
Compatibility of Insecticides with Rice Resistance to Planthoppers as Influenced by the Timing and Frequency of Applications


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Herbivore Species
2.2. Plant Materials
2.3. Insecticides
2.4. Effects of Application Frequency on BPH and Host Plants
2.5. Effects of Application Timing on BPH and Host Plants
2.6. Data Analyses
3. Results
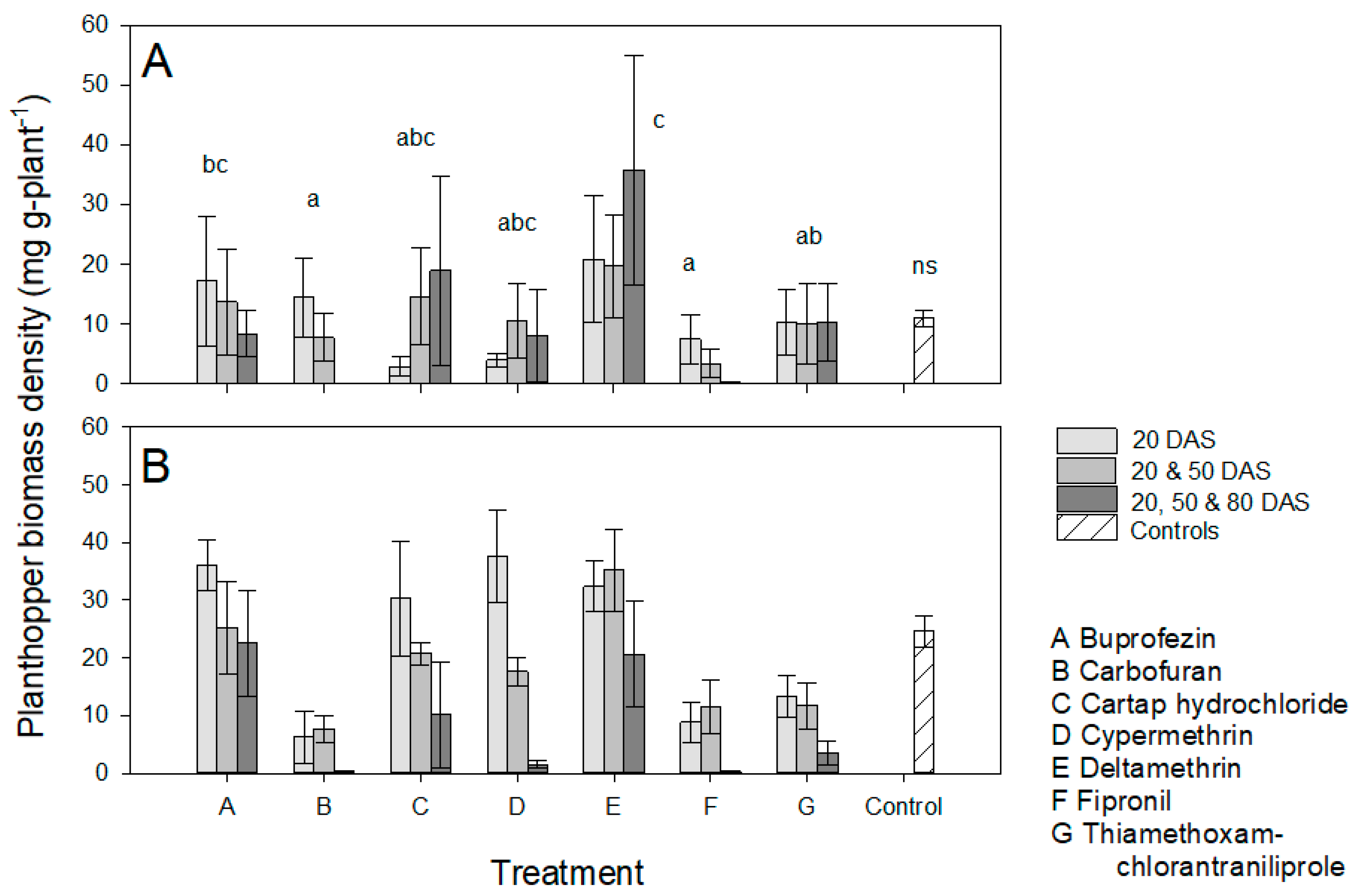
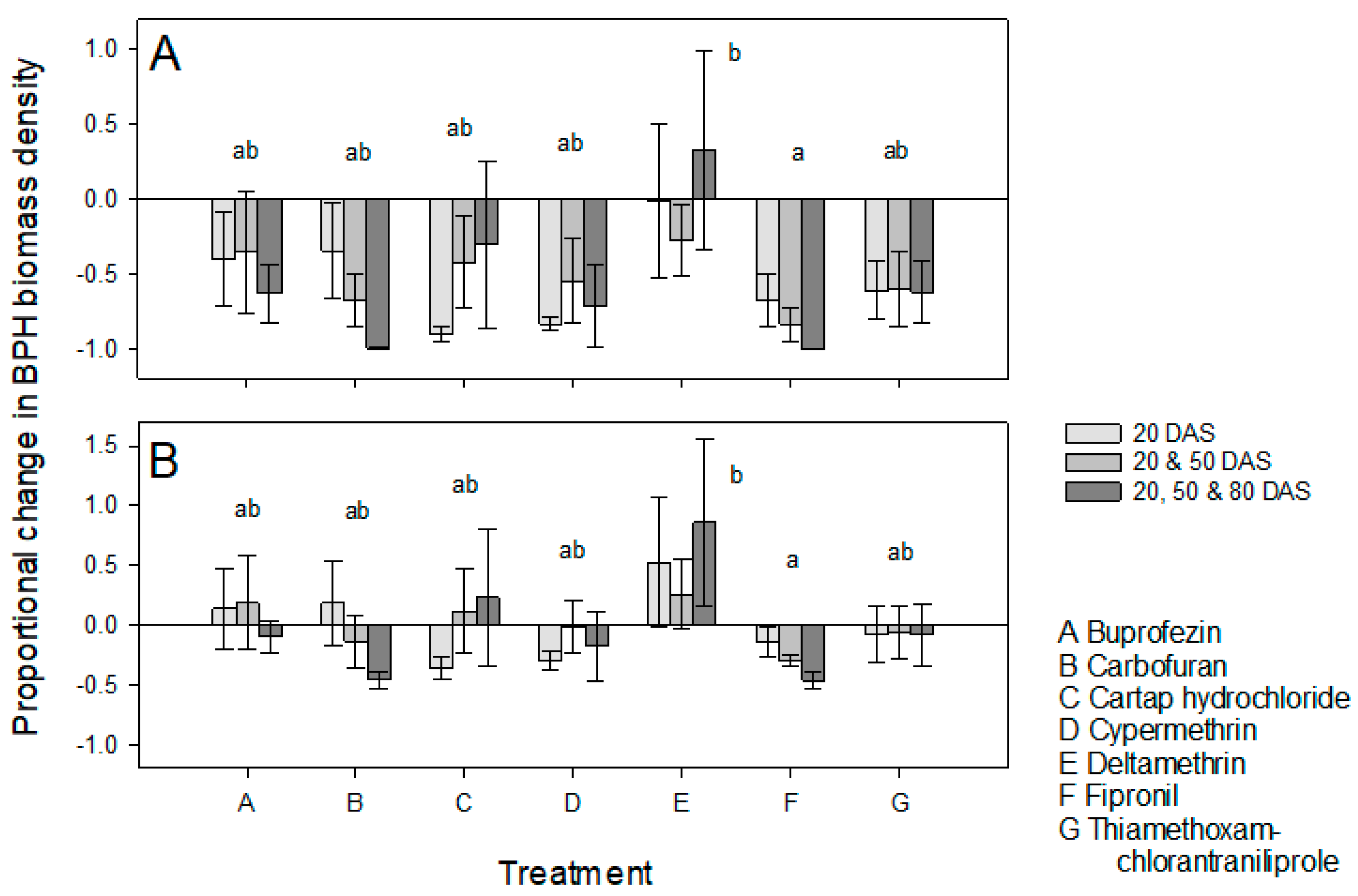
3.1. Effects of Host Resistance and the Frequency of Insecticide Applications on BPH Populations
3.2. Effects of Host Resistance and the Frequency of Insecticide Applications on Plant Growth
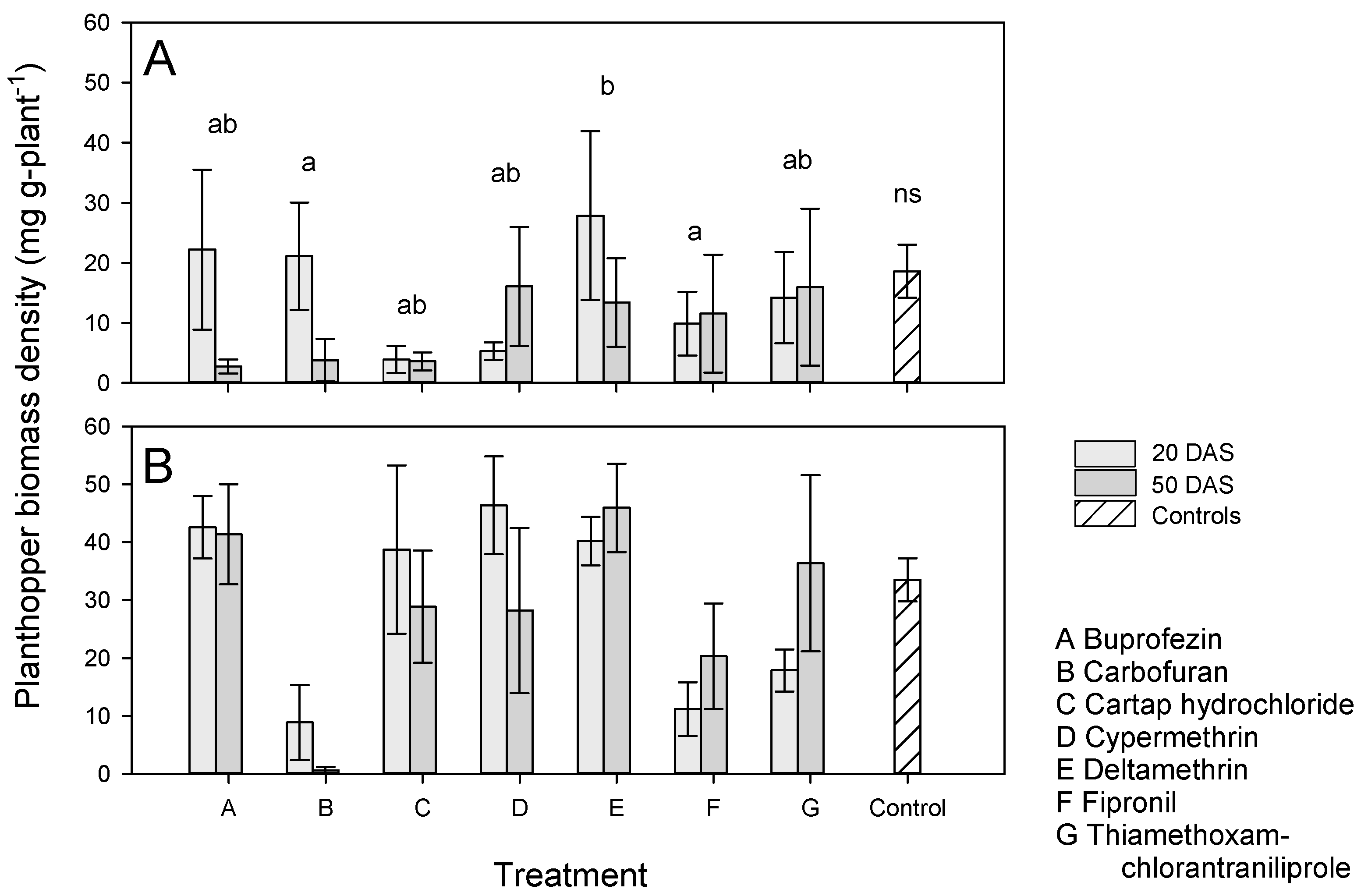
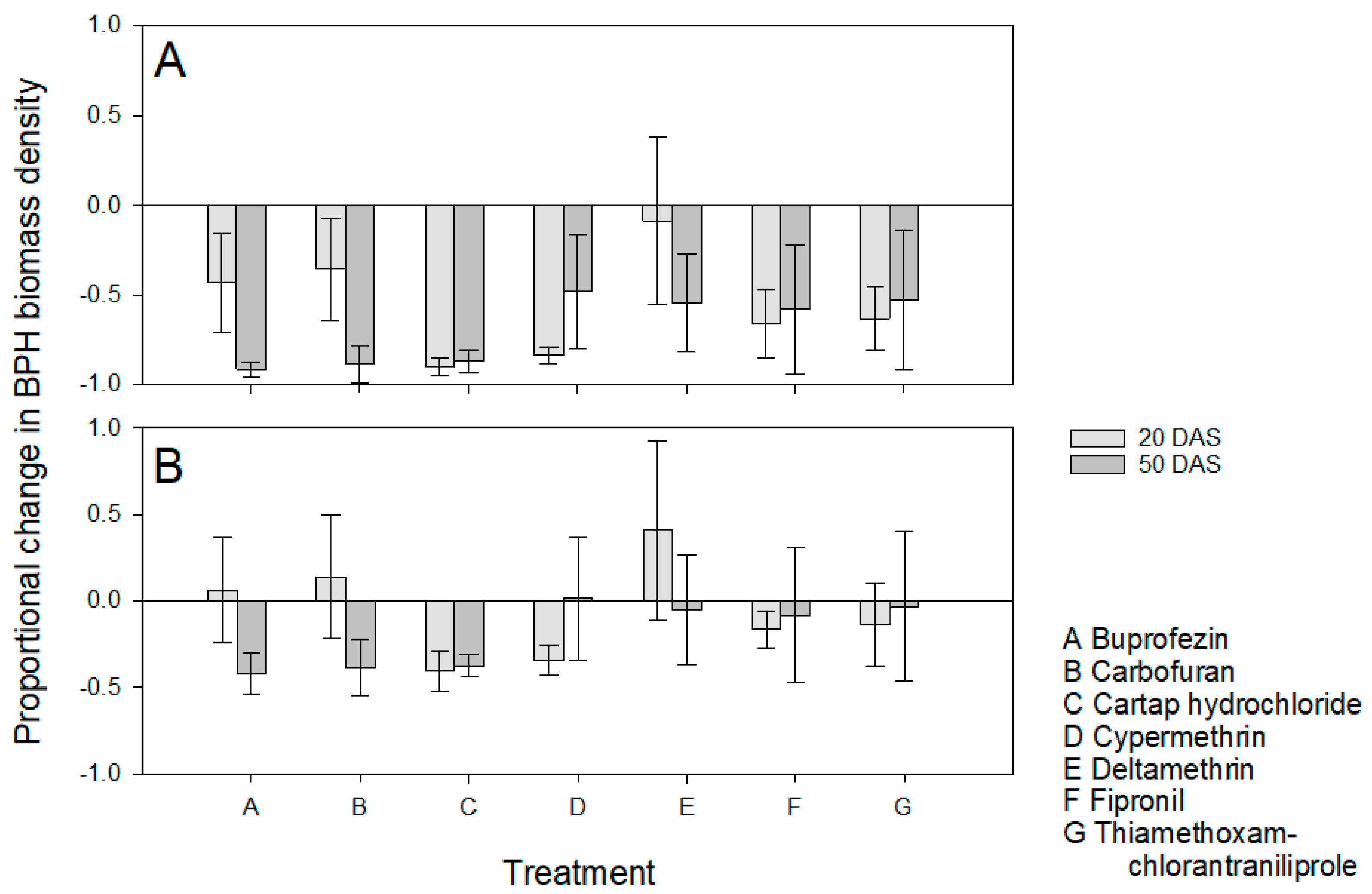
3.3. Effects of Application Time and Host Resistance on BPH Populations
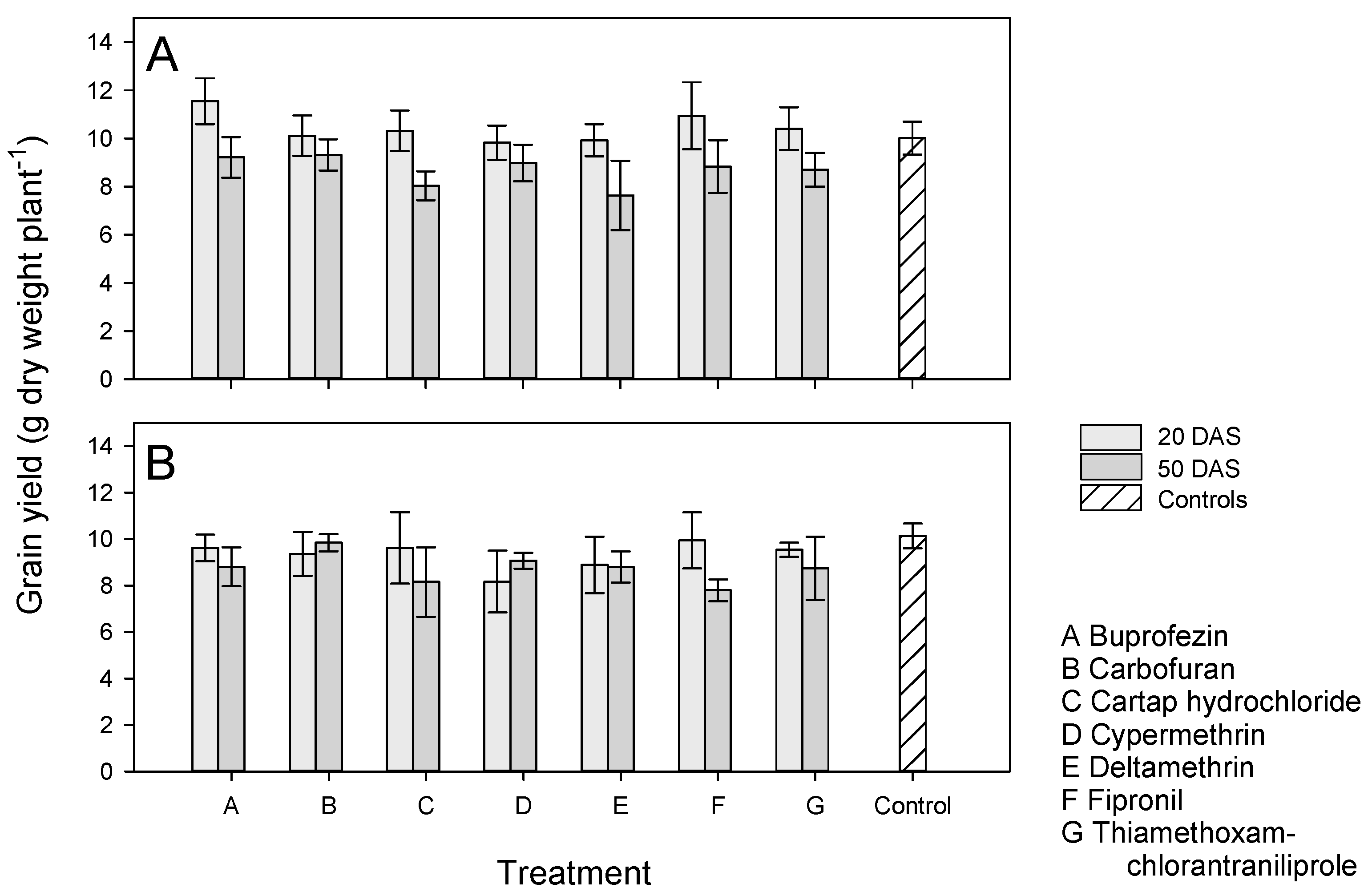
3.4. Effects of Application Time and Host Resistance on Plant Growth
4. Discussion
4.1. Potential Synergies
4.2. Potential Antagonisms
4.3. Phytotoxicity
4.4. Implications for BPH Management
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Muthayya, S.; Sugimoto, J.D.; Montgomery, S.; Maberly, G.F. An overview of global rice production, supply, trade, and consumption. Ann. N. Y. Acad. Sci. 2014, 1324, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Maclean, J.; Hardy, B.; Hettel, G. Rice Almanac; International Rice Research Institute: Los Baños, Philippines, 2013. [Google Scholar]
- Horgan, F.G. Integrated pest management for sustainable rice cultivation: A holistic approach. In Achieving Sustainable Cultivation of Rice: Cultivation, Pest and Disease Management; Sasaki, T., Ed.; Burleigh Dodds Scientific: Cambridge, UK, 2017; pp. 309–342. [Google Scholar]
- Bottrell, D.G.; Schoenly, K.G. Resurrecting the ghost of green revolutions past: The brown planthopper as a recurring threat to high-yielding rice production in tropical Asia. J. Asia-Pac. Entomol. 2012, 15, 122–140. [Google Scholar] [CrossRef]
- Heinrichs, E.A. Biology and Management of Rice Insects; International Rice Research Institute: Los Baños, Philippines, 1994. [Google Scholar]
- Pathak, M.D.; Khan, Z.R. Insect Pests of Rice; International Rice Research Institute: Los Baños, Philippines, 1994. [Google Scholar]
- Horgan, F.G. Integrating gene deployment and crop management for improved rice resistance to Asian planthoppers. Crop Prot. 2018, 110, 21–33. [Google Scholar] [CrossRef]
- Horgan, F.G.; Crisol, E. Hybrid rice and insect herbivores in Asia. Entomol. Exp. Appl. 2013, 148, 1–19. [Google Scholar] [CrossRef]
- Wu, J.; Ge, L.; Liu, F.; Song, Q.; Stanley, D. Pesticide-induced planthopper population resurgence in rice cropping systems. Annu. Rev. Entomol. 2020, 65, 409–429. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Xu, J.; Yuan, S.; Liu, J.; Jiang, Y.; Xu, J. Pesticide-induced susceptibility of rice to brown planthopper Nilaparvata Lugens. Entomol. Exp. Appl. 2001, 100, 119–126. [Google Scholar] [CrossRef] [Green Version]
- Heinrichs, E.A.; Reissig, W.H.; Valencia, S.; Chellliah, S. Rates and effect of resurgence-inducing insecticides on populations of Nilaparvata lugens (Homoptera: Delphacidae) and its Predators. Environ. Entomol. 1982, 11, 1269–1273. [Google Scholar] [CrossRef] [Green Version]
- Fujita, D.; Kohli, A.; Horgan, F.G. Rice resistance to planthoppers and leafhoppers. Crit. Rev. Plant Sci. 2013, 32, 162–191. [Google Scholar] [CrossRef]
- Horgan, F.G.; Almazan, M.L.P.; Vu, Q.; Ramal, A.F.; Bernal, C.C.; Yasui, H.; Fujita, D. Unanticipated benefits and potential ecological costs associated with pyramiding leafhopper resistance loci in rice. Crop Prot. 2019, 115, 47–58. [Google Scholar] [CrossRef]
- Widawsky, D.; Rozelle, S.; Jin, S.; Huang, J. Pesticide productivity, host-plant resistance and productivity in China. Agric. Econ. 1998, 19, 203–217. [Google Scholar]
- Wu, J.; Qiu, H.; Yang, G.; Liu, J.; Liu, G.; Wilkins, R.M. Effective duration of pesticide-induced susceptibility of rice to brown planthopper (Nilaparvata lugens Stål, Homoptera: Delphacidae), and physiological and biochemical changes in rice plants following pesticide application. Int. J. Pest Manag. 2004, 50, 55–62. [Google Scholar] [CrossRef]
- Wu, J.; Wang, A.; Xu, J.; Yang, G.; Qiu, H.; Li, D.H. Studies on stimulating effect of two selective insecticides on the number of egg laid by rice yellow borer, Tryporyza incertulas (Walker) and their effects on biochemistry of rice plants. Sci. Agric. Sin. 2003, 36, 1163–1170. [Google Scholar]
- Nanthakumar, M.; Jhansi Lakshmi, V.; Shashi Bhushan, V.; Balachandran, S.M.; Mohan, M. Decrease of rice plant resistance and induction of hormesis and carboxylesterase titre in brown planthopper, Nilaparvata lugens (Stål) by xenobiotics. Pestic. Biochem. Physiol. 2012, 102, 146–152. [Google Scholar] [CrossRef]
- Azzam, S.; Wang, F.; Wu, J.; Shen, J.; Wang, L.; Yang, G.; Guo, Y. Comparisons of stimulatory effects of a series of concentrations of four insecticides on reproduction in the rice brown planthopper Nilaparvata lugens Stål (Homoptera: Delphacidae). Int. J. Pest Manag. 2009, 55, 347–358. [Google Scholar] [CrossRef]
- Reissig, W.H.; Heinrichs, E.A.; Valencia, S.L. Insecticide-induced resurgence of the brown planthopper, Nilaparvata lugens, on rice varieties with different levels of resistance. Environ. Entomol. 1982, 11, 165–168. [Google Scholar] [CrossRef] [Green Version]
- Salim, M.; Heinrichs, E.A. Insecticide-induced changes in the levels of resistance of rice cultivars to the whitebacked planthopper Sogatella furcifera (Horváth) (Homoptera: Delphacidae). Crop Prot. 1987, 6, 28–32. [Google Scholar] [CrossRef]
- Wang, L.; Shen, J.; Ge, L.; Wu, J.; Yang, G.; Jahn, G.C. Insecticide-induced increase in the protein content of male accessory glands and its effect on the fecundity of females in the brown planthopper Nilaparvata lugens Stål (Hemiptera: Delphacidae). Crop Prot. 2010, 29, 1280–1285. [Google Scholar] [CrossRef]
- Suri, K.S.; Singh, G. Insecticide-induced resurgence of the whitebacked planthopper Sogatella furcifera (Horvath) (Hemiptera: Delphacidae) on rice varieties with different levels of resistance. Crop Prot. 2011, 30, 118–124. [Google Scholar] [CrossRef]
- Joshi, R.; Shepard, B.; Kenmore, P.; Lydia, R. Insecticide-induced resurgence of brown planthopper (BPH) on IR62. Int. Rice Res. Newsl. 1992, 17, 9–10. [Google Scholar]
- Horgan, F.G.; Peñalver-Cruz, A.; Almazan, M.L.P. Rice resistance buffers against the induced enhancement of brown planthopper fitness by some insecticides. Crops 2021, 1, 166–184. [Google Scholar] [CrossRef]
- Cheng, C.; Chang, W. Studies on varietal resistance to the brown planthopper in Taiwan. In Brown Planthopper: Threat to Rice Production in Asia; International Rice Research Institute: Los Baños, Philippines, 1979; pp. 251–271. [Google Scholar]
- Horgan, F.G.; Garcia, C.P.F.; Haverkort, F.; de Jong, P.W.; Ferrater, J.B. Changes in insecticide resistance and host range performance of planthoppers artificially selected to feed on resistant rice. Crop Prot. 2020, 127, 104963. [Google Scholar] [CrossRef] [PubMed]
- Matteson, P.C.; Gallagher, K.D.; Kenmore, P.E. Extension of integrated pest management for planthoppers in Asian irrigated rice: Empowering the user. In Planthoppers; Denno, R.F., Perfect, T.J., Eds.; Springer: Boston, MA, USA, 1994; pp. 656–685. [Google Scholar]
- Thorburn, C. The rise and demise of integrated pest management in rice in Indonesia. Insects 2015, 6, 381–408. [Google Scholar] [CrossRef]
- Pedigo, L.P.; Hutchins, S.H.; Higley, L.G. Economic injury levels in theory and practice. Annu. Rev. Entomol. 1986, 31, 341–368. [Google Scholar] [CrossRef]
- Bandong, J.P.; Canapi, B.L.; dela Cruz, C.G.; Litsinger, J.A. Insecticide decision protocols: A case study of untrained Filipino rice farmers. Crop Prot. 2002, 21, 803–816. [Google Scholar] [CrossRef]
- Horgan, F.G.; Kudavidanage, E.P. Use and avoidance of pesticides as responses by farmers to change impacts in rice ecosystems of southern Sri Lanka. Environ. Manag. 2020, 65, 787–803. [Google Scholar] [CrossRef]
- Horgan, F.G.; Ramal, A.F.; Bentur, J.S.; Kumar, R.; Bhanu, K.V.; Sarao, P.S.; Iswanto, E.H.; Chien, H.V.; Phyu, M.H.; Bernal, C.C.; et al. Virulence of brown planthopper (Nilaparvata lugens) populations from South and South East Asia against resistant rice varieties. Crop Prot. 2015, 78, 222–231. [Google Scholar] [CrossRef] [Green Version]
- Horgan, F.G.; Srinivasan, T.S.; Bentur, J.S.; Kumar, R.; Bhanu, K.V.; Sarao, P.S.; Chien, H.V.; Almazan, M.L.P.; Bernal, C.C.; Ramal, A.F.; et al. Geographic and research center origins of rice resistance to Asian planthoppers and leafhoppers: Implications for rice breeding and gene deployment. Agronomy 2017, 7, 62. [Google Scholar] [CrossRef] [Green Version]
- Heinrichs, E.A.; Fabellar, L.T.; Basilio, R.P.; Wen, T.-C.; Medrano, F. Susceptibility of rice planthoppers Nilaparvata lugens and Sogatella furcifera (Homoptera: Delphacidae) to insecticides as influenced by level of resistance in the host plant. Environ. Entomol. 1984, 13, 455–458. [Google Scholar] [CrossRef]
- Khush, G.S.; Virk, P.S. IR Varieties and Their Impact; International Rice Research Institute: Los Baños, Philippines, 2005. [Google Scholar]
- Nguyen, C.D.; Zheng, S.-H.; Sanada-Morimura, S.; Matsumura, M.; Yasui, H.; Daisuke, F. Substitution mapping and characterization of brown planthopper resistance genes from indica rice variety, ‘PTB33’ (Oryza sativa L.). Breed. Sci. 2021, 71, 5. [Google Scholar] [CrossRef]
- Horgan, F.G.; Peñalver Cruz, A.; Bernal, C.C.; Ramal, A.F.; Almazan, M.L.P.; Wilby, A. Resistance and tolerance to the brown planthopper, Nilaparvata lugens (Stål), in rice infested at different growth stages across a gradient of nitrogen applications. Field Crops Res. 2018, 217, 53–65. [Google Scholar] [CrossRef] [PubMed]
- Peñalver Cruz, A.; Arida, A.; Heong, K.L.; Horgan, F.G. Aspects of brown planthopper adaptation to resistant rice varieties with the Bph3 gene. Entomol. Exp. Appl. 2011, 141, 245–257. [Google Scholar] [CrossRef]
- Mackill, D.J.; Khush, G.S. IR64: A high-quality and high-yielding mega variety. Rice 2018, 11, 18. [Google Scholar] [CrossRef] [Green Version]
- Satapathy, M.K.; Anjaneyulu, A. Use of cypermethrin, a synthetic pyrethroid, in the control of rice tungro virus disease and its vector. Trop. Pest Manag. 1984, 30, 170–178. [Google Scholar] [CrossRef]
- Zhang, C.P.; He, H.M.; Yu, J.Z.; Hu, X.Q.; Zhu, Y.H.; Wang, Q. Residues of carbosulfan and its metabolites carbofuran and 3-hydroxy carbofuran in rice field ecosystem in China. J. Environ. Sci. Health Part B 2016, 51, 351–357. [Google Scholar] [CrossRef] [PubMed]
- Samanta, S.; Barman, M.; Samanta, N.R.A. Bio-efficacy trials of carbofuran 3% CG against insect pests of rice. J. Entomol. Zool. Stud. 2020, 8, 1529–1533. [Google Scholar]
- Kumar, R.; Singh, B. Persistence and metabolism of Fipronil in rice (Oryza sativa Linnaeus) field. Bull. Environ. Contam. Toxicol. 2013, 90, 482–488. [Google Scholar] [CrossRef]
- Dhaka, S.; Prajapati, C.; Singh, D.; Singh, R. Field evaluation of insecticides and bio-pesticides against rice leaf folder, Cnaphalocrocis Medinalis (Guenee). Ann. Plant Prot. Sci. 2011, 19, 324–326. [Google Scholar]
- Horgan, F.G.; Mundaca, E.A.; Quintana, R.; Naredo, A.I.; Almazan, M.L.P.; Bernal, C.C. Efficacy and cost-effectiveness of phenotyping for rice resistance and tolerance to planthoppers. Insects 2021, 12, 847. [Google Scholar] [CrossRef]
- Addelman, S. Computing the analysis of variance table for experiments involving qualitative factors and zero amounts of quantitative factors. Am. Stat. 1974, 28, 21–22. [Google Scholar]
- Horgan, F.G.; Naik, B.S.; Iswanto, E.H.; Almazan, M.L.P.; Ramal, A.F.; Bernal, C.C. Responses by the brown planthopper, Nilaparvata lugens, to conspecific density on resistant and susceptible rice varieties. Entomol. Exp. Et Appl. 2016, 158, 284–294. [Google Scholar] [CrossRef]
- Nagata, T. Timing of buprofezin application for control of the brown planthopper, Nilaparvata lugens Stål (Homoptera: Delphacidae). Appl. Entomol. Zool. 1986, 21, 357–362. [Google Scholar] [CrossRef]
- Hamid, A. Nitrogen and carbofuran effects on the growth and yield performance of mungbean (Vigna radiata [L.] Wilczek). J. Agron. Crop Sci. 1988, 161, 11–16. [Google Scholar] [CrossRef]
- Kim, S.-C. Effect of carbofuran on rice growth. Korean J. Weed Sci. 1987, 7, 98–106. [Google Scholar]
- Nayak, A.; Mahapatra, P.; Behera, K. Biochemical changes in rice plant due to application of carbofuran. J. Entomol. Zool. Stud. 2017, 5, 1177–1180. [Google Scholar]
- Borkar, V.; Gokhale, N.; Khobragade, N.; More, S.; Dhopavkar, R. Influence of phorate and carbofuran insecticides on nitrogen availability and their residues in soil and rice. Int. J. Chem. Stud. 2018, 6, 9–14. [Google Scholar]
- Anand Kumar, A.D.V.S.L.P.; Rama Rao, C.V.; Mallikharjuna Rao, N.; Krishnam Raju, S.; Nafeez Umar, S.K. Influence of certain insecticides on the resurgence of rice brown planthopper, Nilaparvata lugens (Stål). J. Entomol. Zool. Stud. 2019, 7, 874–878. [Google Scholar]
- Anand Kumar, A.D.V.S.L.P.; Rama Rao, C.V.; Mallikharjuna Rao, N. Insecticides and resurgence of rice brown planthopper Nilaparvata lugens (Stal). Indian J. Entomol. 2020, 82, 809–812. [Google Scholar] [CrossRef]
- Heinrichs, E.; Viajante, F.; Aquino, G. Resurgence-inducing insecticides as a tool in field screening of rice against the brown planthopper. Int. Rice Res. Newsl. 1978, 3, 10–11. [Google Scholar]
- Heinrichs, E.A.; Aquino, G.B.; Chelliah, S.; Valencia, S.L.; Reissig, W.H. Resurgence of Nilaparvata lugens (Stål) populations as influenced by method and timing of insecticide applications in lowland rice. Environ. Entomol. 1982, 11, 78–84. [Google Scholar] [CrossRef]
- Vorley, W. Spider mortality implicated in insecticide-induced resurgence of whitebacked planthopper (WBPH) and brown planthopper (BPH) in Kedah, Malaysia. Int. Rice Res. Newsl. 1987, 10, 19–20. [Google Scholar]
- Horgan, F.G.; de Freitas, T.F.S.; Crisol-Martínez, E.; Mundaca, E.A.; Bernal, C.C. Nitrogenous fertilizer reduces resistance but enhances tolerance to the brown planthopper in fast-growing, moderately resistant rice. Insects 2021, 12, 989. [Google Scholar] [CrossRef]
- Gallagher, K.D.; Kenmore, P.E.; Sogawa, K. Judicial use of insecticides deter planthopper outbreaks and extend the life of resistant varieties in Southeast Asian rice. In Planthoppers; Denno, R.F., Perfect, T.J., Eds.; Springer: Boston, MA, USA, 1994; pp. 599–614. [Google Scholar]
- Bragança, I.; Grosso, C.; Rede, D.; Sousa, S.R.; Lemos, P.C.; Domingues, V.F. Ecotoxicological effects of insecticides in plants assessed by germination and other phytotoxicity tools. In Biotic and Abiotic Stress Tolerance in Plants; Vats, S., Ed.; Springer: Singapore, 2018; pp. 47–76. [Google Scholar]
- Giménez–Moolhuyzen, M.; van der Blom, J.; Lorenzo–Mínguez, P.; Cabello, T.; Crisol–Martínez, E. Photosynthesis inhibiting effects of pesticides on sweet pepper leaves. Insects 2020, 11, 69. [Google Scholar] [CrossRef] [Green Version]
- Moriya, S.; Noda, M.; Haraguchi, H. On the growth inhibition of rice seedlings by Cartap insecticide. Proc. Assoc. Plant Prot. Kyushu 1975, 21, 68–69. [Google Scholar]
- Moore, M.T.; Kröger, R. Effect of three insecticides and two herbicides on rice (Oryza sativa) seedling germination and growth. Arch. Environ. Contam. Toxicol. 2010, 59, 574–581. [Google Scholar] [CrossRef]
- Jahn, G.C.; Domingo, I.; Almazan, M.L.P.; Pacia, J. Effect of rice bug Leptocorisa oratorius (Hemiptera: Alydidae) on rice yield, grain quality, and seed viability. J. Econ. Entomol. 2004, 97, 1923–1927. [Google Scholar] [CrossRef] [PubMed]
- Van Den Berg, H.; Soehardi. The influence of the rice bug Leptocorisa oratorius on rice yield. J. Appl. Ecol. 2000, 37, 959–970. [Google Scholar] [CrossRef]
- Chelliah, S.; Heinrichs, E.A. Factors affecting insecticide-induced resurgence of the brown planthopper, Nilaparvata lugens on rice. Environ. Entomol. 1980, 9, 773–777. [Google Scholar] [CrossRef]
- Cuong, N.L.; Ben, P.T.; Phuong, L.T.; Chau, L.M.; Cohen, M.B. Effect of host plant resistance and insecticide on brown planthopper Nilaparvata lugens (Stål) and predator population development in the Mekong Delta, Vietnam. Crop Prot. 1997, 16, 707–715. [Google Scholar] [CrossRef]
- Yang, G.-Q.; Gao, X.; Zhang, N.-N.; Chen, D.-Y.; Liu, F.; Xu, J.-X.; Wu, J.-C. Pesticide-induced changes in fecundity and rice stripe virus transmission ability in Laodelphax striatellus (Homoptera: Delphacidae). J. Asia-Pac. Entomol. 2017, 20, 830–834. [Google Scholar] [CrossRef]
- Deng, L.; Xu, M.; Cao, H.; Dai, J. Ecotoxicological effects of buprofezin on fecundity, growth, development, and predation of the wolf spider Pirata piratoides (Schenkel). Arch. Environ. Contam. Toxicol. 2008, 55, 652. [Google Scholar] [CrossRef] [PubMed]
- Ko, K.; Liu, Y.; Hou, M.; Babendreier, D.; Zhang, F.; Song, K. Toxicity of insecticides targeting rice planthoppers to adult and immature stages of Trichogramma chilonis (Hymenoptera: Trichogrammatidae). J. Econ. Entomol. 2015, 108, 69–76. [Google Scholar] [CrossRef]
- Tanaka, K.; Endo, S.; Kazano, H. Toxicity of insecticides to predators of rice planthoppers: Spiders, the mirid bug and the dryinid wasp. Appl. Entomol. Zool. 2000, 35, 177–187. [Google Scholar] [CrossRef]
- Wang, H.Y.; Yang, Y.; Su, J.Y.; Shen, J.L.; Gao, C.F.; Zhu, Y.C. Assessment of the impact of insecticides on Anagrus nilaparvatae (Pang et Wang) (Hymenoptera: Mymanidae), an egg parasitoid of the rice planthopper, Nilaparvata lugens (Hemiptera: Delphacidae). Crop Prot. 2008, 27, 514–522. [Google Scholar] [CrossRef]
- Pawar, R.D.; Sawai, H.R.; Thakare, V.S.; Wankhade, S.V. Evaluation of safety of some new insecticides to per cent parasitisation and survival of Trichogramma japonicum. J. Entomol. Zool. Stud. 2020, 8, 685–689. [Google Scholar]
- Rubia, E. Biological and Toxicological Studies of the Cricket Metioche Vittaticollis (Stål)(Orthoptera: Gryllidae), a Predator of Rice Insect Pests [Philippines]; University of the Philippines: Los Baños, Philippines, 1986. [Google Scholar]
- Kim, J.; Cho, D.; Hah, J.; Chang, S.; Bark, Y. Effect of density variation on the natural enemies and rice pests by application method and time of carbofuran granules. Korean J. Appl. Entomol. 1984, 23, 233–236. [Google Scholar]
- Jafar, W.; Mazlan, N.; Adam, N.A.; Omar, D. Evaluation on the effects of insecticides on biodiversity of arthropod in rice ecosystem. Acta Biol. Malays. 2013, 2, 115–123. [Google Scholar]
- Parsaeyan, E.; Safavi, S.A.; Saber, M.; Poorjavad, N. Effects of emamectin benzoate and cypermethrin on the demography of Trichogramma brassicae Bezdenko. Crop Prot. 2018, 110, 269–274. [Google Scholar] [CrossRef]
- Fabellar, L.T.; Heinrichs, E.A. Relative toxicity of insecticides to rice planthoppers and leafhoppers and their predators. Crop Prot. 1986, 5, 254–258. [Google Scholar] [CrossRef]
- Pazini, J.B.; Grützmacher, A.D.; Martins, J.F.S.; Pasini, R.A.; Rakes, M. Selectivity of pesticides used in rice crop on Telenomus podisi and Trichogramma pretiosum. Pesqui. Agropecuária Trop. 2016, 46, 327–335. [Google Scholar] [CrossRef] [Green Version]
- Nakanishi, K.; Yokomizo, H.; Hayashi, T.I. Were the sharp declines of dragonfly populations in the 1990s in Japan caused by fipronil and imidacloprid? An analysis of Hill’s causality for the case of Sympetrum frequens. Environ. Sci. Pollut. Res. 2018, 25, 35352–35364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hegde, M.; Nidagundi, J. Effect of newer chemicals on planthoppers and their mirid predator in rice. Karnataka J. Agric. Sci. 2009, 22, 511–513. [Google Scholar]
- Horgan, F.G.; Arida, A.; Ardestani, G.; Almazan, M.L.P. Elevated temperatures diminish the effects of a highly resistant rice variety on the brown planthopper. Sci. Rep. 2021, 11, 262. [Google Scholar] [CrossRef] [PubMed]
- Horgan, F.G.; Arida, A.; Ardestani, G.; Almazan, M.L.P. Intraspecific competition counters the effects of elevated and optimal temperatures on phloem-feeding insects in tropical and temperate rice. PLoS ONE 2020, 15, e0240130. [Google Scholar] [CrossRef] [PubMed]
- Horgan, F.G.; Crisol Martínez, E.; Stuart, A.M.; Bernal, C.C.; de Cima Martín, E.; Almazan, M.L.P.; Ramal, A.F. Effects of vegetation strips, fertilizer levels and varietal resistance on the integrated management of arthropod biodiversity in a tropical rice ecosystem. Insects 2019, 10, 328. [Google Scholar] [CrossRef] [Green Version]
- Uddin, A.; Islam, K.S.; Jahan, M.; Ara, A.; Khan, M.A.I. Factors influencing the resurgence of brown planthopper in Bangladesh. SAARC J. Agric. 2020, 18, 117–128. [Google Scholar] [CrossRef]
- Otieno, P.O.; Lalah, J.O.; Virani, M.; Jondiko, I.O.; Schramm, K.-W. Soil and water contamination with carbofuran residues in agricultural farmlands in Kenya following the application of the technical formulation Furadan. J. Environ. Sci. Health Part B 2010, 45, 137–144. [Google Scholar] [CrossRef]
- Ghosal, A.; Hati, A. Impact of some new generation insecticides on soil arthropods in rice maize cropping system. J. Basic Appl. Zool. 2019, 80, 6. [Google Scholar] [CrossRef]
- González-Doncel, M.; Fernández-Torija, C.; Hinton, D.E.; Tarazona, J.V. Stage-specific toxicity of cypermethrin to medaka (Oryzias latipes) eggs and embryos using a refined methodology for an in vitro fertilization bioassay. Arch. Environ. Contam. Toxicol. 2004, 48, 87–98. [Google Scholar] [CrossRef]
- Sharma, R.K.; Rai, D.K.; Sharma, B. In-vitro carbofuran induced micronucleus formation in human blood lymphocytes. Cell. Mol. Biol. 2012, 58, 128–133. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Categories of Outcome 1 | Outcome Criteria | Tests | Test Consequences 3 |
|---|---|---|---|
| Resistance effective | BPH densities on resistant rice are lower than densities on susceptible rice; yields are higher on resistant rice | Comparison of final planthopper densities and grain yields on BPH-infested IR62 and IR64 | Significantly lower BPH densities and/or higher yields on IR62 |
| Insecticide effective 1 | BPH densities lower on insecticide-treated rice than on non-treated rice; yields are higher on treated rice | Comparison of final planthopper densities on insecticide-treated and non-treated IR62 and/or IR64 infested with BPH | Significantly lower BPH densities on treated plants |
| Insecticide resurgence-causing 1 | BPH densities higher on insecticide-treated rice than on non-treated rice | Comparison of final planthopper densities on insecticide-treated and non-treated IR62 and/or IR64 infested with BPH | Significantly higher BPH densities on treated plants |
| No benefit (redundant use of insecticide) | Applying insecticide to resistant rice produces no reduction in BPH densities or no increase in rice yields | Comparison of final planthopper densities and grain yields on insecticide-treated and non-treated IR62 infested with BPH | No significant difference between BPH densities, or yields on insecticide-treated and non-treated IR62 |
| No benefit (redundant exposure of variety) | Applying an effective insecticide to resistant and susceptible rice results in similar BPH densities or similar yield increases irrespective of host resistance | Comparison of final planthopper densities and grain yields on insecticide-treated and non-treated IR62 and IR64 infested with BPH | Significant variety × insecticide effect due to lower BPH densities/higher yields on untreated IR62 compared to untreated IR64, but similar densities and yields on insecticide-treated plants irrespective of variety |
| Insecticides and resistance synergistic 2 | Applying insecticide to resistant rice reduces BPH densities or increases rice yields more than on non-treated resistant rice and treated susceptible rice | Comparison of final planthopper densities and grain yields on insecticide-treated and non-treated IR62 and IR64 infested with BPH | Significant insecticide and/or variety effects with or without significant interactions depicting lower densities and higher yields on treated IR62 compared to non-treated IR62 |
| Insecticides antagonistic to resistance | Applying insecticide to resistant rice results in higher BPH densities and/or lower yields than on non-treated resistant rice thereby producing similar BPH densities and/or yields on resistant and susceptible rice | Comparison of final planthopper densities and grain yields on insecticide-treated and non-treated IR62 and IR64 infested with BPH | Significant variety × insecticide interaction due to similar densities of BPH and/or yields on treated IR62 and treated IR64, but lower densities and/or higher yields on nontreated IR62 compared to non-treated IR64 |
| Resistance buffers against antagonistic effects | Applying a resurgence-causing insecticide to resistant rice has no effect on BPH densities or rice yields, but the same insecticide on susceptible rice increases BPH densities and/or decreases yields | Comparison of final planthopper densities and grain yields on IR62 and IR64 treated with resurgence-causing insecticide and infested with BPH | Significant variety effect maintained after BPH densities significantly increase on IR64, but not on IR62 |
| Insecticide phytotoxic | Applying insecticide to non-BPH infested resistant or susceptible rice reduces plant vigor, possibly reducing yields | Comparisons of insecticide-treated and non-treated IR62 and/or IR64 plants without BPH infestation | Significant reduction in plant biomass and/or yields |
| Insecticide stimulates plant development | Applying insecticide to non-BPH infested resistant or susceptible rice increases plant vigor, possibly increasing yields | Comparisons of insecticide-treated and non-treated IR62 and/or IR64 plants without BPH infestation | Significant increase in plant biomass and/or yields |
| Variety and Insecticide | Number of Applications | Weight of Filled Grains (g Dry Weight) 1,2 | Number of Filled Grain 1 | Proportion of Grain Unfilled 1 | 1000 Grain Weight 1 |
|---|---|---|---|---|---|
| IR62 | |||||
| Buprofezin | 1 | 6.15 ± 1.98 ab | 314.00 ± 96.98 ab | 0.25 ± 0.04 | 19.44 ± 0.67 ab |
| 2 | 5.88 ± 2.14 | 298.40 ± 105.67 | 0.28 ± 0.10 | 19.83 ± 1.26 | |
| 3 | 5.93 ± 2.09 | 342.00 ± 118.25 | 0.31 ± 0.17 | 15.73 ± 1.81 | |
| Carbofuran | 1 | 4.90 ± 1.84 d | 262.60 ± 86.08 d | 0.46 ± 0.09 | 17.76 ± 1.37 b |
| 2 | 7.34 ± 1.51 | 391.60 ± 81.14 | 0.40 ± 0.12 | 18.82 ± 0.27 | |
| 3 | 10.32 ± 1.09 | 557.60 ± 44.02 | 0.10 ± 0.02 | 18.34 ± 0.58 | |
| Cartap hydrochloride | 1 | 8.62 ± 1.80 abcd | 425.20 ± 88.25 abcd | 0.24 ± 0.09 | 20.17 ± 0.53 b |
| 2 | 6.14 ± 2.54 | 317.20 ± 132.04 | 0.20 ± 0.05 | 19.39 ± 0.34 | |
| 3 | 5.93 ± 1.99 | 319.40 ± 102.64 | 0.32 ± 0.13 | 18.11 ± 0.93 | |
| Cypermethrin | 1 | 7.95 ± 0.83 abc | 428.00 ± 34.52 abc | 0.38 ± 0.04 | 18.44 ± 0.56 ab |
| 2 | 6.23 ± 2.22 | 317.20 ± 113.30 | 0.19 ± 0.05 | 19.63 ± 0.31 | |
| 3 | 7.51 ± 1.95 | 411.20 ± 110.99 | 0.13 ± 0.03 | 18.44 ± 0.60 | |
| Deltamethrin | 1 | 5.22 ± 2.21 a | 273.40 ± 114.52 a | 0.34 ± 0.02 | 19.04 ± 0.57 a |
| 2 | 3.35 ± 1.96 | 204.80 ± 108.93 | 0.34 ± 0.14 | 13.85 ± 3.34 | |
| 3 | 5.04 ± 2.46 | 267.40 ± 123.69 | 0.27 ± 0.05 | 18.39 ± 0.94 | |
| Fipronil | 1 | 6.93 ± 2.09 cd | 360.20 ± 98.25 cd | 0.31 ± 0.06 | 18.81 ± 1.56 b |
| 2 | 8.03 ± 2.15 | 427.20 ± 110.36 | 0.13 ± 0.02 | 18.69 ± 0.92 | |
| 3 | 6.46 ± 0.82 | 361.80 ± 41.57 | 0.28 ± 0.08 | 17.85 ± 0.71 | |
| Thiamethoxam + chlorantraniliprole | 1 | 7.07 ± 3.03 bcd | 321.60 ± 133.66 bcd | 0.24 ± 0.01 | 21.81 ± 1.70 b |
| 2 | 5.48 ± 2.14 | 283.40 ± 112.96 | 0.28 ± 0.15 | 19.62 ± 0.53 | |
| 3 | 4.49 ± 1.99 | 245.60 ± 106.75 | 0.29 ± 0.06 | 18.10 ± 0.60 | |
| Control | 5.66 ± 0.61 | 288.60 ± 22.19 | 0.44 ± 0.05 | 17.59 ± 0.66 | |
| IR64 | |||||
| Buprofezin | 1 | 0.61 ± 0.61 | 42.00 ± 42.00 | 0.54 | 14.48 |
| 2 | 0.71 ± 0.66 | 3.20 ± 1.96 | 0.98 ± 0.00 | 12.50 ± 2.50 | |
| 3 | 2.69 ± 1.50 | 140.20 ± 72.80 | 0.38 ± 0.18 | 18.48 ± 1.70 | |
| Carbofuran | 1 | 6.36 ± 2.57 | 292.40 ± 115.64 | 0.49 ± 0.18 | 20.51 ± 1.45 |
| 2 | 9.46 ± 1.40 | 447.20 ± 61.31 | 0.20 ± 0.05 | 21.02 ± 0.50 | |
| 3 | 8.96 ± 0.48 | 414.80 ± 20.63 | 0.21 ± 0.03 | 21.60 ± 0.46 | |
| Cartap hydrochloride | 1 | 2.18 ± 1.35 | 113.20 ± 66.93 | 0.61 ± 0.13 | 18.41 ± 1.05 |
| 2 | 3.37 ± 1.25 | 166.80 ± 58.64 | 0.46 ± 0.11 | 19.87 ± 0.81 | |
| 3 | 4.18 ± 1.58 | 217.40 ± 76.18 | 0.32 ± 0.10 | 18.75 ± 0.99 | |
| Cypermethrin | 1 | 0.00 ± 0.00 | 0.00 ± 0.00 | NG | NG |
| 2 | 1.57 ± 0.61 | 86.00 ± 27.56 | 0.73 ± 0.04 | 17.97 ± 4.80 | |
| 3 | 3.28 ± 1.81 | 178.00 ± 87.05 | 0.59 ± 0.16 | 16.02 ± 1.97 | |
| Deltamethrin | 1 | 0.06 ± 0.06 | 0.30 ± 0.30 | NG | NG |
| 2 | 0.92 ± 0.92 | 52.40 ± 52.40 | 0.31 | 17.63 | |
| 3 | 0.46 ± 0.28 | 44.00 ± 27.13 | 0.58 ± 0.18 | 10.59 ± 0.51 | |
| Fipronil | 1 | 6.70 ± 1.56 | 315.60 ± 72.69 | 0.36 ± 0.15 | 20.53 ± 0.85 |
| 2 | 7.06 ± 1.67 | 327.20 ± 68.98 | 0.31 ± 0.12 | 20.83 ± 0.93 | |
| 3 | 8.24 ± 1.30 | 389.80 ± 70.96 | 0.24 ± 0.07 | 21.44 ± 0.63 | |
| Thiamethoxam + chlorantraniliprole | 1 | 5.52 ± 1.68 | 285.40 ± 89.64 | 0.40 ± 0.08 | 19.39 ± 1.06 |
| 2 | 5.79 ± 1.52 | 283.20 ± 62.86 | 0.34 ± 0.10 | 19.65 ± 0.93 | |
| 3 | 6.90 ± 1.75 | 353.80 ± 89.53 | 0.27 ± 0.12 | 18.69 ± 1.58 | |
| Control | 0.43 ± 0.19 | 14.90 ± 9.53 | 0.77 ± 0.09 | 15.91 ± 0.93 | |
| F-variety (V) | 15.938 *** | 22.824 *** | 12.982 *** | 0.555 ns | |
| F-treatment (T) | 6.052 *** | 5.442 *** | 1.554 ns | 3.066 ** | |
| F-applications | 0.876 ns | 1.537 ns | 1.976 ns | 0.506 ns | |
| F-V × T | 3.669 *** | 3.305 ** | 2.175 ns | 2.960 * | |
| F-control (C) | 2.508 ns | 2.868 ns | 5.405 * | 2.256 ns | |
| F-C × V | 19.985 *** | 27.325 *** | 11.886 *** | 1.579 ns |
| Variety and Insecticide | Application Time (Days) | Weight of Filled Grains (g Dry Weight) 1 | Number of Filled Grain 1 | Proportion of Grain Unfilled 1 | 1000 Grain Weight 1 |
|---|---|---|---|---|---|
| IR62 | |||||
| Buprofezin | 20 | 6.15 ± 1.98 ab | 260.00 ± 116.23 ab | 0.54 ± 0.19 abc | 19.45 ± 0.54 ab |
| 50 | 8.62 ± 0.80 | 462.40 ± 29.44 | 0.17 ± 0.03 | 18.54 ± 0.82 | |
| Carbofuran | 20 | 4.90 ± 1.84 b | 220.60 ± 101.38 b | 0.65 ± 0.15 a | 18.62 ± 0.80 b |
| 50 | 8.10 ± 2.05 | 407.80 ± 103.23 | 0.33 ± 0.17 | 19.85 ± 0.22 | |
| Cartap hydrochloride | 20 | 8.62 ± 1.80 ab | 425.20 ± 88.25 ab | 0.24 ± 0.09 abc | 20.17 ± 0.53 a |
| 50 | 7.04 ± 0.84 | 358.40 ± 45.42 | 0.30 ± 0.08 | 19.75 ± 0.49 | |
| Cypermethrin | 20 | 7.95 ± 0.83 ab | 428.00 ± 34.52 ab | 0.38 ± 0.04 abc | 18.44 ± 0.56 ab |
| 50 | 4.95 ± 2.00 | 255.00 ± 104.26 | 0.56 ± 0.18 | 19.42 ± 0.37 | |
| Deltamethrin | 20 | 5.22 ± 2.21 a | 273.40 ± 114.52 a | 0.60 ± 0.16 c | 19.13 ± 0.32 ab |
| 50 | 5.97 ± 1.60 | 248.20 ± 101.93 | 0.53 ± 0.19 | 19.65 ± 0.25 | |
| Fipronil | 20 | 6.93 ± 2.09 ab | 360.20 ± 98.25 ab | 0.45 ± 0.15 ab | 18.65 ± 1.22 ab |
| 50 | 5.89 ± 1.54 | 240.80 ± 98.89 | 0.52 ± 0.20 | 19.56 ± 0.26 | |
| Thiamethoxam + chlorantraniliprole | 20 | 7.07 ± 3.03 ab | 321.60 ± 133.66 ab | 0.55 ± 0.19 abc | 21.69 ± 0.94 b |
| 50 | 5.14 ± 2.45 | 266.20 ± 119.80 | 0.54 ± 0.19 | 18.59 ± 0.83 | |
| Control | 4.47 ± 1.27 | 220.60 ± 57.87 | 0.55 ± 0.12 | 20.76 ± 2.89 | |
| IR64 | |||||
| Buprofezin | 20 | 0.61 ± 0.61 | 0.90 ± 0.90 | 1.00 ± 0.00 | NG |
| 50 | 1.17 ± 0.44 | 61.00 ± 25.21 | 0.81 ± 0.08 | 18.03 ± 0.74 | |
| Carbofuran | 20 | 6.36 ± 2.57 | 292.40 ± 115.64 | 0.49 ± 0.18 | 20.50 ± 1.12 |
| 50 | 6.82 ± 0.72 | 301.40 ± 31.18 | 0.16 ± 0.03 | 22.58 ± 0.25 | |
| Cartap hydrochloride | 20 | 2.18 ± 1.35 | 113.20 ± 66.93 | 0.76 ± 0.12 | 18.44 ± 0.60 |
| 50 | 1.37 ± 0.56 | 2.40 ± 2.40 | 0.99 ± 0.01 | 12.50 ± 0.22 | |
| Cypermethrin | 20 | 0.00 ± 0.00 | 0.00 ± 0.00 | 1.00 ± 0.00 | NG |
| 50 | 3.76 ± 1.99 | 175.80 ± 91.21 | 0.53 ± 0.20 | 18.11 ± 2.14 | |
| Deltamethrin | 20 | 0.06 ± 0.06 | 0.20 ± 0.20 | 1.00 ± 0.00 | NG |
| 50 | 0.05 ± 0.05 | 0.20 ± 0.20 | 1.00 ± 0.00 | NG | |
| Fipronil | 20 | 6.70 ± 1.56 | 315.60 ± 72.69 | 0.36 ± 0.15 | 20.53 ± 0.85 |
| 50 | 3.74 ± 1.45 | 196.40 ± 80.69 | 0.41 ± 0.16 | 19.82 ± 0.85 | |
| Thiamethoxam + chlorantraniliprole | 20 | 5.52 ± 1.68 | 285.40 ± 89.64 | 0.52 ± 0.14 | 19.11 ± 0.87 |
| 50 | 1.97 ± 1.97 | 88.20 ± 88.20 | 0.81 ± 0.19 | 22.46 ± 0.23 | |
| Control | 0.43 ± 0.19 | 5.70 ± 5.70 | 0.97 ± 0.03 | 16.57 ± 0.23 | |
| F-variety (V) | 34.375 *** | 37.296 *** | 20.712 *** | 4.689 * | |
| F-treatment (T) | 2.337 ** | 2.084 ns | 2.865 ** | 3.126 ** | |
| F-day | 0.192 ns | 0.308 ns | 0.586 ns | 0.098 ns | |
| F-V × T | 3.189 ** | 3.455 ** | 4.588 *** | 4.508 *** | |
| F-control (C) | 11.218 *** | 12.464 *** | 1.877 ns | 0.592 ns | |
| F-C × V | 3.926 * | 0.037ns | 0.891 ns | 6.087 ** |
| Insecticides | Experiment 1 (Application Frequency) | Experiment 2 (Application Timing) | ||||
|---|---|---|---|---|---|---|
| BPH Biomass Density | Rice Yield | Phytotoxic Effects | BPH Biomass Density | Rice Yield | Phytotoxic Effects | |
| Buprofezin | Ineffective; insecticide redundant | Ineffective; insecticide redundant | No | Ineffective; insecticide redundant at 20 DAS; synergy—tendency to reduce biomass density at 50 DAS | Ineffective; insecticide redundant | Tendency to reduce yield at 50 DAS |
| Cabofuran | Effective (2, 3 applications); resistance redundant | Effective (2, 3 applications); resistance redundant | No | Effective (50 DAS) | Tendency toward effective (50 DAS) | No |
| Cartap hydrochloride | Ineffective; insecticide redundant; synergy—tendency to reduce biomass density on IR62 after 1 application | Ineffective; insecticide redundant | No | Effective; synergy—tendency to reduce biomass density at 20 and 50 DAS | Effective (20 DAS); synergy—tendency to increase IR62 yields at 20 DAS | Tendency to reduce yield at 50 DAS |
| Cypermethrin | Ineffective (1 or 2 applications); Effective (3 applications); synergy—tendency to reduce biomass density on IR62 after 1 application | Ineffective | No | Ineffective; synergy—tendency to reduce biomass density on IR62 at 20 DAS | Ineffective; synergy—tendency to increase yields on IR62 at 20 DAS | No |
| Deltamethrin | Ineffective; antagonistic—tendency to increase biomass density on IR62, but not on IR64, after 3 applications | Ineffective | No | Ineffective; buffer—tendency for biomass density to increase on IR64 at 50 DAS, but not on IR62 | Ineffective | No |
| Fipronil | Effective (1, 2, and 3 applications); resistance redundant | Effective (1, 2, and 3 applications); resistance redundant | No | Ineffective; tendency to reduce biomass density at 20 and 50 DAS | Effective (20 DAS); tendency to increase yields at 50 DAS; resistance redundant | Tendency to reduce yields at 50 DAS |
| Thiamethoxam + chlorantraniliprole | Ineffective; insecticide redundant | Ineffective; tendency to increase yields of IR64; insecticide redundant | No | Ineffective; insecticide redundant | Ineffective; insecticide redundant | No |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Horgan, F.G.; Peñalver-Cruz, A. Compatibility of Insecticides with Rice Resistance to Planthoppers as Influenced by the Timing and Frequency of Applications. Insects 2022, 13, 106. https://doi.org/10.3390/insects13020106
Horgan FG, Peñalver-Cruz A. Compatibility of Insecticides with Rice Resistance to Planthoppers as Influenced by the Timing and Frequency of Applications. Insects. 2022; 13(2):106. https://doi.org/10.3390/insects13020106
Chicago/Turabian StyleHorgan, Finbarr G., and Ainara Peñalver-Cruz. 2022. "Compatibility of Insecticides with Rice Resistance to Planthoppers as Influenced by the Timing and Frequency of Applications" Insects 13, no. 2: 106. https://doi.org/10.3390/insects13020106