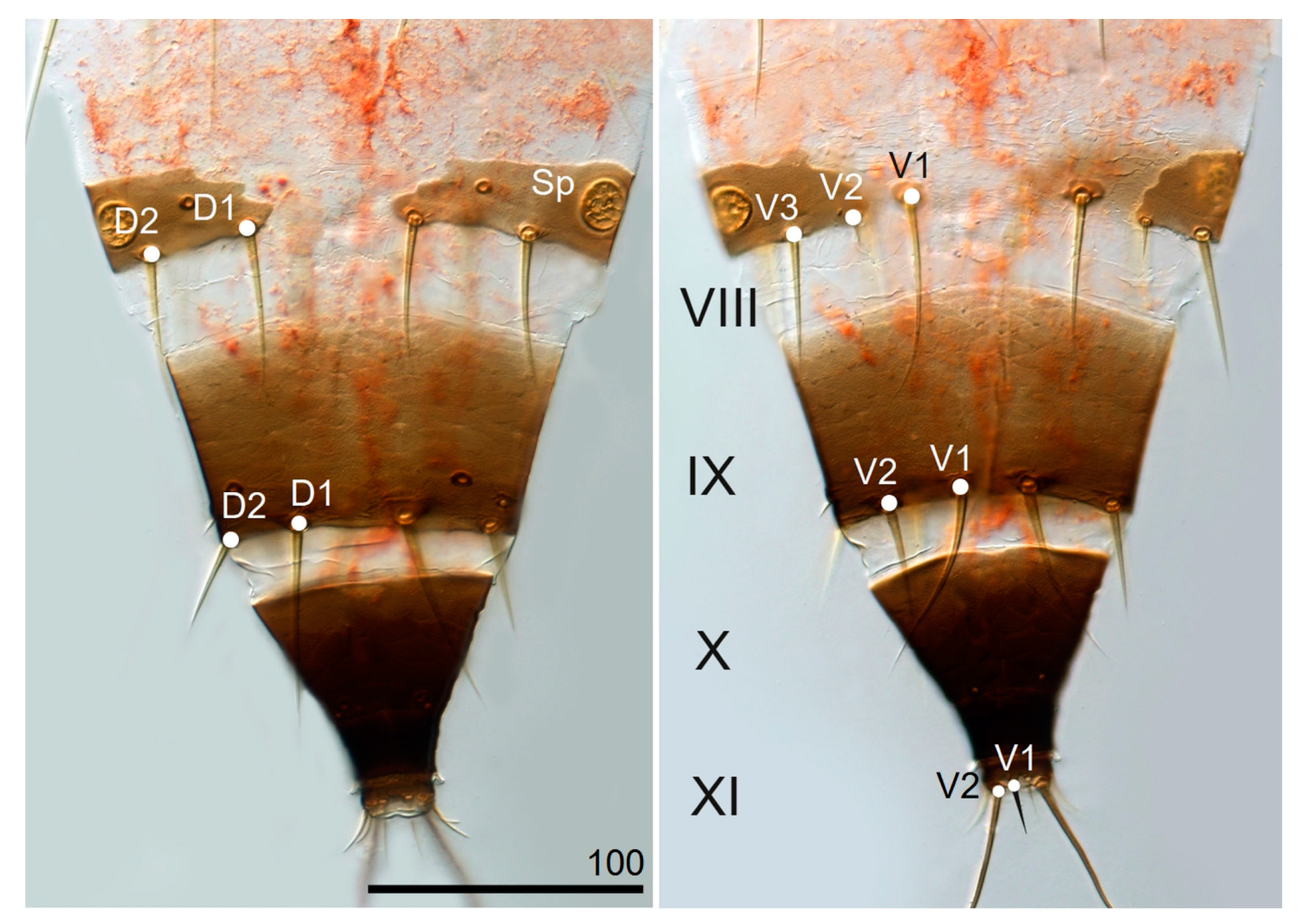
| Abdominal tergite VIII—length of dorsal setae D1, D2 | D1 45.0–62.5 (53.1) D2 55.0–70.0 (61.0) | D1—52.0–72.0 (62.5) D2—52.0–72.0 (63.1) |
| Abdominal tergite IX—length of dorsal setae D1, D2 | D1 72.5–90.0 (81.1) D2 39.0–53.5 (45.2) | D1 72.0–91.0 (82.5) D2 32.0–51.0 (42.7) |
| Abdominal sternite VIII—length of ventral setae V1, V2, V3 | V1 57.0–88.0 (72.0) V2 12.0–21.0 (16.1) V3 46.0–68.5 (56.3) | V1 61.0–90.0 (75.1) V2 11.0–20.0 (16.0) V3 52.0–67.0 (59.9) |
| Abdominal sternite IX—length of ventral setae V1, V2 | V1 73.0–102.0 (86.1) V2 42.5–60.0 (50.1) | V1 70.0–99.0 (85.5) V2 32.0–56.0 (46.3) |
| Abdominal segment X—length | 69.0–89.0 (78.6) | 67.0–84.0 (77.1) |
| Abdominal segment X—proportion of anterior and posteriori rand width | 2.1–3.3 (2.8) | 2.1–2.6 (2.4) |
| Abdominal segment XI, length of setae D1, V1, V2 | D1 17.5–28.0 (22.4) V1 10.5–20.0 (14.2) V2 122.0–160.0 (138.0) | D1 18.0–28.0 (23.8) V1 9.0–18.0 (14.1) V2 118.0–170.0 (147.0) |
| Meso- and metanotum—sclerotization at basis of setae | weak or absent, may be stronger around setae D3, D5, D6 | weak or absent |
| Abdomen—sclerotization at basis of dorsal setae | weak or absent, may be stronger around setae on segments VI-VIII | weak or absent |
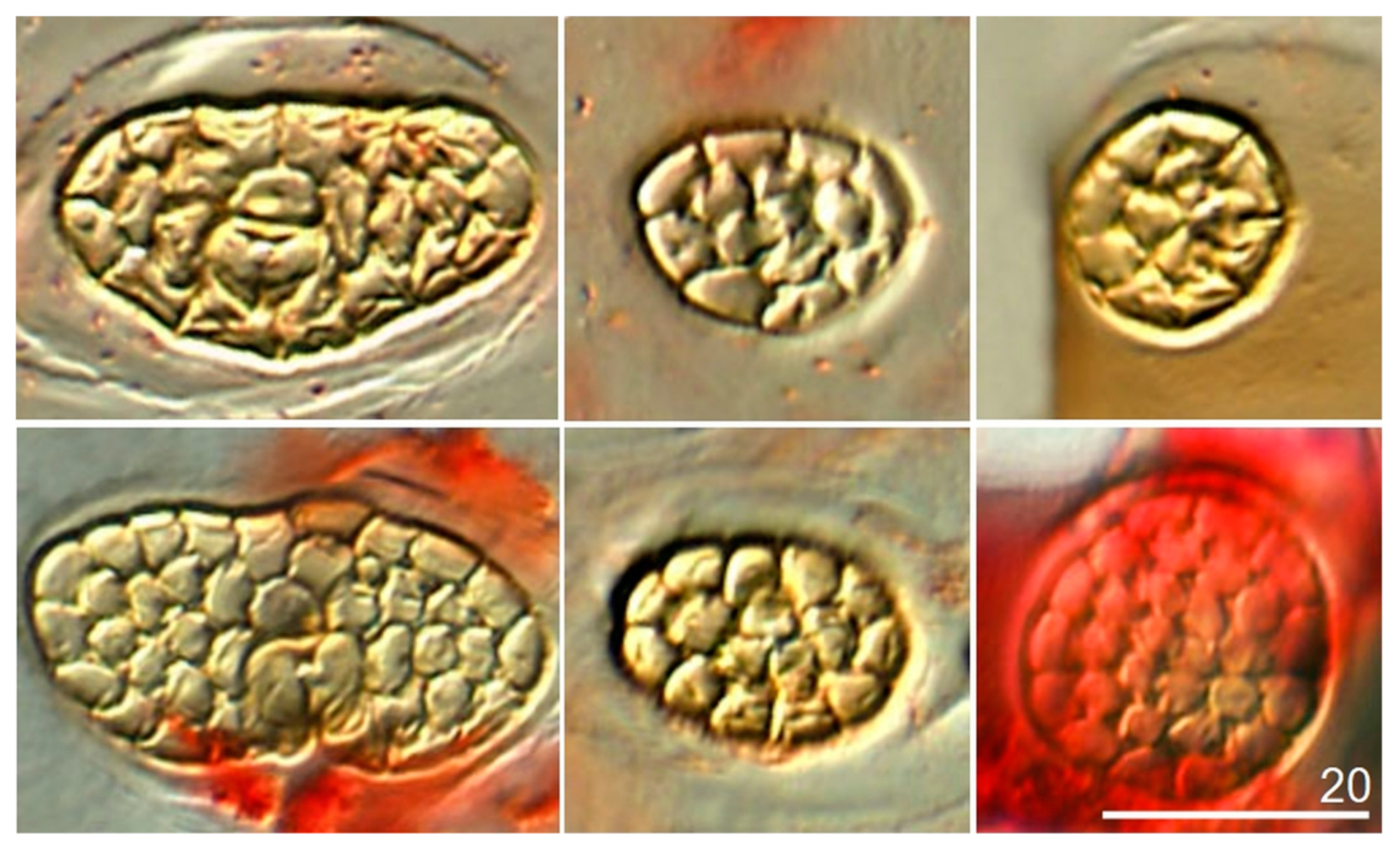
| Spiracle on mesonotum—length/width | 20–28 (23.8)/37–47 (42.1) | 19–21 (20.2)/35–38 (36.3) |
| Spiracle on mesonotum—number of facets (cells). Mean number of cells | 27–46 (35) >30 | 20–27 (23) <30 |
| Spiracles on abdominal tergite II—length/width | 14–26 (18)/17–27 (22) | 14–16 (15)/16–21 (19) |
| Spiracles on abdominal tergite II—number of facets (cells) | 7–22 (15) | 6–17 (11) |
| Spiracles on abdominal tergite VIII—diameter | 17–27 (22) | 16–23 (18) |
| Spiracles on abdominal tergite VIII—number of facets (cells) Mean number of cells | 20–30 (24) >15 | 10–15 (12) <15 |



References
- Mound, L.A.; Hastenpflug-Vesmanis, A. All genera of the world: Order Thysanoptera (Animalia: Arthropoda: Insecta). Megataxa 2021, 6, 2–69. [Google Scholar] [CrossRef]
- Minaei, K.; Mound, L.A. Character-state evaluation when discriminating Thysanoptera taxa (Insecta). Zootaxa 2021, 5061, 377–382. [Google Scholar] [CrossRef] [PubMed]
- Mound, L.A. Fighting, flight and fecundity: Behavioural determinants of thysanoptera structural diversity. In Insects and Phenotypic Plasticity; Ananthakrishnan, T.N., Whitman, D., Eds.; Science Publishers Inc.: Enfield, NH, USA, 2005; pp. 81–105. [Google Scholar]
- Mound, L.A.; Matsunaga, J.N. The species of Haplothrips (Thysanoptera, Phlaeothripinae) and related genera recorded from the Hawaiian Islands. Zookeys 2017, 2017, 79–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Minaei, K.; Mound, L. The Thysanoptera Haplothripini (Insecta: Phlaeothripidae) of Iran. J. Nat. Hist. 2008, 42, 2617–2658. [Google Scholar] [CrossRef]
- Minaei, K.; Mound, L. New synonymy in the wheat thrips, Haplothrips tritici (Thysanoptera: Phlaeothripidae). Zootaxa 2014, 3802, 596–599. [Google Scholar] [CrossRef] [Green Version]
- Timm, A.E.; Stiller, M.; Frey, J.E. A molecular identification key for economically important thrips species (Thysanoptera: Thripidae) in southern Africa Article. Afr. Entomol. 2008, 16, 68–75. [Google Scholar] [CrossRef]
- Tyagi, K.; Kumar, V.; Singha, D.; Chandra, K.; Laskar, B.A.; Kundu, S.; Chakraborty, R.; Chatterjee, S. DNA barcoding studies on thrips in India: Cryptic species and species complexes. Sci. Rep. 2017, 7, 4898. [Google Scholar] [CrossRef] [Green Version]
- Mound, L.A.; Minaei, K. Australian thrips of the Haplothrips lineage (Insecta: Thysanoptera). J. Nat. Hist. 2007, 41, 2919–2978. [Google Scholar] [CrossRef]
- Loan, C.; Holdaway, F.G. Biology of the red clover thrips, Haplothrips niger (Osborn) (Thysanoptera: Phloeothripidae). Can. Entomol. 1955, 87, 210–219. [Google Scholar] [CrossRef]
- Bournier, A. Les Thrips: Biologie, Importance, Agronomique; Institut National de la Recherche Agronomique: Paris, France, 1983; ISBN 2853404641. [Google Scholar]
- Mound, L.A.; Walker, A.K. Tubulifera (Insecta: Thysanoptera). Fauna N. Z. 1986, 10, 53–54. [Google Scholar] [CrossRef]
- Priesner, H. Ordnung Thysanoptera (Fransenflügler Thripse), 2nd ed.; Bestimmungsbücher zur Bodenfauna Europas: Berlin, Germany, 1964. [Google Scholar]
- Schliephake, G.; Klimt, K. Thysanoptera, Franseflügler; Fischer, G., Ed.; Die Tierwelt Deutschlands: Bonn, Germany, 1979; pp. 5–477. [Google Scholar]
- Moritz, G. Thripse: Fransenflügler, Thysanoptera. Die Neue Brehm-Bücherei Bd. 663; Westarp Wissenschaften: Hohenwarsleben, Germany, 2006. [Google Scholar]
- Kucharczyk, H.; Stanisławek, K. Haplothrips second instar larvae (Thysanoptera: Phlaeothripidae); character states and key to Central European species. Zootaxa 2020, 4845, 375–392. [Google Scholar] [CrossRef] [PubMed]
- Folmer, O.; Black, M.; Hoeh, W.; Lutz, R.; Vrijenhoek, R. DNA primers for amplification of mitochondrial cytochrome c oxidase subunit I from diverse metazoan invertebrates. Mol. Mar. Biol. Biotechnol. 1994, 3, 294–299. [Google Scholar] [PubMed]
- Whiting, M.F. Mecoptera is paraphyletic: Multiple genes and phylogeny of Mecoptera and Siphonaptera. Zool. Scr. 2002, 31, 93–104. [Google Scholar] [CrossRef] [Green Version]
- Buckman, R.S.; Mound, L.A.; Whiting, M.F. Phylogeny of thrips (Insecta: Thysanoptera) based on five molecular loci. Syst. Entomol. 2013, 38, 123–133. [Google Scholar] [CrossRef]
- Kumar, V.; Dickey, A.M.; Seal, D.R.; Shatters, R.G.; Osborne, L.S.; McKenzie, C.L. Unexpected high intragenomic variation in two of three major pest thrips species does not affect ribosomal internal transcribed spacer 2 (ITS2) utility for thrips identification. Int. J. Mol. Sci. 2017, 18, 2100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Klindworth, A.; Pruesse, E.; Schweer, T.; Peplies, J.; Quast, C.; Horn, M.; Glöckner, F.O. Evaluation of general 16S ribosomal RNA gene PCR primers for classical and next-generation sequencing-based diversity studies. Nucleic Acids Res. 2013, 41, e1. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Hall, T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Soding, J.; et al. Fast, scalable generation of high-quality protein multiple sequence alignments using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Rice, P.; Longden, I.; Bleasby, A. EMBOSS: The European molecular biology open software suite. Trends Genet. 2000, 16, 276–277. [Google Scholar] [CrossRef]
- Goujon, M.; McWilliam, H.; Li, W.; Valentin, F.; Squizzato, S.; Paern, J.; Lopez, R. A new bioinformatics analysis tools framework at EMBL-EBI. Nucleic Acids Res. 2010, 38, W695–W699. [Google Scholar] [CrossRef] [PubMed]
- Librado, P.; Rozas, J. DnaSP v5: A software for comprehensive analysis of DNA polymorphism data. Bioinformatics 2009, 25, 1451–1452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. jModelTest 2: More models, new heuristics and parallel computing. Nat. Methods 2012, 9, 772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huelsenbeck, J.P.; Ronquist, F. MRBAYES: Bayesian inference of phylogenetic trees. Bioinformatics 2001, 17, 754–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Templeton, A.R.; Crandall, K.A.; Sing, C.F. A cladistic analysis of phenotypic associations with haplotypes inferred from restriction endonuclease mapping and DNA sequence data. III. Cladogram estimation. Genetics 1992, 132, 619–633. [Google Scholar] [CrossRef]
- Leigh, J.W.; Bryant, D. PopART: Full-feature software for haplotype network construction. Methods Ecol. Evol. 2015, 6, 1110–1116. [Google Scholar] [CrossRef]
- Bolyen, E.; Rideout, J.R.; Dillon, M.R.; Bokulich, N.A.; Abnet, C.C.; Al-Ghalith, G.A.; Alexander, H.; Alm, E.J.; Arumugam, M.; Asnicar, F.; et al. Reproducible, interactive, scalable and extensible microbiome data science using QIIME 2. Nat. Biotechnol. 2019, 37, 852–857. [Google Scholar] [CrossRef]
- Callahan, B.J.; McMurdie, P.J.; Rosen, M.J.; Han, A.W.; Johnson, A.J.A.; Holmes, S.P. DADA2: High resolution sample inference from Illumina amplicon data. Nat. Methods 2016, 13, 581–583. [Google Scholar] [CrossRef] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glöckner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Accids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Clarke, K.R.; Gorley, R. PRIMER Version 7: User Manual/Tutorial; Prime Data Centre (Plymouth) Limited: Plymouth, UK, 2015. [Google Scholar]
- Hammer, Ø.; Harper, D.A.T.; Ryan, P.D. PAST: Paleontological statistics software package for education and data analysis Palaeontol. Electron. 2001, 4, 1–9. [Google Scholar]
- Clarke, K.R.; Somerfield, P.J.; Gorley, R.N. Testing of null hypotheses in exploratory community analyses: Similarity profiles and biota-environment linkage. J. Exp. Mar. Bio. Ecol. 2008, 366, 56–69. [Google Scholar] [CrossRef]
- Murai, T.; Toda, S. Variation on Thrips tabaci in colour and size. In Proceedings of the 7th International Symposium on Thysanoptera, Reggio Calabria, Italy, 1–8 July 2001; pp. 377–378. [Google Scholar]
- Mound, L.A.; Wheeler, G.; Williams, D. Zootaxa, resolving cryptic species with morphology and DNA: Thrips as a potential biocontrol agent of Brazilian peppetree, with a new species and overview of Pseudophilothrips (Thysanoptera). Zootaxa 2010, 2432, 59–68. [Google Scholar] [CrossRef]
- Brunner, P.C.; Chatzivassiliou, E.K.; Katis, N.I.; Frey, J.E. Host-associated genetic differentiation in Thrips tabaci (Insecta; Thysanoptera), as determined from mtDNA sequence data. Heredity 2004, 93, 364–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iftikhar, R.; Ashfaq, M.; Rasool, A.; Hebert, P.D.N. DNA barcode analysis of Thrips (Thysanoptera) diversity in Pakistan Reveals cryptic species complexes. PLoS ONE 2016, 11, e0146014. [Google Scholar] [CrossRef]
- Kadirvel, P.; Srinivasan, R.; Hsu, Y.C.; Su, F.C.; Del La Peña, R. Application of cytochrome oxidase i sequences for phylogenetic analysis and identification of thrips species occurring on vegetable crops. J. Econ. Entomol. 2013, 106, 408–418. [Google Scholar] [CrossRef]
- Chakraborty, R.; Singha, D.; Kumar, V.; Pakrashi, A.; Kundu, S.; Chandra, K.; Patnaik, S.; Tyagi, K. DNA barcoding of selected Scirtothrips species (Thysanoptera) from India. Mitochondrial DNA Part B Resour. 2019, 4, 2710–2714. [Google Scholar] [CrossRef] [Green Version]
- Cubillos-Salamanca, Y.P.; Rodríguez-Maciel, J.C.; Pineda-Guillermo, S.; Silva-Rojas, H.V.; Berzosa, J.; Tejeda-Reyes, M.A.; Rebollar-Alviter, Á. Identification of Thrips species and resistance of Frankliniella occidentalis (Thysanoptera: Thripidae) to malathion, spinosad, and bifenthrin in blackberry crops. Fla. Entomol. 2019, 102, 738–746. [Google Scholar] [CrossRef] [Green Version]
- Karimi, J.; Hassani-Kakhki, M.; Modarres Awal, M. Identifying thrips (Insecta: Thysanoptera) using DNA barcodes. J. Cell Mol. Res. 2011, 2, 35–41. [Google Scholar] [CrossRef]
- Brunner, P.C.; Fleming, C.; Frey, J.E. A molecular identification key for economically important thrips species (Thysanoptera: Thripidae) using direct sequencing and a PCR-RFLP-based approach. Agric. For. Entomol. 2002, 4, 127–136. [Google Scholar] [CrossRef]
- Asokan, R.; Krishna Kumar, N.K.; Kumar, V.; Ranganath, H.R. Molecular differences in the mitochondrial cytochrome oxidase I (mtCOI) gene and development of a species-specific marker for onion thrips, Thrips tabaci Lindeman, and melon thrips, T. palmi Karny (Thysanoptera: Thripidae), vectors of tospoviruses bunyav. Bull. Entomol. Res. 2007, 97, 461–470. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, K.; Hasegawa, E. Discrimination of reproductive forms of Thrips tabaci (Thysanoptera: Thripidae) by PCR with sequence specific primers. J. Econ. Entomol. 2012, 105, 555–559. [Google Scholar] [CrossRef] [PubMed]
- Marullo, R.; Mercati, F.; Vono, G. DNA barcoding: A reliable method for the identification of thrips species (Thysanoptera, Thripidae) collected on sticky traps in onion fields. Insects 2020, 11, 489. [Google Scholar] [CrossRef] [PubMed]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; DeWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. B Biol. Sci. 2003, 270, 313–321. [Google Scholar] [CrossRef] [Green Version]
- Hebert, P.D.N.; Ratnasingham, S.; DeWaard, J.R. Barcoding animal life: Cytochrome c oxidase subunit 1 divergences among closely related species. Proc. R. Soc. B Biol. Sci. 2003, 270, 96–99. [Google Scholar] [CrossRef] [Green Version]
- Ghosh, A.; Jangra, S.; Dietzgen, R.G.; Yeh, W. Bin frontiers approaches to the diagnosis of thrips (Thysanoptera): How effective are the molecular and electronic detection platforms? Insects 2021, 12, 920. [Google Scholar] [CrossRef]
- Kim, J.M.; Choi, M.-Y.; Kim, J.-W.; Lee, S.A.; Ahn, J.-H.; Song, J.; Kim, S.-H.; Weon, H.-Y. Effects of diet type, developmental stage, and gut compartment in the gut bacterial communities of two Cerambycidae species (Coleoptera). J. Microbiol. 2017, 55, 21–30. [Google Scholar] [CrossRef]
- Colman, D.R.; Toolson, E.C.; Takacs-Vesbach, C.D. Do diet and taxonomy influence insect gut bacterial communities? Mol. Ecol. 2012, 21, 5124–5137. [Google Scholar] [CrossRef]
- Yun, J.-H.; Roh, S.W.; Whon, T.W.; Jung, M.-J.; Kim, M.-S.; Park, D.-S.; Yoon, C.; Nam, Y.-D.; Kim, Y.-J.; Choi, J.-H.; et al. Insect gut bacterial diversity determined by environmental habitat, diet, developmental stage, and phylogeny of host. Appl. Environ. Microbiol. 2014, 80, 5254–5264. [Google Scholar] [CrossRef] [Green Version]
- Kaczmarczyk, A.; Kucharczyk, H.; Kucharczyk, M.; Kapusta, P.; Sell, J.; Zielińska, S. First insight into microbiome profile of fungivorous thrips Hoplothrips carpathicus (Insecta: Thysanoptera) at different developmental stages: Molecular evidence of Wolbachia endosymbiosis. Sci. Rep. 2018, 8, 14376. [Google Scholar] [CrossRef]
- Dickey, A.M.; Trease, A.J.; Jara-Cavieres, A.; Kumar, V.; Christenson, M.K.; Potluri, L.-P.; Morgan, J.K.; Shatters, R.G.; Mckenzie, C.L.; Davis, P.H.; et al. Estimating bacterial diversity in Scirtothrips dorsalis (Thysanoptera: Thripidae) via Next Generation Sequencing. Fla. Entomol. 2014, 97, 362–366. [Google Scholar] [CrossRef] [PubMed]
- Gawande, S.J.; Anandhan, S.; Ingle, A.; Roylawar, P.; Khandagale, K.; Gawai, T.; Jacobson, A.; Asokan, R.; Singh, M. Microbiome profiling of the onion thrips, Thrips tabaci Lindeman (Thysanoptera: Thripidae). PLoS ONE 2019, 14, e0223281. [Google Scholar] [CrossRef] [PubMed]
- Zhao, X.; Zhang, X.; Chen, Z.; Wang, Z.; Lu, Y.; Cheng, D. The divergence in bacterial components associated with Bactrocera dorsalis across developmental stages. Front. Microbiol. 2018, 9, 114. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Majumder, R.; Sutcliffe, B.; Taylor, P.W.; Chapman, T.A. Microbiome of the queensland fruit fly through metamorphosis. Microorganisms 2020, 8, 795. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Xiang, X.; Wan, X. Divergence in gut bacterial community among life stages of the rainbow stag beetle Phalacrognathus muelleri (Coleptera: Lucanidae). Insects 2020, 11, 719. [Google Scholar] [CrossRef]
- Kaczmarczyk-Ziemba, A.; Wagner, G.K.; Grzywnowicz, K.; Kucharczyk, M.; Zielińska, S. The microbiome profiling of fungivorous black tinder fungus beetle Bolitophagus reticulatus reveals the insight into bacterial communities associated with larvae and adults. PeerJ 2019, 7, e6852. [Google Scholar] [CrossRef]
- Ma, W.J.; Schwander, T. Patterns and mechanisms in instances of endosymbiont-induced parthenogenesis. J. Evol. Biol. 2017, 30, 868–888. [Google Scholar] [CrossRef] [Green Version]
- Kumm, S.; Moritz, G. First detection of Wolbachia in arrhenotokous populations of Thrips species (Thysanoptera: Thripidae and Phlaeothripidae) and its role in reproduction. Environ. Entomol. 2008, 37, 1422–1428. [Google Scholar] [CrossRef]
- Arakaki, N.; Miyoshi, T.; Noda, H. Wolbachia-mediated parthenogenesis in the predatory thrips Franklinothrips vespiformis (Thysanoptera: Insecta). Proc. R. Soc. B. Biol. Sci. 2001, 268, 1011–1016. [Google Scholar] [CrossRef] [Green Version]
- Mound, L.A.; Kibby, G. Thysanoptera: An Identification Guide, 2nd ed.; CAB International: Wallingford, UK, 1998. [Google Scholar]





| Species | Sample Code | Locality/Coordinates | Host Plant | Collected Developmental Stages |
|---|---|---|---|---|
| Haplothrips leucanthemi | HL-POL | Kol. Wola Wereszczyńska 51.45° N, 23.15° E | Leucanthemum vulgare (diploid) | HL-POL.1—adult |
| HL-B2 | Brzeźno 51.17° N, 23.62° E | HL-B2.1—larva | ||
| HL-B2.2—larva | ||||
| HL-B2.3—larva | ||||
| HL-B2.4—larva | ||||
| HL-B5 | Brzeźno 51.17° N, 23.62° E | Leucanthemum ircutianum (tetraploid) | HL-B5.1—adult | |
| HL-B5.2—adult | ||||
| HL-B5.3—adult | ||||
| HL-B5.4—adult | ||||
| HL-B5.5—adult | ||||
| HL-B9 | Strzyżów 50.85° N, 24.02° E | HL-B9.1—adult | ||
| HL-B9.2—adult | ||||
| HL-B9.3—adult | ||||
| Haplothrips niger | HN-B | Brzeźno 51.17° N 23.62° E | Trifolium montanum | HN-B.2—larva |
| HN-B.2—larva | ||||
| HN-B.5—larva | ||||
| HN-C | Ciechanki Łańcuchowskie 51.28° N, 22.95° E | Trifolium pratense | HN-C.1—larva | |
| HN-C.2—adult | ||||
| HN-Z | Zarzecze near San River 50.53° N, 22.20° E | HN-Z.1—adult | ||
| HN-Z.3—larva |
| Sample | COI | 28S | ITS2 |
|---|---|---|---|
| H. leucanthemi | |||
| Kol. Wola Wereszczyńska (HL-POL) | H-C1 (1) | H-S6 (1) | H-I12 (1) |
| Brzeźno—diploid host plant (HL-B2) | H-C3 (2), H-C4 (1) | H-S1 (1) | H-I2 (1) |
| Brzeźno—tetraploid host plant (HL-B5) | H-C1 (1) | H-S2 (1), H-S3 (1), H-S4 (1) | H-I3 (1), H-I4 (1) |
| Strzyżów (HL-B9) | H-C1 (1), H-C2 (1) | H-S2 (1), H-S5 (1) | H-I10 (1), H-I11 (1) |
| H. niger | |||
| Brzeźno (HN-B) | H-C5 (4) | H-S7 (1), H-S8 (1), H-S9 (1) | H-I1 (1), H-I5 (1), H-I6 (1) |
| Ciechanki Łańcuchowskie (HN-C) | H-C5 (2) | H-S10 (1), H-S11 (1) | H-I7 (1), H-I8 (1) |
| Zarzecze near San River (HN-Z) | H-C6 (1), H-C7 (1) | H-S12 (1) | H-I9 (1) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaczmarczyk-Ziemba, A.; Kucharczyk, H.; Kucharczyk, M.; Kucharska, K. Integrative Insight into Relationships between Florivorous Thrips Haplothrips leucanthemi and H. niger (Insecta, Thysanoptera). Insects 2022, 13, 279. https://doi.org/10.3390/insects13030279
Kaczmarczyk-Ziemba A, Kucharczyk H, Kucharczyk M, Kucharska K. Integrative Insight into Relationships between Florivorous Thrips Haplothrips leucanthemi and H. niger (Insecta, Thysanoptera). Insects. 2022; 13(3):279. https://doi.org/10.3390/insects13030279
Chicago/Turabian StyleKaczmarczyk-Ziemba, Agnieszka, Halina Kucharczyk, Marek Kucharczyk, and Kinga Kucharska. 2022. "Integrative Insight into Relationships between Florivorous Thrips Haplothrips leucanthemi and H. niger (Insecta, Thysanoptera)" Insects 13, no. 3: 279. https://doi.org/10.3390/insects13030279