
Seasonal Prevalence of the Invasive Longhorn Beetle Aromia bungii in Osaka Prefecture, Japan


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
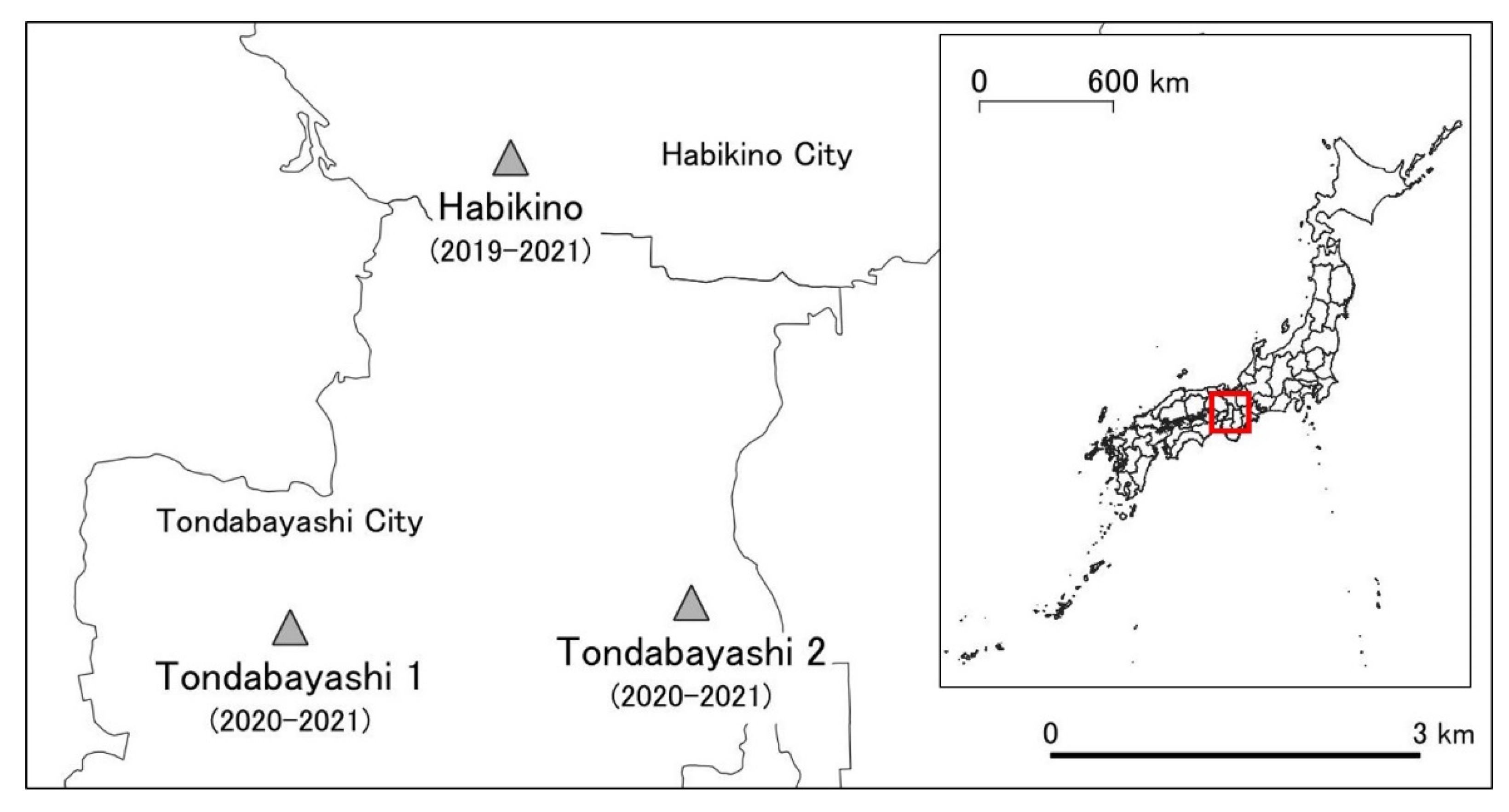
2.1. Survey Years and Study Sites
- Research Institute of Environment, Agriculture and Fisheries, Habikino City, Osaka Prefecture (34°32′8″ N, 135°35′53″ E) (hereafter referred to as Habikino) from 2019 to 2021.
- Sidewalk along Route 309 in Tondabayashi City (34°30′22″ N, 135°34′33″ E) (hereafter referred to as Tondabayashi 1) from 2020 to 2021.
- Riverside road of Ishikawa River in Tondabayashi City (34°30′9″ N, 135°36′50″ E) (hereafter referred to as Tondabayashi 2) from 2020 to 2021 (Figure 1).
2.2. Determination of the Number of Adults Sighted on Trees in the Field
2.3. Determination of Emergence Dates of Emerged Adults from Infested Trees in the Field
2.4. Statistical Analyses
3. Results
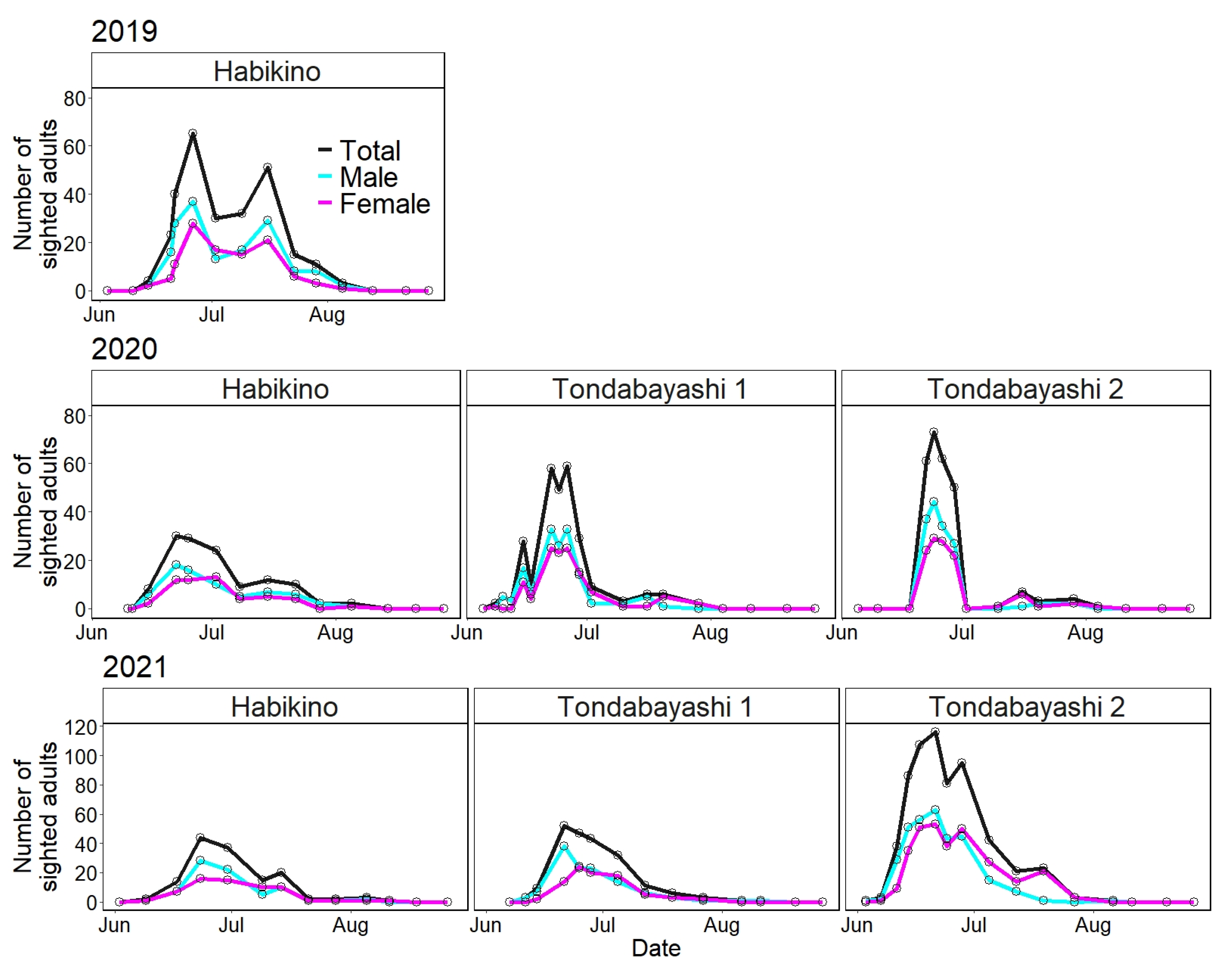
3.1. Seasonal Prevalence Based on the Number of Sighted Adults in the Field
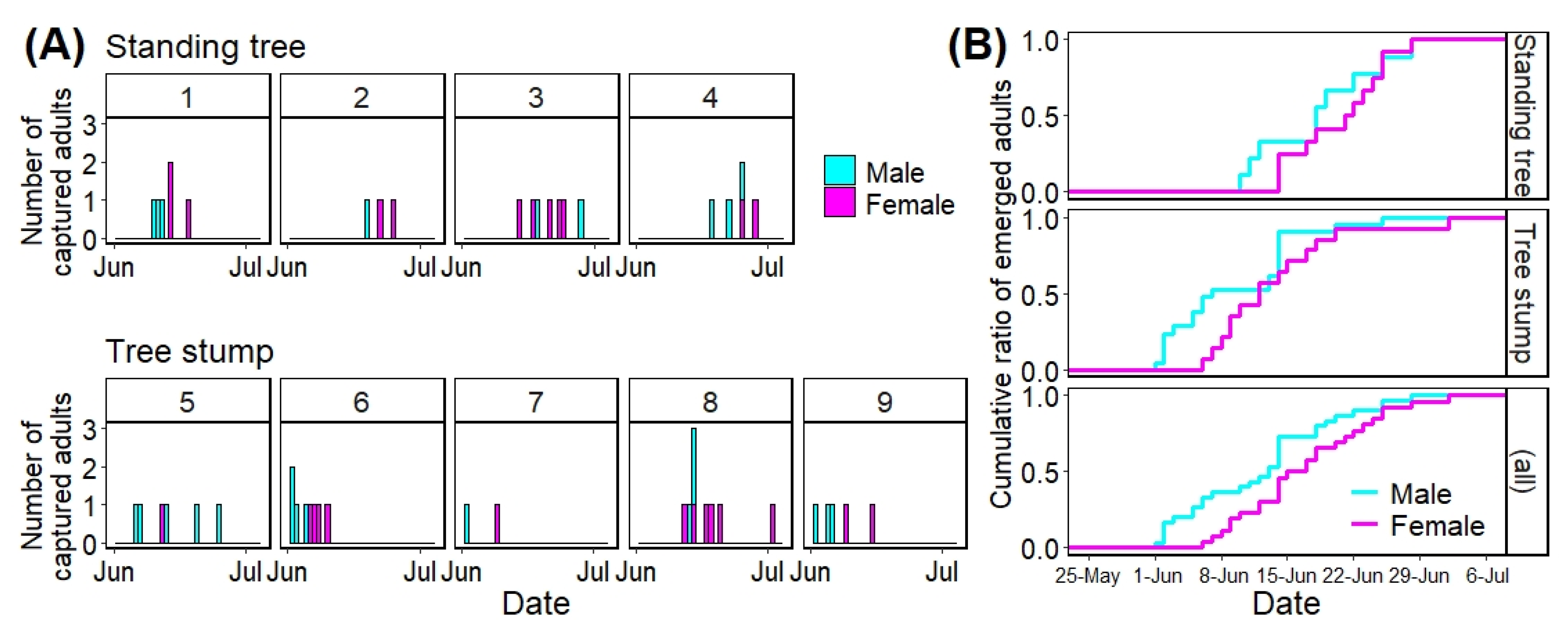
3.2. Adult Emergence from Infested Trees
3.2.1. Dates of Adult Emergence from Infested Trees
3.2.2. Factors Affecting Adult Emergence Day
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Study Site | Tree Species | 2019 | 2020 | 2021 | ||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Start | During Survey | Start | During Survey | Start | During Survey | |||||
| Cut | No Access | Cut | No Access | Cut | No Access | |||||
| Habikino | Ornamental cherry (Cerasus spp.) | 138 | 0 | 0 | 87 | 0 | 0 | 91 | 0 | 0 |
| Prunus mume | 27 | 0 | 0 | 27 | 0 | 0 | 44 | 0 | 0 | |
| Prunus cerasifera var. atropurpurea | 16 | 0 | 0 | 10 | 0 | 0 | 10 | 0 | 0 | |
| Tondabayashi 1 | Ornamental cherry (Cerasus spp.) | ― | ― | ― | 244 | 10 | 0 | 231 | 11 | 0 |
| Tondabayashi 2 | Ornamental cherry (Cerasus spp.) | ― | ― | ― | 134 | 5 | 36 | 249 | 0 | 0 |
Appendix B
| Tree Type | Tree No. | Random Intercept Value |
|---|---|---|
| Standing tree | 1 | −5.424 |
| 2 | 1.387 | |
| 3 | 0.351 | |
| 4 | 3.686 | |
| Tree stump | 5 | 4.552 |
| 6 | −4.719 | |
| 7 | −3.202 | |
| 8 | 5.513 | |
| 9 | −2.143 |
Appendix C
| Sex | Tree Type | Mean ± SD | Min | Max | Median |
|---|---|---|---|---|---|
| Male | Standing tree | 22.9 ± 2.2 | 18.7 | 25.1 | 23.7 |
| Tree stump | 22.4 ± 1.9 | 18.8 | 26.3 | 22.2 | |
| Female | Standing tree | 24.0 ± 1.5 | 20.8 | 26.5 | 23.9 |
| Tree stump | 23.8 ± 1.7 | 20.0 | 25.7 | 24.3 |
References
- EPPO. EPPO datasheet on pests recommended for regulation Aromia bungii. EPPO Bull. 2015, 45, 4–8. [Google Scholar] [CrossRef]
- EFSA. Aromia bungii Pest Report to Support Ranking of EU Candidate Priority Pests; Pest Report EN-1639; EFSA: Parma, Italy, 2019; pp. 1–33. [CrossRef]
- Russo, E.; Nugnes, F.; Vicinanza, F.; Garonna, A.P.; Bernardo, U. Biological and molecular characterization of Aromia bungii (Faldermann, 1835) (Coleoptera: Cerambycidae), an emerging pest of stone fruits in Europe. Sci. Rep. 2020, 10, 7112. [Google Scholar] [CrossRef] [PubMed]
- Iwata, R. Aromia bungii (Coleoptera: Cerambycidae): Taxonomy, distribution, biology and eradication. For. Pests 2018, 67, 7–34. (In Japanese) [Google Scholar]
- Tamura, S.; Shoda-Kagaya, E. Process of distribution expansion of the red-necked longicorn beetles Aromia bungii in Japan. Shinrin Kagaku 2020, 89, 21–25. (In Japanese) [Google Scholar] [CrossRef]
- Uechi, N. Fruit tree damage by the red-necked longhorn beetle Aromia bungii. Shinrin Kagaku 2020, 89, 14–17. (In Japanese) [Google Scholar] [CrossRef]
- Yamamoto, Y. Invasion and control strategy of the red-necked longhorn beetle Aromia bungii. J. Pestic. Sci. 2020, 45, 127–133. (In Japanese) [Google Scholar] [CrossRef]
- Nakano, A.; Watanabe, T. Damage caused by the red-necked longhorn beetle, Aromia bungii in peach orchards in Tokushima Prefecture and trial control method. Plant Prot. 2017, 71, 723–728. (In Japanese) [Google Scholar]
- Kano, M.; Nonaka, T.; Kiriyama, S.; Iwata, R. Aromia bungii (Coleoptera: Cerambycidae), an invasive cerambycid, found at Soka, Saitama Pref., Japan, infesting cherry trees, Cerasus × yedoensis ‘Somei-yoshino’. For. Pests 2014, 63, 101–105. (In Japanese) [Google Scholar]
- Enda, N. Life history of the Japanese pine sawyer Monochamus alternatus. For. Pests 1976, 12, 182–185. Available online: https://forest-pests.sakura.ne.jp/backnumber/files/vol25/25-12.pdf (accessed on 31 January 2022). (In Japanese).
- Iba, M.; Inoue, S.; Kikuchi, M. Ecological studies on the yellow-spotted longicorn beetle, Psacothea hilaris Pascoe Ι. The local difference in the seasonal prevalence of the adult insect. J. Sericult. Sci. Jpn. 1976, 45, 156–160. (In Japanese) [Google Scholar] [CrossRef]
- Flaherty, L.; Quiring, D.; Pureswaran, D.; Sweeney, J. Evaluating seasonal variation in bottom-up and top-down forces and their impact on an exotic wood borer, Tetropium fuscum (Coleoptera: Cerambycidae). Environ. Entomol. 2013, 42, 957–966. [Google Scholar] [CrossRef] [PubMed]
- Akessé, E.N.; Ouali-N’Goran, S.-W.M.; N’Dépo, O.R.; Tano, D.K.C. Population fluctuation of Diastocera trifasciata (Fabricius, 1775) (Coleoptera: Cerambycidae), cashew branches girdler in the Brobo locality (Central Côte d’Ivoire). J. Entomol. Zool. Stud. 2018, 6, 1064–1070. Available online: https://www.entomoljournal.com/archives/2018/vol6issue5/PartR/6-5-59-552.pdf (accessed on 31 January 2022).
- Hanks, L.M.; Reagel, P.F.; Mitchell, R.F.; Wong, J.C.H.; Meier, L.R.; Silliman, C.A.; Graham, E.E.; Striman, B.L.; Robinson, K.P.; Mongold-Diers, J.A.; et al. Seasonal phenology of the cerambycid beetles of east central Illinois. Ann. Entomol. Soc. Am. 2014, 107, 211–226. [Google Scholar] [CrossRef] [Green Version]
- Handley, K.; Hough-Goldstein, J.; Hanks, L.M.; Millar, J.G.; D’amico, V. Species richness and phenology of cerambycid beetles in urban forest fragments of northern Delaware. Ann. Entomol. Soc. Am. 2015, 108, 251–262. [Google Scholar] [CrossRef]
- Lyons-Yerion, C.D.; Barbour, J.D.; Mongold-Diers, J.A.; Williams, C.J.; Cook, S.P. Identification of a male-produced volatile pheromone for Phymatodes dimidiatus (Coleoptera: Cerambycidae) and seasonal flight phenology of four Phymatodes species endemic to the North American intermountain west. Environ. Entomol. 2020, 49, 1077–1087. [Google Scholar] [CrossRef] [PubMed]
- Shibata, E. Seasonal fluctuation and spatial pattern of the adult population of the Japanese pine sawyer, Monochamus alternatus Hope (Coleoptera: Cerambycidae), in young pine forests. Appl. Entomol. Zool. 1981, 16, 306–309. [Google Scholar] [CrossRef]
- Keszthelyi, S. Diversity and seasonal patterns of longhorn beetles (Coleoptera: Cerambycidae) in the Zselic region, Hungary. North-West. J. Zool. 2015, 11, 62–69. Available online: http://biozoojournals.ro/nwjz/content/v11n1/nwjz_141105_Keszthelyi.pdf (accessed on 31 January 2022).
- Xu, T.; Yasui, H.; Teale, S.A.; Fujiwara-Tsujii, N.; Wickham, J.D.; Fukaya, M.; Hansen, L.; Kiriyama, S.; Hao, D.; Nakano, A.; et al. Identification of male-produced sex-aggregation pheromone for a highly invasive cerambycid beetle, Aromia bungii. Sci. Rep. 2017, 7, 7330. [Google Scholar] [CrossRef]
- Germinara, G.S.; Pistillo, M.; Griffo, R.; Garonna, A.P.; Di Palma, A. Electroantennographic responses of Aromia bungii (Faldermann, 1835) (Coleoptera, Cerambycidae) to a range of volatile compounds. Insects 2019, 10, 274. [Google Scholar] [CrossRef] [Green Version]
- Yasui, H.; Fujiwara-Tsujii, N.; Kugimiya, S.; Haruyama, N. Extension of sustained pheromone release for monitoring an emerging invader, red-necked longicorn beetle Aromia bungii (Coleoptera: Cerambycidae). Appl. Entomol. Zool. 2021, 56, 291–297. [Google Scholar] [CrossRef]
- Esaki, K. Seasonal prevalence of the Japanese pine sawyer, Monochamus alternatus. Bull. Ishikawa. Agr. For. Res. Cen. For. Exp. Stn. 2013, 34, 47. Available online: https://www.pref.ishikawa.lg.jp/ringyo/publish/kenkyuuhoukoku/documents/34-11_kamikirihassei.pdf (accessed on 31 January 2022). (In Japanese).
- Han, J.-H.; Yoon, C.; Shin, S.-C.; Kim, G.-H. Seasonal occurrence and morphological measurements of pine sawyer, Monochamus saltuarius adults (Coleoptera: Cerambycidae). J. Asia-Pac. Entomol. 2007, 10, 63–67. [Google Scholar] [CrossRef]
- Maier, C.T. Emergence, trapping, and seasonal abundance of adult Cerambycidae (Coleoptera) associated with Cupressaceae in Connecticut. J. Econ. Entomol. 2008, 101, 430–437. [Google Scholar] [CrossRef] [PubMed]
- Urano, T.; Shoda-Kagaya, E. Longevity and lifetime fecundity of rearing adults of Aromia bungii (Coleoptera: Cerambycidae). Kanto J. For. Res. 2017, 68, 25–28. (In Japanese) [Google Scholar]
- Urano, T. Reproductive biology of Aromia bungii (Faldermann) (Coleoptrra: Cerambycidae) adults emerged from infested trunks of Cerasus × yedoensis ‘Somei-Yoshino’. For. Pests 2018, 67, 230–236. (In Japanese) [Google Scholar]
- Gotoh, T.; Shishiuchi, J.; Sugita, H. Emergence and oviposition of the alder longicorn beetle, Cagosima sanguinolenta Thomson (Coleoptera, Cerambycidae), and its damage in a young stand of Betula maximowicziana Regel (Betulaceae). Tohoku J. For. Sci. 2007, 12, 291–297. (In Japanese) [Google Scholar] [CrossRef]
- Yamamoto, Y.; Uehara, K.; Ishikawa, Y.; Tsuyoshi, Y. Survey on the actual damage by the invasive red-necked longhorn beetle (Aromia bungii) harmful to trees such as the Japanese flowering cherries. A. R. Pro. Nat. Found. Jpn. 2019, 8, 24–35. (In Japanese) [Google Scholar] [CrossRef]
- R: A Language and Environment for Statistical Computing. Available online: https://www.R-project.org/ (accessed on 31 January 2022).
- Akaike, H. A new look at the statistical model identification. IEEE Trans. Autom. Contr. 1974, 19, 716–723. [Google Scholar] [CrossRef]
- Funaki, Y. The emergency approach to pesticide registration for the red-necked longhorn beetle, Aromia bungii. Plant Prot. 2019, 73, 413–419. (In Japanese) [Google Scholar]
- Nakano, A. Damage situations and control measures for the red-necked longhorn beetle Aromia bungii in Tokushima Prefecture. Noyaku Jidai 2018, 199, 30–34. Available online: https://www.nippon-soda.co.jp/nougyo/pdf/no199/199_030.pdf (accessed on 31 January 2022). (In Japanese).
- Sano, N.; Fujishita, A. Methods of forecasting the occurrence and seasonal prevalence of adult emergence of Japanese pine sawyer (Monochamus alternatus) in Shizuoka Prefecture. Bull. Shizuoka. For. Tec. Cen. 1996, 24, 9–13. Available online: https://agriknowledge.affrc.go.jp/RN/2030550422.pdf (accessed on 31 January 2022).
- Matsuura, T.; Aiura, H. Prediction of damage dynamics by pine wilt disease reflected recent meteorological data in Toyama prefecture: Vertical distribution of the damage and emergence period of Monochamus alternatus adult. Bull. Toyama For. Res. Inst. 2018, 10, 24–31. Available online: https://taffrc.pref.toyama.jp/nsgc/shinrin/webfile/t1_b6faf7efa839c2dacd83250ca174b27c.pdf (accessed on 31 January 2022).
- Ohsawa, M.; Iijima, H. Adult emergence of Monochamus alternatus, the insect vector of pine wilt disease, in Yamanashi Prefecture. Bull. Yamanashi. For. Res. Inst. 2021, 40, 7–10. Available online: https://www.pref.yamanashi.jp/shinsouken/report/documents/40_1_osawaiijima.pdf (accessed on 31 January 2022).
- Keena, M.A.; Moore, P.M. Effects of temperature on Anoplophora glabripennis (Coleoptera: Cerambycidae) larvae and pupae. Environ. Entomol. 2010, 39, 1323–1335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Esaki, K. Spatio-temporal distribution of adult emergence holes in trees infested by the Japanese pine sawyer Monochamus alternatus (Coleoptera: Cerambycidae) and its relation to inside tree temperature. Bull. Ishikawa Agr. For. Res. Cen. For. Exp. Stn. 2021, 52, 6–11. Available online: https://www.pref.ishikawa.lg.jp/ringyo/publish/kenkyuuhoukoku/documents/52-2_distribution-temperature.pdf (accessed on 31 January 2022). (In Japanese).
- Akbulut, S.; Keten, A.; Baysal, I.; Yuksel, B. The effect of log seasonality on the reproductive potential of Monochamus galloprovincialis Olivier (Coleoptera: Cerambycidae) reared in black pine logs under laboratory conditions. Turk. J. Agric. For. 2007, 31, 413–422. [Google Scholar] [CrossRef]
- Taniwaki, T. Temperature and humidity conditions in the xylem of the damaged tree by pine wilt disease placed at bare and tree-shaded sites. J. Jpn. For. Soc. 2008, 90, 253–256. (In Japanese) [Google Scholar] [CrossRef] [Green Version]




| Survey Year | Study Site | Survey Date and Number of Sighted Adults by Sex (Male vs. Female) | Total Number of Sighted Adults | |||||
|---|---|---|---|---|---|---|---|---|
| Initial Sighting | Highest Number Sighting | Last Sighting | ||||||
| 2019 | Habikino | 14 Jun | 4 (2|2) | 26 Jun | 65 (37|28) | 5 Aug | 3 (2|1) | 274 (160|109|5) |
| 2020 | Habikino | 15 Jun | 8 (6|2) | 22 Jun | 30 (18|12) | 5 Aug | 2 (1|1) | 126 (71|53|2) |
| Tondabayashi 1 | 8 Jun | 2 (1|1) | 26 Jun | 59 (33|25|1) | 29 Jul | 2 (0|2) | 269 (148|120|1) | |
| Tondabayashi 2 | 22 Jun | 61 (37|24) | 24 Jun | 73 (44|29) | 4 Aug | 1 (0|1) | 262 (147|114|1) | |
| 2021 | Habikino | 9 Jun | 2 (1|1) | 23 Jun | 44 (28|16) | 11 Aug | 1 (0|1) | 140 (77|63) |
| Tondabayashi 1 | 11 Jun | 3 (3|0) | 21 Jun | 52 (38|14) | 11 Aug | 1 (1|0) | 208 (120|88) | |
| Tondabayashi 2 | 3 Jun | 1 (1|0) | 21 Jun | 116 (63|53) | 6 Aug | 1 (1|0) | 617 (314|302|1) | |
| Tree Type | Tree No. | First Capture | Last Capture | Emergence Period (Day) | Total Number of Adults | |||
|---|---|---|---|---|---|---|---|---|
| Date | Elapsed Day | Date | Elapsed Day | Emerged | Not Emerged | |||
| Standing tree | 1 | 10 Jun | Day 10 | 18 Jun | Day 18 | 9 | 6 (3|3) | 3 |
| 2 | 19 Jun | Day 19 | 25 Jun | Day 25 | 7 | 3 (1|2) | 1 | |
| 3 | 14 Jun | Day 14 | 28 Jun | Day 28 | 15 | 7 (2|5) | 2 | |
| 4 | 18 Jun | Day 18 | 28 Jun | Day 28 | 11 | 5 (3|2) | 1 | |
| Tree stump | 5 | 6 Jun | Day 6 | 25 Jun | Day 25 | 20 | 10 (9|1) | 8 |
| 6 | 1 Jun | Day 1 | 10 Jun | Day 10 | 10 | 9 (5|4) | 5 | |
| 7 | 2 Jun | Day 2 | 9 Jun | Day 9 | 8 | 2 (1|1) | 1 | |
| 8 | 12 Jun | Day 12 | 2 Jul | Day 32 | 21 | 9 (3|6) | 2 | |
| 9 | 2 Jun | Day 2 | 15 Jun | Day 15 | 14 | 5 (3|2) | 1 | |
| Rank | AIC | Conditional R2 | Regression Coefficient | |||
|---|---|---|---|---|---|---|
| Forewing Length | Sex (Female = 0) | Tree Type (Standing Tree = 0) | Intercept | |||
| 1 | 348.7 | 0.706 | – | −3.684 | −9.169 | 21.365 |
| 2 | 350.5 | 0.709 | 0.154 | −3.542 | −9.145 | 17.670 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yamamoto, Y.; Kaneko, S. Seasonal Prevalence of the Invasive Longhorn Beetle Aromia bungii in Osaka Prefecture, Japan. Insects 2022, 13, 222. https://doi.org/10.3390/insects13030222
Yamamoto Y, Kaneko S. Seasonal Prevalence of the Invasive Longhorn Beetle Aromia bungii in Osaka Prefecture, Japan. Insects. 2022; 13(3):222. https://doi.org/10.3390/insects13030222
Chicago/Turabian StyleYamamoto, Yuichi, and Shuji Kaneko. 2022. "Seasonal Prevalence of the Invasive Longhorn Beetle Aromia bungii in Osaka Prefecture, Japan" Insects 13, no. 3: 222. https://doi.org/10.3390/insects13030222