
Insecticidal Activity of Plectranthus amboinicus Essential Oil against the Stable Fly Stomoxys calcitrans (Diptera: Muscidae) and the Horse Fly Tabanus megalops (Diptera: Tabanidae)


 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Ethical Statement
2.2. Insects
2.3. Essential Oil Extraction and Quantification
2.4. Contact Toxicity Test
2.5. Fumigant Toxicity Test
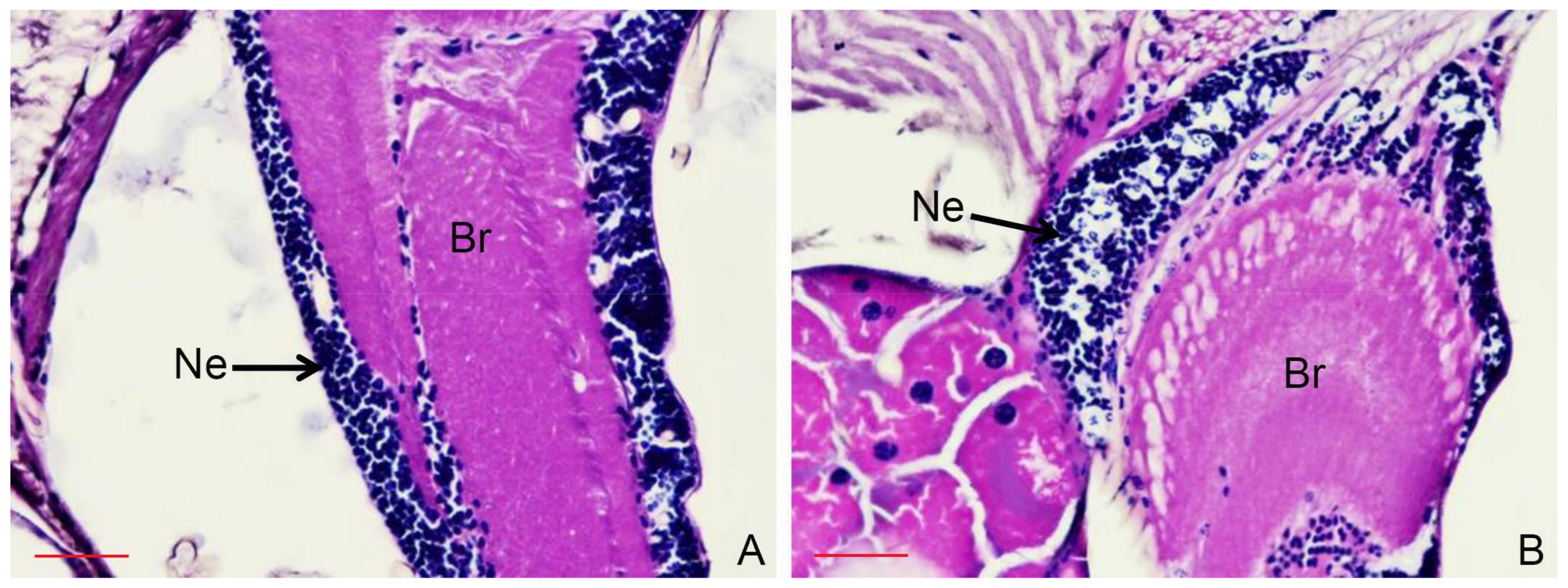
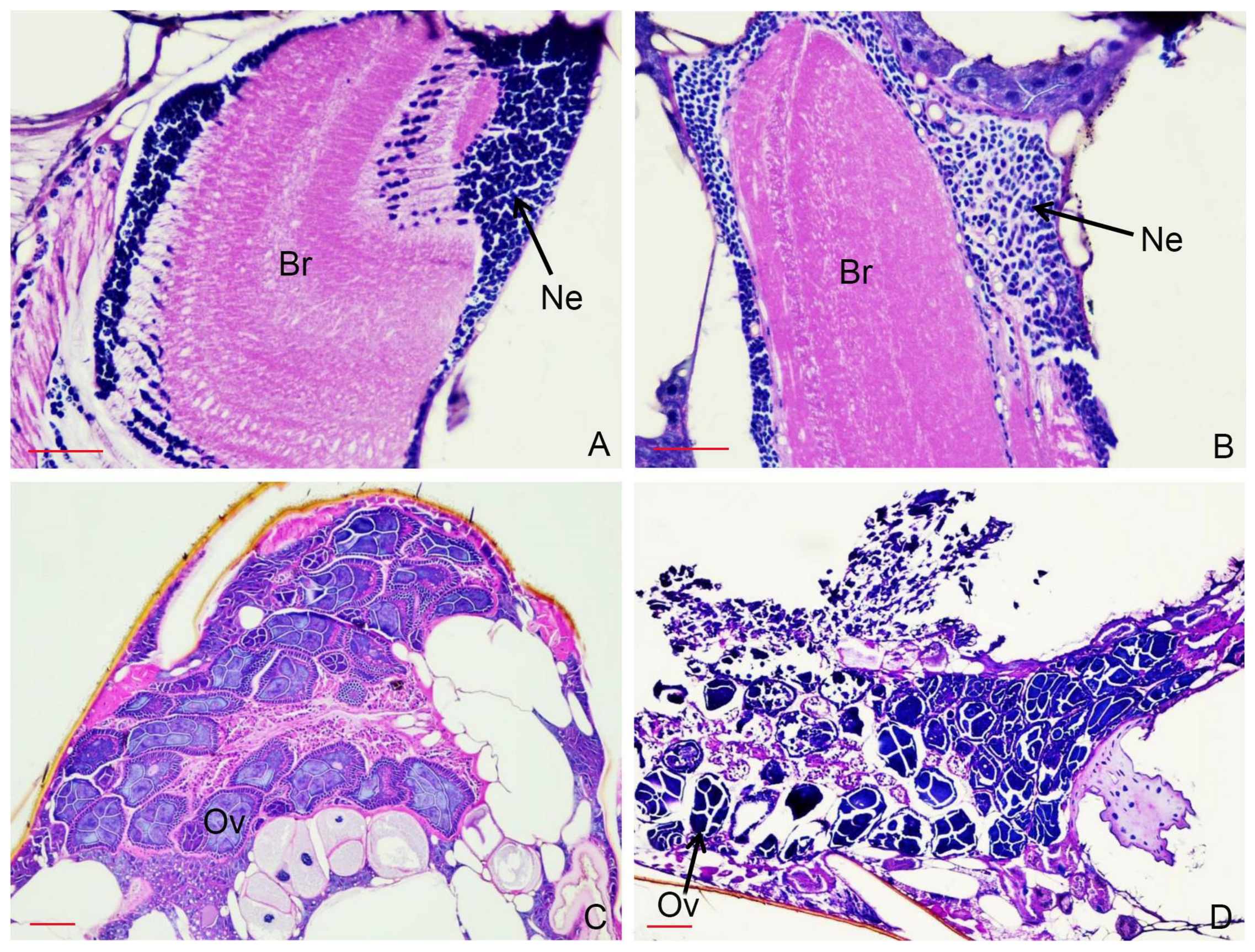
2.6. Histopathological Study
2.7. Scanning Electron Microscopic Study
2.8. Data Analysis
3. Results
3.1. Essential Oil Extraction and Quantification
3.2. Contact Toxicity Test
3.3. Fumigant Toxicity Test
3.4. Histopathological Study
3.5. Scanning Electron Microscopy
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Tuntasuvan, D.; Luckins, A.G. Status of surra in livestock in Thailand. J. Protozool. Res. 1998, 8, 162–170. [Google Scholar]
- Changbunjong, T.; Weluwanarak, T.; Ratanakorn, P.; Maneeon, P.; Ganpanakngan, M.; Apiwathnasorn, C.; Sungvornyothin, S.; Sriwichai, P.; Sumruayphol, S.; Ruangsittichai, J. Distribution and abundance of Stomoxyini flies (Diptera: Muscidae) in Thailand. Southeast Asian J. Trop. Med. Public Health 2012, 43, 1400–1410. [Google Scholar] [PubMed]
- Desquesnes, M.; Holzmuller, P.; Lai, D.H.; Dargantes, A.; Lun, Z.R.; Jittaplapong, S. Trypanosoma evansi and surra: A review and perspectives on origin, history, distribution, taxonomy, morphology, hosts, and pathogenic effects. Biomed. Res. Int. 2013, 2013, 194176. [Google Scholar] [CrossRef] [Green Version]
- Changbunjong, T.; Sedwisi, P.; Weluwanarak, T.; Nitiyamatawat, E.; Sariwongchan, R.; Chareonviriyaphap, T. Species diversity and abundance of Tabanus spp. (Diptera: Tabanidae) in different habitats of Thailand. J. Asia. Pac. Entomol. 2018, 21, 134–139. [Google Scholar] [CrossRef]
- Taylor, D.B.; Moon, R.D.; Mark, D.R. Economic impact of stable flies (Diptera: Muscidae) on dairy and beef cattle production. J. Med. Entomol. 2012, 49, 198–209. [Google Scholar] [CrossRef] [PubMed]
- Baldacchino, F.; Muenworn, V.; Desquesnes, M.; Desoli, F.; Charoenviriyaphap, T.; Duvallet, G. Transmission of pathogens by Stomoxys flies (Diptera, Muscidae): A review. Parasite 2013, 20, 26. [Google Scholar] [CrossRef] [Green Version]
- Baldacchino, F.; Desquesnes, M.; Mihok, S.; Foil, L.D.; Duvallet, G.; Jittapalapong, S. Tabanids: Neglected subjects of research, but important vectors of disease agents. Infect. Genet. Evol. 2014, 28, 596–615. [Google Scholar] [CrossRef]
- Sprygin, A.; Pestova, Y.; Wallace, D.B.; Tuppurainen, E.; Kononov, A.V. Transmission of lumpy skin disease virus: A short review. Virus Res. 2019, 269, 197637. [Google Scholar] [CrossRef]
- Foil, L.D.; Hogsette, J.A. Biology and control of tabanids, stable flies and horn flies. Rev. Sci. Technol. 1994, 13, 1125–1158. [Google Scholar] [CrossRef]
- Cook, D. A historical review of management options used against the stable fly (Diptera: Muscidae). Insects 2020, 11, 313. [Google Scholar] [CrossRef]
- Cilek, J.E.; Greene, G.L. Stable fly (Diptera: Muscidae) insecticide resistance in Kansas cattle feedlots. J. Econ. Entomol. 1994, 87, 275–279. [Google Scholar] [CrossRef]
- Barros, A.T.M.; Rodrigues, V.D.; Cançado, P.H.D.; Domingues, L.N. Resistance of the stable fly, Stomoxys calcitrans (Diptera: Muscidae), to cypermethrin in outbreak areas in Midwestern Brazil. Rev. Bras. Parasitol. Vet. 2019, 28, 802–806. [Google Scholar] [CrossRef] [Green Version]
- Reissert-Oppermann, S.; Bauer, B.; Steuber, S.; Clausen, P.H. Insecticide resistance in stable flies (Stomoxys calcitrans) on dairy farms in Germany. Parasitol. Res. 2019, 118, 2499–2507. [Google Scholar] [CrossRef]
- Showler, A.T. Botanically based repellent and insecticidal effects against horn flies and stable flies (Diptera: Muscidae). J. Integr. Pest Manag. 2017, 8, 15. [Google Scholar] [CrossRef] [Green Version]
- Tlak Gajger, I.; Dar, S.A. Plant allelochemicals as sources of insecticides. Insects 2021, 12, 189. [Google Scholar] [CrossRef]
- Chotchoungchatchai, S.; Saralamp, P.; Jenjittikul, T.; Pornsiripongse, S.; Prathanturarug, S. Medicinal plants used with Thai traditional medicine in modern healthcare services: A case study in Kabchoeng Hospital, Surin Province, Thailand. J. Ethnopharmacol. 2012, 141, 193–205. [Google Scholar] [CrossRef]
- Maneenoon, K.; Khuniad, C.; Teanuan, Y.; Saedan, N.; Prom-In, S.; Rukleng, N.; Kongpool, W.; Pinsook, P.; Wongwiwat, W. Ethnomedicinal plants used by traditional healers in Phatthalung Province, Peninsular Thailand. J. Ethnobiol. Ethnomed. 2015, 11, 43. [Google Scholar] [CrossRef] [Green Version]
- Pintong, A.R.; Ampawong, S.; Komalamisra, N.; Sriwichai, P.; Popruk, S.; Ruangsittichai, J. Insecticidal and histopathological effects of Ageratum conyzoides weed extracts against dengue vector. Aedes Aegypti. Insects 2020, 11, 224. [Google Scholar] [CrossRef] [Green Version]
- Suwannayod, S.; Sukontason, K.L.; Somboon, P.; Junkum, A.; Leksomboon, R.; Chaiwong, T.; Jones, M.K.; Sripa, B.; Balthaisong, S.; Phuyao, C.; et al. Activity of kaffirlime (Citrus hystrix) essential oil against blow flies and house fly. Southeast Asian J. Trop. Med. Public Health 2018, 49, 32–45. [Google Scholar]
- Ebadollahi, A.; Ziaee, M.; Palla, F. Essential oils extracted from different species of the Lamiaceae plant family as prospective bioagents against several detrimental pests. Molecules 2020, 25, 1556. [Google Scholar] [CrossRef] [Green Version]
- Arumugam, G.; Swamy, M.K.; Sinniah, U.R. Plectranthus amboinicus (Lour.) Spreng: Botanical, phytochemical, pharmacological and nutritional significance. Molecules 2016, 21, 369. [Google Scholar] [CrossRef]
- Wadikar, D.D.; Patki, P.E. Coleus aromaticus: A therapeutic herb with multiple potentials. J. Food Sci. Technol. 2016, 53, 2895–2901. [Google Scholar] [CrossRef] [Green Version]
- Jimmy, J.L. Coleus aromaticus Benth.: An update on its bioactive constituents and medicinal properties. All Life 2021, 14, 756–773. [Google Scholar] [CrossRef]
- Singh, G.; Singh, O.P.; Prasad, Y.R.; Lamposona, M.P.; Catalan, C. Studies on essential oils. Part 33. chemical and insecticidal investigations on leaf oil of Coleus amboinicus (Lour). Flavour. Frag. J. 2002, 17, 440–442. [Google Scholar] [CrossRef]
- Satongrod, B.; Wanna, R. Chemical composition and bioactivity of essential oil from Indian borage (Plectranthus amboincus (Lour.) Spreng) against Callosobruchus maculatus (F.). Int. J. Agric. Technol. 2020, 16, 1243–1256. [Google Scholar]
- Lima, M.A.; Oliveira, F.F.M.; Gomes, G.A.; Lavor, P.L.; Santiago, G.M.; Nagao-Dias, A.T.; Arriaga, A.M.; Lemos, T.L.; Carvalho, M.G. Evaluation of larvicidal activity of the essential oils of plants species from Brazil against Aedes aegypti (Diptera: Culicidae). Afr. J. Biotechnol. 2011, 10, 11716–11720. [Google Scholar]
- Verma, R.S.; Padalia, R.C.; Chauhan, A. Essential oil composition of Coleus aromaticus Benth. from Uttarakhand. J. Essent. Oil Bear. Plant 2012, 15, 174–179. [Google Scholar] [CrossRef]
- Singh, J.P.; Prakash, B.; Dubey, N.K. Insecticidal activity of Ageratum conyzoides L., Coleus aromaticus Benth. and Hyptis suaveolens (L.) Poit essential oils as fumigant against storage grain insect Tribolium castaneum Herbst. J. Food Sci. Technol. 2014, 51, 2210–2215. [Google Scholar]
- Singh, M.; Lal, K.; Singh, S.B.; Singh, M. Effect of Calotropis (Calotropis procera) extract on infestation of termite (Odontotermes obesus) in sugarcane hybrid. Indian J. Agric. Sci. 2002, 72, 439–441. [Google Scholar]
- Mihok, S. The development of a multipurpose trap (the Nzi) for tsetse and other biting flies. Bull. Entomol. Res. 2002, 92, 385–403. [Google Scholar] [CrossRef] [PubMed]
- Tunnakundacha, S.; Desquesnes, M.; Masmeatathip, R. Comparison of Vavoua, Malaise and Nzi traps with and without attractants for trapping of Stomoxys spp. (Diptera: Muscidae) and tabanids (Diptera: Tabanidae) on cattle farms. Agric. Nat. Resour. 2017, 51, 319–323. [Google Scholar] [CrossRef]
- WHO. Test Procedures for Insecticide Resistance Monitoring in Malaria Vector Mosquitoes; World Health Organization: Geneva, Switzerland, 2016; ISBN 9783642105654. [Google Scholar]
- Zhu, J.J.; Li, A.Y.; Pritchard, S.; Tangtrakulwanich, K.; Baxendale, F.P.; Brewer, G. Contact and fumigant toxicity of a botanical-based feeding deterrent of the stable fly, Stomoxys calcitrans (Diptera: Muscidae). J. Agric. Food Chem. 2011, 59, 10394–10400. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abbott, W.S. A method of computing the effectiveness of an insecticide. J. Econ. Entomol. 1925, 18, 265–267. [Google Scholar] [CrossRef]
- Karakus, M.; Gocmen, B.; Özbel, Y. Insecticide susceptibility status of wild-caught sand fly populations collected from two leishmaniasis endemic areas in western Turkey. J. Arthropod Borne Dis. 2017, 11, 86–94. [Google Scholar]
- González, M.A.; Bell, M.J.; Bernhardt, S.A.; Brazil, R.P.; Dilger, E.; Courtenay, O.; Hamilton, J.G.C. Susceptibility of wild-caught Lutzomyia longipalpis (Diptera: Psychodidae) sand flies to insecticide after an extended period of exposure in western São Paulo, Brazil. Parasit. Vectors. 2019, 12, 110. [Google Scholar] [CrossRef] [Green Version]
- Tainchum, K.; Shukri, S.; Duvallet, G.; Etienne, L.; Jacquiet, P. Phenotypic susceptibility to pyrethroids and organophosphate of wild Stomoxys calcitrans (Diptera: Muscidae) populations in southwestern France. Parasitol. Res. 2018, 117, 4027–4032. [Google Scholar] [CrossRef]
- Shivanandappa, T.; Rajashekar, Y. Mode of action of plant-derived natural insecticides. In Advances in Plant Biopesticides; Singh, D., Ed.; Springer: New Delhi, India, 2014; pp. 323–345. [Google Scholar]
- Dillmann, J.B.; Cossetin, L.F.; de Giacometi, M.; Oliveira, D.; de Matos, A.F.I.M.; Avrella, P.D.; Garlet, Q.I.; Heinzmann, B.M.; Monteiro, S.G. Adulticidal activity of Melaleuca alternifolia (Myrtales: Myrtaceae) essential oil with high 1,8-Cineole content against stable flies (Diptera: Muscidae). J. Econ. Entomol. 2020, 113, 1810–1815. [Google Scholar] [CrossRef]
- Hieu, T.T.; Kim, S.I.; Ahn, Y.J. Toxicity of Zanthoxylum piperitum and Zanthoxylum armatum oil constituents and related compounds to Stomoxys calcitrans (Diptera: Muscidae). J. Med. Entomol. 2012, 49, 1084–1091. [Google Scholar] [CrossRef] [Green Version]
- Adorjan, B.; Buchbauer, G. Biological properties of essential oils: An updated review. Flavour. Fragr. J. 2010, 25, 407–426. [Google Scholar] [CrossRef]
- Reis, S.L.; Mantello, A.G.; Macedo, J.M.; Gelfuso, E.A.; da Silva, C.P.; Fachin, A.L.; Cardoso, A.M.; Beleboni, R.O. Typical monoterpenes as insecticides and repellents against stored grain pests. Molecules 2016, 21, 258. [Google Scholar] [CrossRef] [Green Version]
- Youssefi, M.R.; Tabari, M.A.; Esfandiari, A.; Kazemi, S.; Moghadamnia, A.A.; Sut, S.; Acqua, S.D.; Benelli, G.; Maggi, F. Efficacy of two monoterpenoids, carvacrol and thymol, and their combinations against eggs and larvae of the West Nile Vector Culex pipiens. Molecules 2019, 24, 1867. [Google Scholar] [CrossRef] [Green Version]
- Park, J.H.; Jeon, Y.J.; Lee, C.H.; Chung, N.; Lee, H.S. Insecticidal toxicities of carvacrol and thymol derived from Thymus vulgaris Lin. against Pochazia shantungensis Chou & Lu., newly recorded pest. Sci. Rep. 2017, 7, 40902. [Google Scholar]
- Gaire, S.; Scharf, M.E.; Gondhalekar, A.D. Toxicity and neurophysiological impacts of plant essential oil components on bed bugs (Cimicidae: Hemiptera). Sci. Rep. 2019, 9, 3961. [Google Scholar] [CrossRef] [Green Version]
- Dias, M.L.; Auad, A.M.; Magno, M.C.; Resende, T.T.; Fonseca, M.G.; Silva, S.E.B. Insecticidal activity of compounds of plant origin on Mahanarva spectabilis (Hemiptera: Cercopidae). Insects 2019, 10, 360. [Google Scholar] [CrossRef] [Green Version]
- Ma, S.; Jia, R.; Guo, M.; Qin, K.; Zhang, L. Insecticidal activity of essential oil from Cephalotaxus sinensis and its main components against various agricultural pests. Ind. Crops Prod. 2020, 150, 112403. [Google Scholar] [CrossRef]
- Tong, F.; Coats, J.R. Effects of monoterpenoid insecticides on [3H]-TBOB binding in house fly GABA receptor and 36Cl− uptake in American cockroach ventral nerve cord. Pestic. Biochem. Physiol. 2010, 98, 317–324. [Google Scholar] [CrossRef]
- Tong, F.; Coats, J.R. Quantitative structure–activity relationships of monoterpenoid binding activities to the housefly GABA receptor. Pest Manag. Sci. 2012, 68, 1122–1129. [Google Scholar] [CrossRef] [Green Version]
- Tong, F.; Gross, A.D.; Dolan, M.C.; Coats, J.R. The phenolic monoterpenoid carvacrol inhibits the binding of nicotine to the housefly nicotinic acetylcholine receptor. Pest Manag. Sci. 2013, 69, 775–780. [Google Scholar] [CrossRef] [Green Version]
- Glavan, G.; Novak, S.; Božič, J.; Jemec, K.A. Comparison of sublethal effects of natural acaricides carvacrol and thymol on honeybees. Pestic. Biochem. Physiol. 2020, 166, 104567. [Google Scholar] [CrossRef]
- Orhan, I.; Kartal, M.; Kan, Y.; Şener, B. Activity of essential oils and individual components against acetyl and butyrylcholinesterase. Z. Nat. C 2008, 63, 547–553. [Google Scholar]
- Anderson, J.A.; Coats, J.R. Acetylcholinesterase inhibition by nootkatone and carvacrol in arthropods. Pestic. Biochem. Physiol. 2012, 102, 124–128. [Google Scholar] [CrossRef] [Green Version]
- Parnas, M.; Peters, M.; Dadon, D.; Lev, S.; Vertkin, I.; Slutsky, I.; Minke, B. Carvacrol is a novel inhibitor of Drosophila TRPL and mammalian TRPM7 channels. Cell Calcium 2009, 45, 300–309. [Google Scholar] [CrossRef] [Green Version]
- Waliwitiya, R.; Belton, P.; Nicholson, R.A.; Lowenberger, C.A. Effects of the essential oil constituent thymol and other neuroactive chemicals on flight motor activity and wing beat frequency in the blowfly Phaenicia. Sericata. Pest Manag. Sci. 2010, 66, 277–289. [Google Scholar] [CrossRef] [PubMed]
- Prasantha, B.D.R.; Reichmuth, C.; Adler, C.; Felgentreu, D. Lipid adsorption of diatomaceous earths and increased water permeability in the epicuticle layer of the cowpea weevil Callosobruchus maculatus (F.) and the bean weevil Acanthoscelides obtectus (Say) (Chrysomelidae). J. Stored Prod. Res. 2015, 64, 36–41. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Concentration (µg/µL) | Mortality (%) | |||||
|---|---|---|---|---|---|---|
| 1 h | 2 h | 4 h | 6 h | 12 h | 24 h | |
| Negative control | 0 a | 0 a | 0 a | 0 a | 0 a | 0 a |
| Cypermethrin (1%) | 100 d | 100 c | 100 c | 100 c | 100 d | 100 c |
| 9.3 | 0 a | 0 a | 13.3 ± 15.3 ab | 13.3 ± 15.3 a | 16.7 ± 11.6 ab | 26.7 ± 11.6 b |
| 18.7 | 3.3 ± 5.8 a | 3.3 ± 5.8 a | 6.7 ± 5.8 ab | 13.3 ± 15.3 a | 20.0 ± 10.0 ab | 33.3 ± 5.8 b |
| 37.4 | 16.7 ± 11.6 ab | 16.7 ± 11.6 ab | 23.3 ± 15.3 ab | 23.3 ± 15.3 ab | 30.0 ± 10.0 b | 50.0 ± 17.3 b |
| 46.7 | 33.3 ± 15.3 b | 36.7 ± 11.6 b | 36.7 ± 11.6 b | 50.0 ± 10.0 b | 63.3 ± 15.3 c | 80.0 ± 10.0 c |
| 93.4 | 70.0 ± 17.3 c | 73.3 ± 20.8 c | 73.3 ± 20.8 c | 83.3 ± 5.8 c | 83.3 ± 5.8 cd | 96.7 ± 5.8 c |
| df | 6, 14 | 6, 14 | 6, 14 | 6, 14 | 6, 14 | 6, 14 |
| F | 45.873 | 45.121 | 27.302 | 37.280 | 49.630 | 50.796 |
| p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Concentration (µg/µL) | Mortality (%) | |||||
|---|---|---|---|---|---|---|
| 1 h | 2 h | 4 h | 6 h | 12 h | 24 h | |
| Negative control | 0 a | 0 a | 0 a | 0 a | 0 a | 0 a |
| Cypermethrin (1%) | 100 b | 100 d | 100 d | 100 d | 100 d | 100 e |
| 93.4 | 0 a | 0 a | 0 a | 3.3 ± 5.78 a | 6.7 ± 5.8 a | 10.0 ± 0.0 a |
| 116.8 | 6.8 ± 5.8 a | 10.0 ± 0.0 ab | 13.3 ± 5.8 ab | 20.0 ± 10.0 ab | 43.3 ± 5.8 b | 43.3 ± 5.8 b |
| 140.1 | 10.0 ± 0.0 a | 20.0 ± 17.3 ab | 26.7 ± 20.8 abc | 26.7 ± 20.8 abc | 56.7 ± 5.8 bc | 60.0 ± 0.0 c |
| 163.5 | 16.7 ± 15.3 a | 33.3 ± 15.3 bc | 36.7 ± 11.6 bc | 43.3 ± 15.1 bc | 66.7 ± 11.6 c | 73.3 ± 5.8 d |
| 186.8 | 20.0 ± 10.0 a | 53.3 ± 15.3 c | 56.7 ± 15.3 c | 56.7 ± 15.3 c | 73.3 ± 5.8 c | 83.3 ± 5.8 d |
| df | 6, 14 | 6, 14 | 6, 14 | 6, 14 | 6, 14 | 6, 14 |
| F | 71.242 | 35.377 | 32.200 | 24.667 | 102.208 | 292.000 |
| p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Treatment | Contact Toxicity Test | Fumigant Toxicity Test | ||
|---|---|---|---|---|
| S. calcitrans | T. megalops | S. calcitrans | T. megalops | |
| LD50 [µg/fly] (95% CL) | 12.05 (9.15–15.18) | 131.41 (121.05–141.50) | - | - |
| LD90 [µg/fly] (95% CL) | 45.53 (32.22–83.64) | 200.62 (178.95–264.20) | - | - |
| LC50 [mg/L air] (95% CL) | - | - | 1.34 (1.05–1.68) | 7.12 (5.33–9.20) |
| LC90 [mg/L air] (95% CL) | - | - | 4.39 (3.18–7.57) | 30.37 (24.82–57.61) |
| Slope ± SE | 2.21 ± 0.36 | 6.97 ± 1.14 | 2.49 ± 0.39 | 2.03 ± 0.91 |
| χ2 | 7.74 | 1.52 | 2.94 | 3.85 |
| Concentration (mg/L Air) | Mortality (%) | |||||
|---|---|---|---|---|---|---|
| 1 h | 2 h | 4 h | 6 h | 12 h | 24 h | |
| Negative control | 0 a | 0 a | 0 a | 0 a | 0 a | 0 a |
| Cypermethrin (1%) | 100 b | 100 b | 100 d | 100 b | 100 e | 100 c |
| 0.47 | 3.3 ± 5.8 a | 3.3 ± 5.8 a | 3.3 ± 5.8 ab | 6.7 ± 5.8 a | 6.7 ± 5.8 ab | 10.0 ± 10.0 a |
| 0.93 | 0 a | 0 a | 3.3 ± 5.8 ab | 26.7 ± 15.3 a | 26.7 ± 15.3 bc | 36.7 ± 23.1 ab |
| 1.87 | 0 a | 3.3 ± 5.8 a | 6.7 ± 5.8 ab | 26.7 ± 15.3 a | 36.7 ± 5.8 c | 63.3 ± 15.3 b |
| 2.80 | 3.3 ± 5.8 a | 3.3 ± 5.8 a | 16.7 ± 5.8 bc | 33.3 ± 25.2 a | 73.3 ± 5.8 d | 86.7 ± 5.8 c |
| 4.67 | 6.7 ± 5.8 a | 10.0 ± 0.0 a | 23.3 ± 5.8 c | 73.3 ± 15.3 c | 76.7 ± 11.6 d | 93.3 ± 11.6 c |
| df | 6, 14 | 6, 14 | 6, 14 | 6, 14 | 6, 14 | 6, 14 |
| F | 288.111 | 282.667 | 158.133 | 19.967 | 65.452 | 24.295 |
| p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
| Concentration (mg/L Air) | Mortality (%) | |||||
|---|---|---|---|---|---|---|
| 1 h | 2 h | 4 h | 6 h | 12 h | 24 h | |
| Negative control | 0 a | 0 a | 0 a | 0 a | 0 a | 0 a |
| Cypermethrin (1%) | 100 b | 100 b | 100 b | 100 c | 100 d | 100 d |
| 2.34 | 0 a | 0 a | 0 a | 3.3 ± 5.8 a | 10.0 ± 10.0 ab | 13.3 ± 11.6 a |
| 4.67 | 0 a | 0 a | 0 a | 0 a | 23.3 ± 11.6 b | 46.7±11.6 b |
| 9.34 | 0 a | 0 a | 0 a | 16.7 ± 5.8 b | 50.0 ± 0.0 c | 50.0 ± 0.0 b |
| 18.68 | 0 a | 0 a | 0 a | 3.3 ± 5.8 a | 46.7 ± 11.6 c | 76.7 ± 5.8 c |
| 28.02 | 3.3 ± 5.8 a | 6.7 ± 11.6 a | 6.7 ± 11.6 a | 16.7 ± 5.8 b | 86.7 ± 15.1 d | 93.3 ± 5.8 cd |
| df | 6, 14 | 6, 14 | 6, 14 | 6, 14 | 6, 14 | 6, 14 |
| F | 891.000 | 221.000 | 221.000 | 204.167 | 79.611 | 92.567 |
| p-value | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 | <0.001 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Leesombun, A.; Sungpradit, S.; Boonmasawai, S.; Weluwanarak, T.; Klinsrithong, S.; Ruangsittichai, J.; Ampawong, S.; Masmeatathip, R.; Changbunjong, T. Insecticidal Activity of Plectranthus amboinicus Essential Oil against the Stable Fly Stomoxys calcitrans (Diptera: Muscidae) and the Horse Fly Tabanus megalops (Diptera: Tabanidae). Insects 2022, 13, 255. https://doi.org/10.3390/insects13030255
Leesombun A, Sungpradit S, Boonmasawai S, Weluwanarak T, Klinsrithong S, Ruangsittichai J, Ampawong S, Masmeatathip R, Changbunjong T. Insecticidal Activity of Plectranthus amboinicus Essential Oil against the Stable Fly Stomoxys calcitrans (Diptera: Muscidae) and the Horse Fly Tabanus megalops (Diptera: Tabanidae). Insects. 2022; 13(3):255. https://doi.org/10.3390/insects13030255
Chicago/Turabian StyleLeesombun, Arpron, Sivapong Sungpradit, Sookruetai Boonmasawai, Thekhawet Weluwanarak, Suriyo Klinsrithong, Jiraporn Ruangsittichai, Sumate Ampawong, Roungthip Masmeatathip, and Tanasak Changbunjong. 2022. "Insecticidal Activity of Plectranthus amboinicus Essential Oil against the Stable Fly Stomoxys calcitrans (Diptera: Muscidae) and the Horse Fly Tabanus megalops (Diptera: Tabanidae)" Insects 13, no. 3: 255. https://doi.org/10.3390/insects13030255