
Mutualism between Gut-Borne Yeasts and Their Host, Thaumatotibia leucotreta, and Potential Usefulness in Pest Management

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Yeast Isolation and Purification
2.2. Yeast Identification
2.3. Thaumatotibia leucotreta Culture
2.4. Larval Development Assays
2.5. Larval Feeding Assay
2.6. Oviposition Preference Assays
2.7. Statistical Analysis
3. Results
3.1. Yeast Isolation and Identification
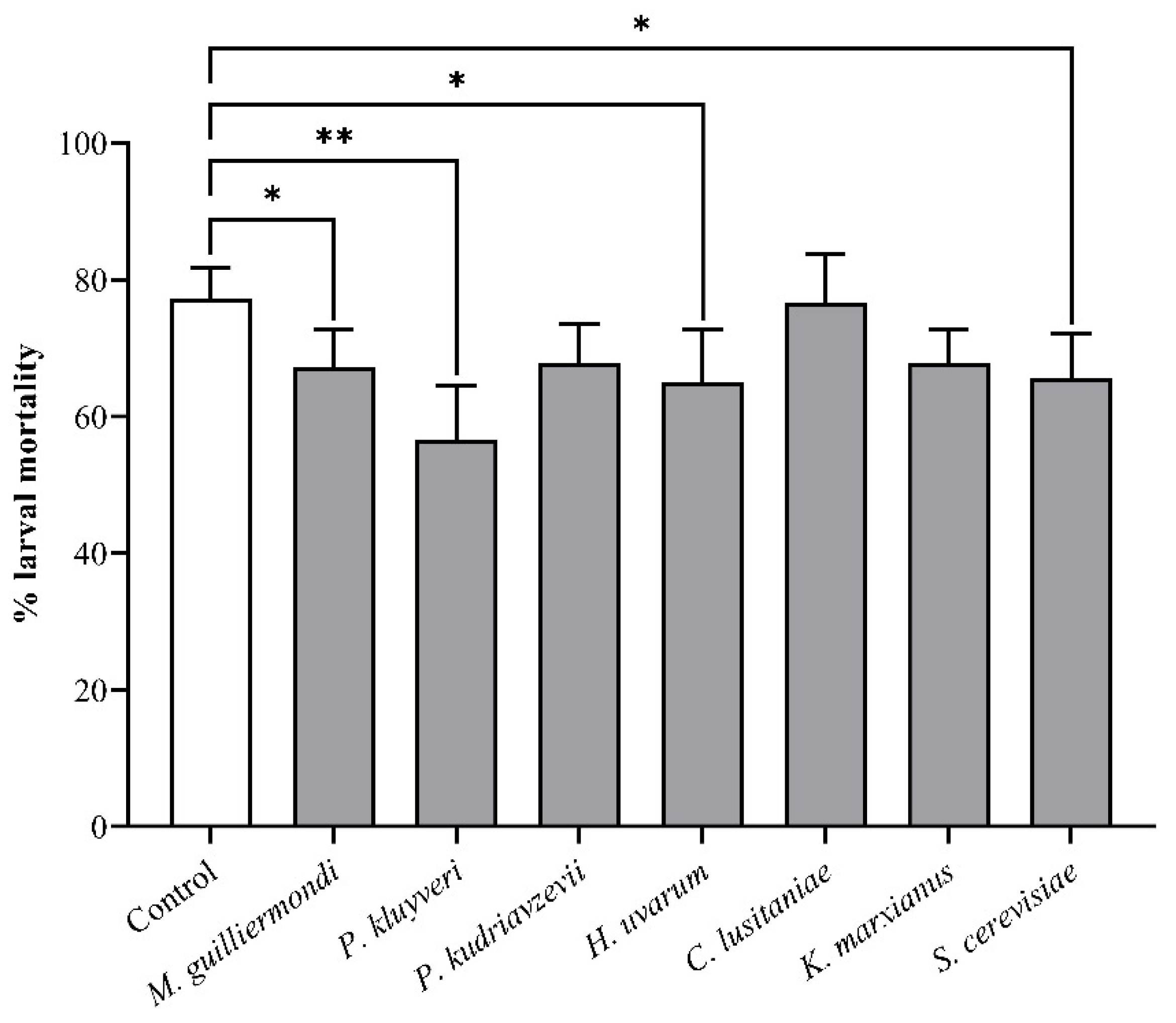
3.2. Larval Development Assays
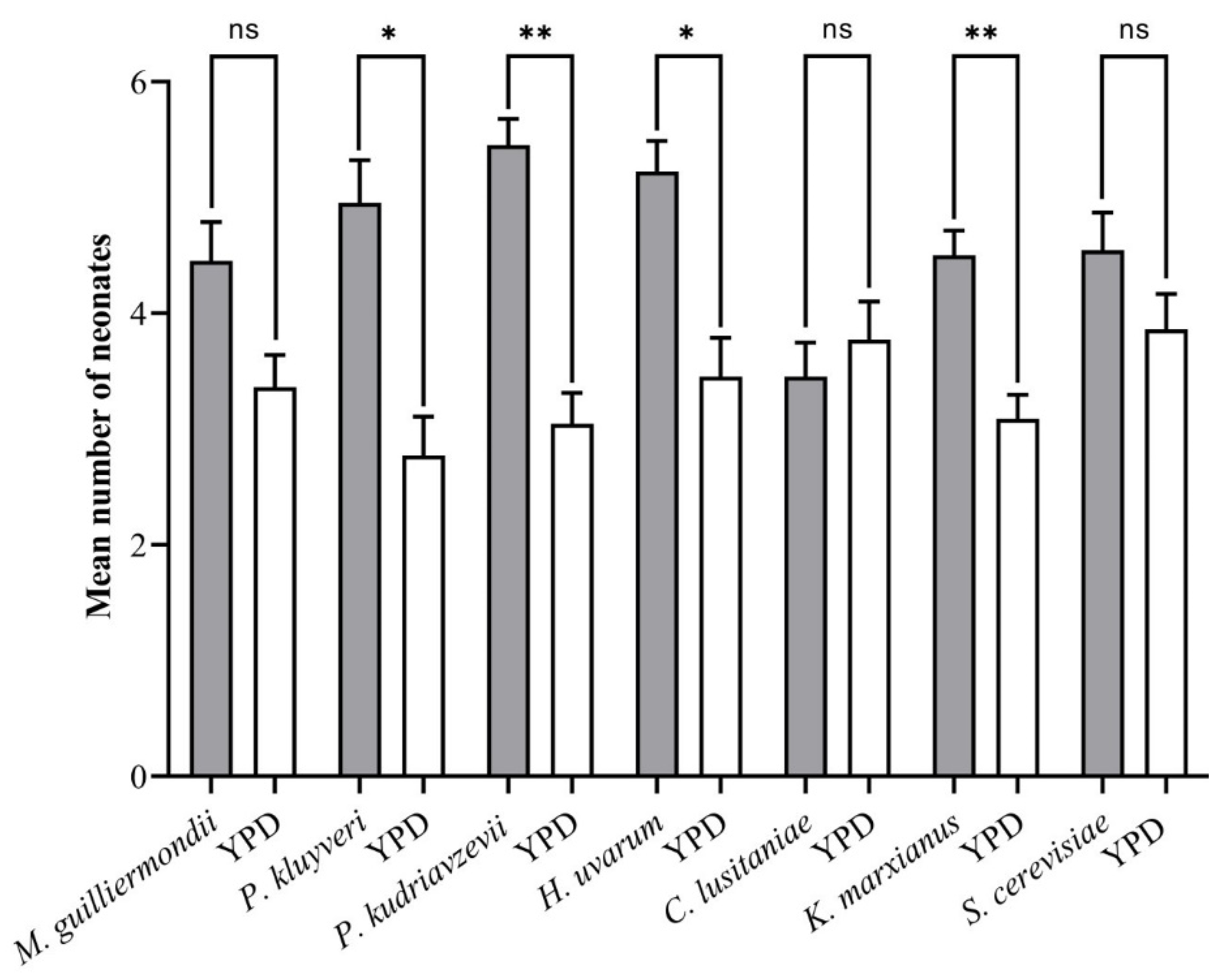
3.3. Larval Feeding Assay
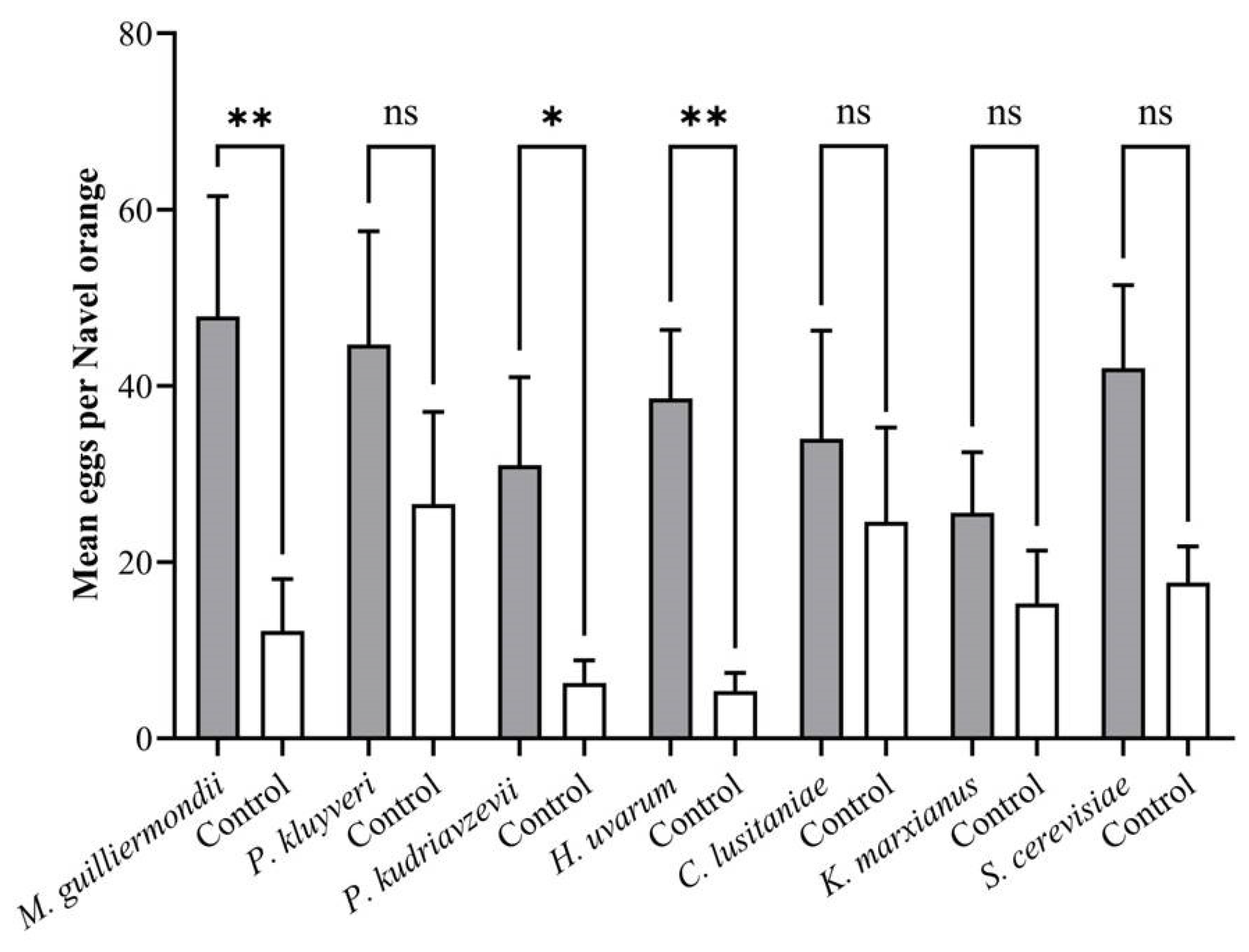
3.4. Oviposition Preference Assays
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hosokawa, T.; Fukatsu, T. Relevance of microbial symbiosis to insect behavior. Curr. Opin. Insect Sci. 2020, 39, 91–100. [Google Scholar] [CrossRef]
- Douglas, A.E. Multiorganismal Insects: Diversity and Function of Resident Microorganisms. Annu. Rev. Entomol. 2015, 60, 17–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Engel, P.; Moran, N.A. The gut microbiota of insects—Diversity in structure and function. FEMS Microbiol. Rev. 2013, 37, 699–735. [Google Scholar] [CrossRef]
- Gupta, A.; Nair, S. Dynamics of Insect–Microbiome Interaction Influence Host and Microbial Symbiont. Front. Microbiol. 2020, 11, 1357. [Google Scholar] [CrossRef] [PubMed]
- Becher, P.G.; Hagman, A.; Verschut, V.; Chakraborty, A.; Rozpędowska, E.; Lebreton, S.; Bengtsson, M.; Flick, G.; Witzgall, P.; Piškur, J. Chemical signaling and insect attraction is a conserved trait in yeasts. Ecol. Evol. 2018, 8, 2962–2974. [Google Scholar] [CrossRef] [Green Version]
- Madden, A.A.; Epps, M.J.; Fukami, T.; Irwin, R.E.; Sheppard, J.; Sorger, D.M.; Dunn, R.R. The ecology of insect–yeast relationships and its relevance to human industry. Proc. R. Soc. B Biol. Sci. 2018, 285, 20172733. [Google Scholar] [CrossRef] [Green Version]
- Douglas, A.E. The Molecular Basis of Bacterial–Insect Symbiosis. J. Mol. Biol. 2014, 426, 3830–3837. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martino, M.E.; Ma, D.; Leulier, F. Microbial influence on Drosophila biology. Curr. Opin. Microbiol. 2017, 38, 165–170. [Google Scholar] [CrossRef]
- Gurung, K.; Wertheim, B.; Salles, J.F. The microbiome of pest insects: It is not just bacteria. Entomol. Exp. Appl. 2019, 167, 156–170. [Google Scholar] [CrossRef] [Green Version]
- Wang, S.; Wang, L.; Fan, X.; Yu, C.; Feng, L.; Yi, L. An Insight into Diversity and Functionalities of Gut Microbiota in Insects. Curr. Microbiol. 2020, 77, 1976–1986. [Google Scholar] [CrossRef] [PubMed]
- Barraclough, T.G. How Do Species Interactions Affect Evolutionary Dynamics Across Whole Communities? Annu. Rev. Ecol. Evol. Syst. 2015, 46, 25–48. [Google Scholar] [CrossRef]
- Chandler, D.; Bailey, A.S.; Tatchell, G.M.; Davidson, G.; Greaves, J.; Grant, W.P. The development, regulation and use of biopesticides for integrated pest management. Philos. Trans. R. Soc. B Biol. Sci. 2011, 366, 1987–1998. [Google Scholar] [CrossRef] [PubMed]
- McFall-Ngai, M.; Hadfield, M.G.; Bosch, T.C.G.; Carey, H.V.; Domazet-Lošo, T.; Douglas, A.E.; Dubilier, N.; Eberl, G.; Fukami, T.; Gilbert, S.F.; et al. Animals in a bacterial world, a new imperative for the life sciences. Proc. Natl. Acad. Sci. USA 2013, 110, 3229–3236. [Google Scholar] [CrossRef] [Green Version]
- Murgier, J.; Everaerts, C.; Farine, J.-P.; Ferveur, J.-F. Live yeast in juvenile diet induces species-specific effects on Drosophila adult behaviour and fitness. Sci. Rep. 2019, 9, 8873. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yun, J.-H.; Roh, S.W.; Whon, T.W.; Jung, M.-J.; Kim, M.-S.; Park, D.-S.; Yoon, C.; Nam, Y.-D.; Kim, Y.-J.; Choi, J.-H.; et al. Insect Gut Bacterial Diversity Determined by Environmental Habitat, Diet, Developmental Stage, and Phylogeny of Host. Appl. Environ. Microbiol. 2014, 80, 5254–5264. [Google Scholar] [CrossRef] [Green Version]
- MsangoSoko, K.; Gandotra, S.; Chandel, R.K.; Sharma, K.; Ramakrishinan, B.; Subramanian, A.S. Composition and Diversity of Gut Bacteria Associated with the Eri Silk Moth, Samia ricini, (Lepidoptera: Saturniidae) as Revealed by Culture-Dependent and Metagenomics Analysis. J. Microbiol. Biotechnol. 2020, 30, 1367–1378. [Google Scholar] [CrossRef]
- Paniagua Voirol, L.R.; Frago, E.; Kaltenpoth, M.; Hilker, M.; Fatouros, N.E. Bacterial Symbionts in Lepidoptera: Their Diversity, Transmission, and Impact on the Host. Front. Microbiol. 2018, 9, 556. [Google Scholar] [CrossRef]
- Sree, K.S.; Varma, A. (Eds.) Biocontrol of Lepidopteran Pests: Use of Soil Microbes and Their Metabolites; Soil Biology; Springer International Publishing: Cham, Switzerland, 2015; ISBN 978-3-319-14498-6. [Google Scholar]
- Stefanini, I. Yeast-insect associations: It takes guts. Yeast 2018, 35, 315–330. [Google Scholar] [CrossRef] [Green Version]
- Zchori-Fein, E.; Bourtzis, K. Manipulative Tenants: Bacteria Associated with Arthropods; CRC Press: Boca Raton, FL, USA, 2011; ISBN 978-1-4398-2749-9. [Google Scholar]
- Günther, C.S.; Goddard, M.R. Do yeasts and Drosophila interact just by chance? Fungal Ecol. 2019, 38, 37–43. [Google Scholar] [CrossRef] [Green Version]
- Bellutti, N.; Gallmetzer, A.; Innerebner, G.; Schmidt, S.; Zelger, R.; Koschier, E.H. Dietary yeast affects preference and performance in Drosophila suzukii. J. Pest Sci. 2017, 91, 651–660. [Google Scholar] [CrossRef] [Green Version]
- Spitaler, U.; Bianchi, F.; Eisenstecken, D.; Castellan, I.; Angeli, S.; Dordevic, N.; Robatscher, P.; Vogel, R.F.; Koschier, E.H.; Schmidt, S. Yeast species affects feeding and fitness of Drosophila suzukii adults. J. Pest Sci. 2020, 93, 1295–1309. [Google Scholar] [CrossRef]
- Becher, P.G.; Flick, G.; Rozpędowska, E.; Schmidt, A.; Hagman, A.; Lebreton, S.; Larsson, M.C.; Hansson, B.S.; Piškur, J.; Witzgall, P.; et al. Yeast, not fruit volatiles mediate Drosophila melanogaster attraction, oviposition and development. Funct. Ecol. 2012, 26, 822–828. [Google Scholar] [CrossRef]
- Grunseich, J.M.; Thompson, M.N.; Aguirre, N.M.; Helms, A.M. The Role of Plant-Associated Microbes in Mediating Host-Plant Selection by Insect Herbivores. Plants 2020, 9, 6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Witzgall, P.; Proffit, M.; Rozpedowska, E.; Becher, P.G.; Andreadis, S.; Coracini, M.; Lindblom, T.; Ream, L.J.; Hagman, A.; Bengtsson, M.; et al. “This is not an Apple”–Yeast Mutualism in Codling Moth. J. Chem. Ecol. 2012, 38, 949–957. [Google Scholar] [CrossRef]
- Knight, A.L.; Witzgall, P. Combining Mutualistic Yeast and Pathogenic Virus—A Novel Method for Codling Moth Control. J. Chem. Ecol. 2013, 39, 1019–1026. [Google Scholar] [CrossRef] [Green Version]
- Knight, A.L.; Basoalto, E.; Witzgall, P. Improving the Performance of the Granulosis Virus of Codling Moth (Lepidoptera: Tortricidae) by Adding the Yeast Saccharomyces cerevisiae with Sugar. Environ. Entomol. 2015, 44, 252–259. [Google Scholar] [CrossRef] [Green Version]
- Knight, A.L.; Basoalto, E.; Yee, W.; Hilton, R.; Kurtzman, C.P. Adding yeasts with sugar to increase the number of effective insecticide classes to manage Drosophila suzukii (Matsumura) (Diptera: Drosophilidae) in cherry: Adding Yeast and Sugar to Manage Spotted-Wing Drosophila. Pest Manag. Sci. 2016, 72, 1482–1490. [Google Scholar] [CrossRef]
- Bedford, E.C.G.; Van den Berg, M.A.; De Villiers, E.A. Citrus Pests in the Republic of South Africa; Institute for Tropical and Subtropical Crops; Outspan International: Nelspruit, South Africa, 1998; ISBN 978-0-620-22031-6. [Google Scholar]
- Hattingh, V.; Moore, S.; Kirkman, W.; Goddard, M.; Thackeray, S.; Peyper, M.; Sharp, G.; Cronjé, P.; Pringle, K. An Improved Systems Approach as a Phytosanitary Measure for Thaumatotibia leucotreta (Lepidoptera: Tortricidae) in Export Citrus Fruit from South Africa. J. Econ. Entomol. 2020, 113, 700–711. [Google Scholar] [CrossRef]
- Moore, S.D.; Grout, T.G.; Hattingh, V.; Hofmeyr, J.H. Thresholds and Guidelines for Intervention against Citrus Pests. S. Afr. Fruit J. 2008, 7, 77–81. [Google Scholar]
- Moore, S.D. Biological Control of a Phytosanitary Pest (Thaumatotibia leucotreta): A Case Study. Int. J. Environ. Res. Public Health 2021, 18, 1198. [Google Scholar] [CrossRef]
- Mkiga, A.M.; Mohamed, S.A.; Du Plessis, H.; Khamis, F.M.; Ekesi, S. Field and Laboratory Performance of False Codling Moth, Thaumatotibia leucotreta (Lepidoptera: Troticidae) on Orange and Selected Vegetables. Insects 2019, 10, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hatting, J.L.; Moore, S.D.; Malan, A.P. Microbial control of phytophagous invertebrate pests in South Africa: Current status and future prospects. J. Invertebr. Pathol. 2019, 165, 54–66. [Google Scholar] [CrossRef] [PubMed]
- Moore, S.D.; Kirkman, W.; Richards, G.I.; Stephen, P.R. The Cryptophlebia leucotreta Granulovirus—10 Years of Commercial Field Use. Viruses 2015, 7, 1284–1312. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pincus, D.H.; Orenga, S.; Chatellier, S. Yeast identification—Past, present, and future methods. Med. Mycol. 2007, 45, 97–121. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Opoku-Debrah, J.K. Studies on Existing and New Isolates of Cryptophlebia leucotreta Granulovirus (CrleGV) on Thaumatotibia leucotreta Populations from a Range of Geographic Regions in South Africa. Ph.D. Thesis, Rhodes University, Makhanda, South Africa, 2012. [Google Scholar]
- Moore, S.D.; Richards, G.; Chambers, C.; Hendry, D.A. An Improved Larval Diet for Commercial Mass Rearing of the False Codling Moth, Thaumatotibia leucotreta (Meyrick) (Lepidoptera: Tortricidae). Afr. Entomol. 2014, 22, 216–219. [Google Scholar] [CrossRef]
- Ljunggren, J.; Borrero-Echeverry, F.; Chakraborty, A.; Lindblom, T.U.T.; Hedenström, E.; Karlsson, M.; Witzgall, P.; Bengtsson, M. Yeast Volatomes Differentially Affect Larval Feeding in an Insect Herbivore. Appl. Environ. Microbiol. 2019, 85, e01761-19. [Google Scholar] [CrossRef] [Green Version]
- Love, C.N.; Hill, M.; Moore, S.D. Thaumatotibia leucotreta and the Navel orange: Ovipositional preferences and host susceptibility. J. Appl. Entomol. 2014, 138, 600–611. [Google Scholar] [CrossRef]
- Hajek, A.E.; McManus, M.L.; Delalibera, I. A review of introductions of pathogens and nematodes for classical biological control of insects and mites. Biol. Control 2007, 41, 1–13. [Google Scholar] [CrossRef]
- Davis, T.S.; Crippen, T.L.; Hofstetter, R.W.; Tomberlin, J.K. Microbial Volatile Emissions as Insect Semiochemicals. J. Chem. Ecol. 2013, 39, 840–859. [Google Scholar] [CrossRef]
- Heras-Vazquez, F.J.L.; Mingorance-Cazorla, L.; Clemente-Jimenez, J.M.; Rodriguez-Vico, F. Identification of yeast species from orange fruit and juice by RFLP and sequence analysis of the 5.8S rRNA gene and the two internal transcribed spacers. FEMS Yeast Res. 2003, 3, 3–9. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vadkertiová, R.; Molnárová, J.; Vránová, D.; Sláviková, E. Yeasts and yeast-like organisms associated with fruits and blossoms of different fruit trees. Can. J. Microbiol. 2012, 58, 1344–1352. [Google Scholar] [CrossRef] [PubMed]
- Hidalgo, C.; Mateo, E.; Mas, A.; Torija, M. Identification of yeast and acetic acid bacteria isolated from the fermentation and acetification of persimmon (Diospyros kaki). Food Microbiol. 2012, 30, 98–104. [Google Scholar] [CrossRef] [PubMed]
- Ganter, P.F.; Morais, P.B.; Rosa, C.A. Yeasts in Cacti and Tropical Fruit. In Yeasts in Natural Ecosystems: Diversity; Buzzini, P., Lachance, M.-A., Yurkov, A., Eds.; Springer International Publishing: Cham, Switzerland, 2017; pp. 225–264. ISBN 978-3-319-62683-3. [Google Scholar]
- Bellamy, D.E.; Sisterson, M.S.; Walse, S.S. Quantifying Host Potentials: Indexing Postharvest Fresh Fruits for Spotted Wing Drosophila, Drosophila suzukii. PLoS ONE 2013, 8, e61227. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hamby, K.A.; Hernández, A.; Boundy-Mills, K.; Zalom, F.G. Associations of Yeasts with Spotted-Wing Drosophila (Drosophila suzukii; Diptera: Drosophilidae) in Cherries and Raspberries. Appl. Environ. Microbiol. 2012, 78, 4869–4873. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.C.; Bruck, D.J.; Curry, H.; Edwards, D.; Haviland, D.R.; Van Steenwyk, R.A.; Yorgey, B.M. The susceptibility of small fruits and cherries to the spotted-wing drosophila, Drosophila suzukii. Pest Manag. Sci. 2011, 67, 1358–1367. [Google Scholar] [CrossRef]
- Wang, X.-G.; Stewart, T.J.; Biondi, A.; Chavez, B.A.; Ingels, C.; Caprile, J.; Grant, J.A.; Walton, V.M.; Daane, K.M. Population dynamics and ecology of Drosophila suzukii in Central California. J. Pest Sci. 2016, 89, 701–712. [Google Scholar] [CrossRef]
- Grangeteau, C.; Yahou, F.; Everaerts, C.; Dupont, S.; Farine, J.-P.; Beney, L.; Ferveur, J.-F. Yeast quality in juvenile diet affects Drosophila melanogaster adult life traits. Sci. Rep. 2018, 8, 13070. [Google Scholar] [CrossRef]
- Lewis, M.T.; Hamby, K.A. Differential Impacts of Yeasts on Feeding Behavior and Development in Larval Drosophila suzukii (Diptera:Drosophilidae). Sci. Rep. 2019, 9, 13370. [Google Scholar] [CrossRef] [Green Version]
- Starmer, W.T.; Aberdeen, V. The Nutritional Importance of Pure and Mixed Cultures of Yeasts in the Development of Drosophila mulleri Larvae in Opuntia Tissues and Its Relationship to Host Plant Shifts. In Ecological and Evolutionary Genetics of Drosophila; Springer: New York, NY, USA, 1990; pp. 145–160. [Google Scholar]
- Geng, P.; Chen, S.; Hu, M.; Rizwan-Ul-Haq, M.; Lai, K.; Qu, F.; Zhang, Y. Combination of Kluyveromyces marxianus and sodium bicarbonate for controlling green mold of citrus fruit. Int. J. Food Microbiol. 2011, 151, 190–194. [Google Scholar] [CrossRef]
- Li, W.; Zhang, H.; Li, P.; Apaliya, M.T.; Yang, Q.; Peng, Y.; Zhang, X. Biocontrol of postharvest green mold of oranges by Hanseniaspora uvarum Y3 in combination with phosphatidylcholine. Biol. Control 2016, 103, 30–38. [Google Scholar] [CrossRef]
- Lima, J.R.; Gondim, D.M.F.; Oliveira, J.T.A.; Oliveira, F.S.A.; Goncalves, L.R.B.; Viana, F.M.P. Use of killer yeast in the management of postharvest papaya anthracnose. Postharvest Biol. Technol. 2013, 83, 58–64. [Google Scholar] [CrossRef]
- Ripley, L.B.; Hepburn, G.A.; Dick, J. Mass Breeding of False Codling-Moth, Argyroploce leucotreta Meyr., in Artificial Media; Science Bulletin of the Department of Agriculture and Forestry of the Union of South Africa: Pretoria, South Africa, 1939. [Google Scholar]
- Theron, P.P.A. Studies in Verband met die Verskaffing van Gashere vir die Massateelt van Appelmotparasiete; Staatsdrukker: Pretoria, South Africa, 1948. [Google Scholar]
- Malan, A.P.; Von Diest, J.; Moore, S.D.; Addison, P. Control Options for False Codling Moth, Thaumatotibia leucotreta (Lepidoptera: Tortricidae), in South Africa, with Emphasis on the Potential Use of Entomopathogenic Nematodes and Fungi. Afr. Entomol. 2018, 26, 14–29. [Google Scholar] [CrossRef]
- Gregg, P.C.; Del Socorro, A.P.; Landolt, P.J. Advances in Attract-and-Kill for Agricultural Pests: Beyond Pheromones. Annu. Rev. Entomol. 2018, 63, 453–470. [Google Scholar] [CrossRef] [PubMed]
- El-Sayed, A.M.; Suckling, D.; Byers, J.A.; Jang, E.B.; Wearing, C.H. Potential of “Lure and Kill” in Long-Term Pest Management and Eradication of Invasive Species. J. Econ. Entomol. 2009, 102, 815–835. [Google Scholar] [CrossRef] [PubMed]
- Anagnostou, C.; Dorsch, M.; Rohlfs, M. Influence of dietary yeasts on Drosophila melanogaster life-history traits. Entomol. Exp. Appl. 2010, 136, 1–11. [Google Scholar] [CrossRef]
- Piper, A.M.; Farnier, K.; Linder, T.; Speight, R.; Cunningham, J.P. Two Gut-Associated Yeasts in a Tephritid Fruit Fly have Contrasting Effects on Adult Attraction and Larval Survival. J. Chem. Ecol. 2017, 43, 891–901. [Google Scholar] [CrossRef] [PubMed]
- Tasin, M.; Herrera, S.L.; Knight, A.L.; Barros-Parada, W.; Fuentes-Contreras, E.; Pertot, I. Volatiles of Grape Inoculated with Microorganisms: Modulation of Grapevine Moth Oviposition and Field Attraction. Microb. Ecol. 2018, 76, 751–761. [Google Scholar] [CrossRef] [Green Version]
- Hamby, K.A.; Becher, P.G. Current knowledge of interactions between Drosophila suzukii and microbes, and their potential utility for pest management. J. Pest Sci. 2016, 89, 621–630. [Google Scholar] [CrossRef]
- Knudsen, J.T.; Tollsten, L.; Bergström, L.G. Floral scents—A checklist of volatile compounds isolated by head-space techniques. Phytochemistry 1993, 33, 253–280. [Google Scholar] [CrossRef]
- Bruce, T.J.A.; Wadhams, L.J.; Woodcock, C.M. Insect host location: A volatile situation. Trends Plant Sci. 2005, 10, 269–274. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Amplicon | Yeast Species | Accession Number | Identity |
|---|---|---|---|---|
| Addo “One” | ITS | Meyerozyma guilliermondii | MN537824.1 | 100% |
| D1/D2 | Meyerozyma guilliermondii | EU285513.1 | 100% | |
| Addo “Two” | ITS | Hanseniaspora uvarum | MN371907.1 | 98.28% |
| D1/D2 | Hanseniaspora uvarum | AY305681.1 | 99.64% | |
| Addo “Three” | ITS | Clavispora lusitaniae | KP765042.1 | 99.52% |
| D1/D2 | Clavispora lusitaniae | MG871742.1 | 99.82% | |
| Stellenbosch | ITS | Kluyveromyces marxianus | KX376261.1 | 99.70% |
| D1/D2 | Kluyveromyces marxianus | CP009307.2 | 100% | |
| Common “One” | ITS | Pichia kluyveri | KM982973.1 | 99.75% |
| D1/D2 | Pichia kluyveri | MN464128.1 | 99.64% | |
| Common “Two” | ITS | Pichia kudriavzevii | LC389027.1 | 100% |
| D1/D2 | Pichia kudriavzevii | MF461295.1 | 100% |
| Treatments | Total Pupated | Pupated | Percentage before 25 d | p-Value | |
|---|---|---|---|---|---|
| Before 25 d | After 25 d | ||||
| Control | 41 | 15 | 26 | 37% | |
| M. guilliermondi | 59 | 35 | 24 | 59% | 0.0414 * |
| P. kluyveri | 78 | 58 | 20 | 74% | 0.0001 ** |
| P. kudriavzevii | 58 | 35 | 23 | 60% | 0.0253 * |
| H. uvarum | 63 | 40 | 23 | 63% | 0.0092 ** |
| C. lusitaniae | 42 | 30 | 12 | 71% | 0.0020 ** |
| K. marxianus | 58 | 38 | 20 | 66% | 0.0075 ** |
| S. cerevisiae | 62 | 42 | 20 | 68% | 0.0024 ** |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
van der Merwe, M.; Jukes, M.D.; Knox, C.; Moore, S.D.; Hill, M.P. Mutualism between Gut-Borne Yeasts and Their Host, Thaumatotibia leucotreta, and Potential Usefulness in Pest Management. Insects 2022, 13, 243. https://doi.org/10.3390/insects13030243
van der Merwe M, Jukes MD, Knox C, Moore SD, Hill MP. Mutualism between Gut-Borne Yeasts and Their Host, Thaumatotibia leucotreta, and Potential Usefulness in Pest Management. Insects. 2022; 13(3):243. https://doi.org/10.3390/insects13030243
Chicago/Turabian Stylevan der Merwe, Marcel, Michael D. Jukes, Caroline Knox, Sean D. Moore, and Martin P. Hill. 2022. "Mutualism between Gut-Borne Yeasts and Their Host, Thaumatotibia leucotreta, and Potential Usefulness in Pest Management" Insects 13, no. 3: 243. https://doi.org/10.3390/insects13030243