
Unexpected High Species Diversity of Mesolycus Gorham (Coleoptera, Lycidae) from China, with a Preliminary Investigation on Its Phylogenetic Position Based on Multiple Genes †

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Morphological and Geographical Studies
2.3. DNA Extraction, Sequencing and Annotation
2.4. Phylogenetic Analyses
3. Results
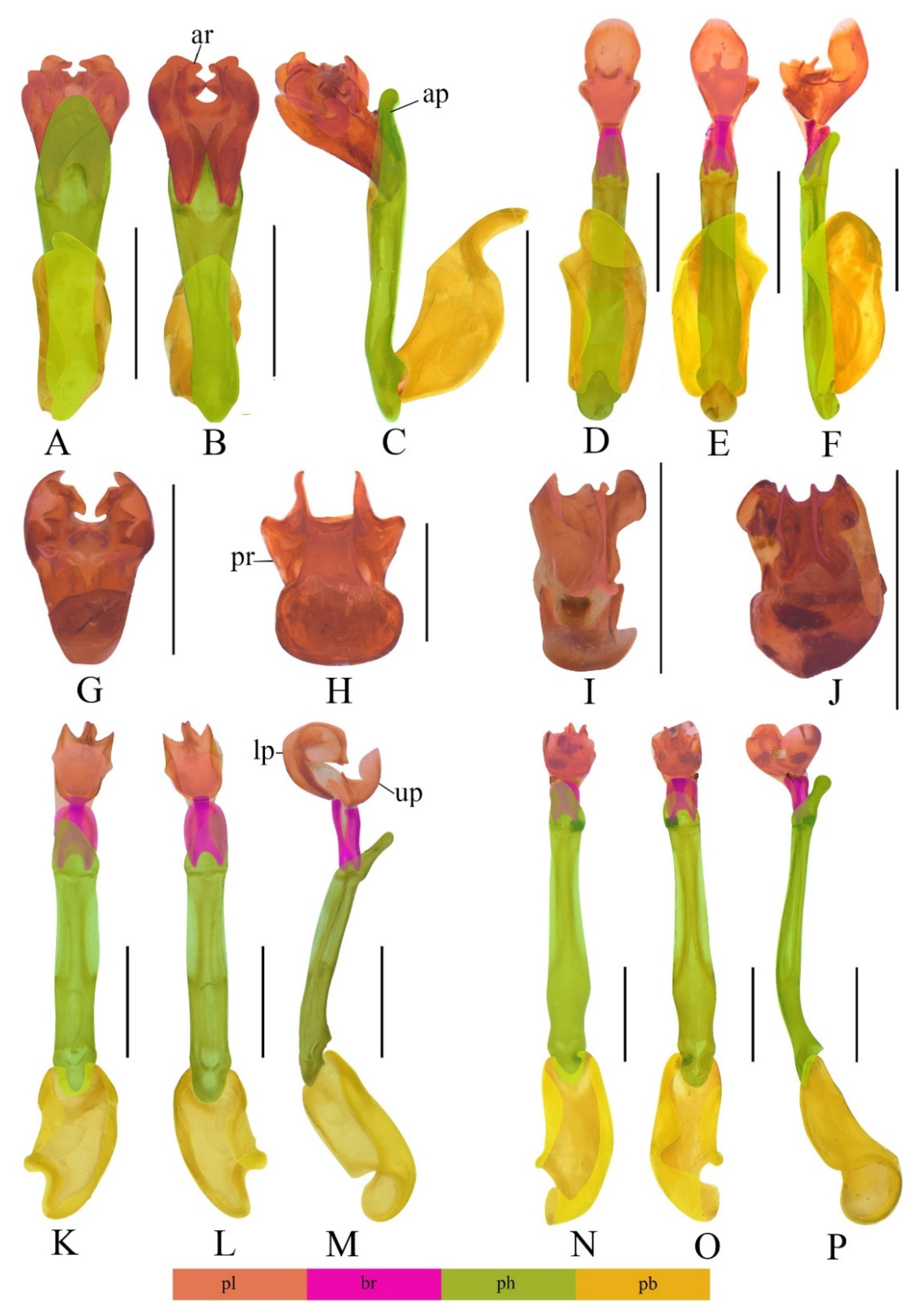
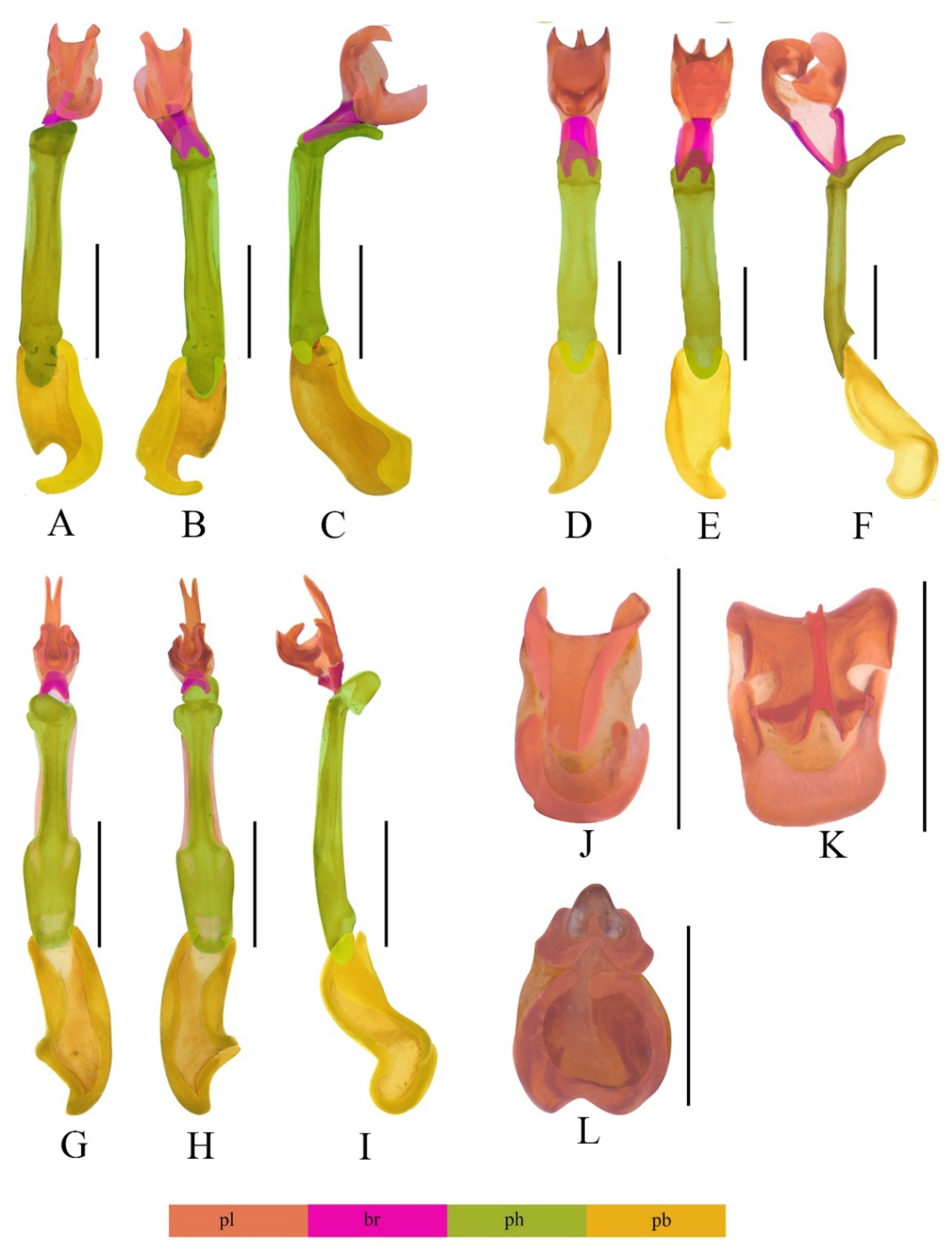
3.1. Descriptions of the Species
3.2. Taxonomic Notes
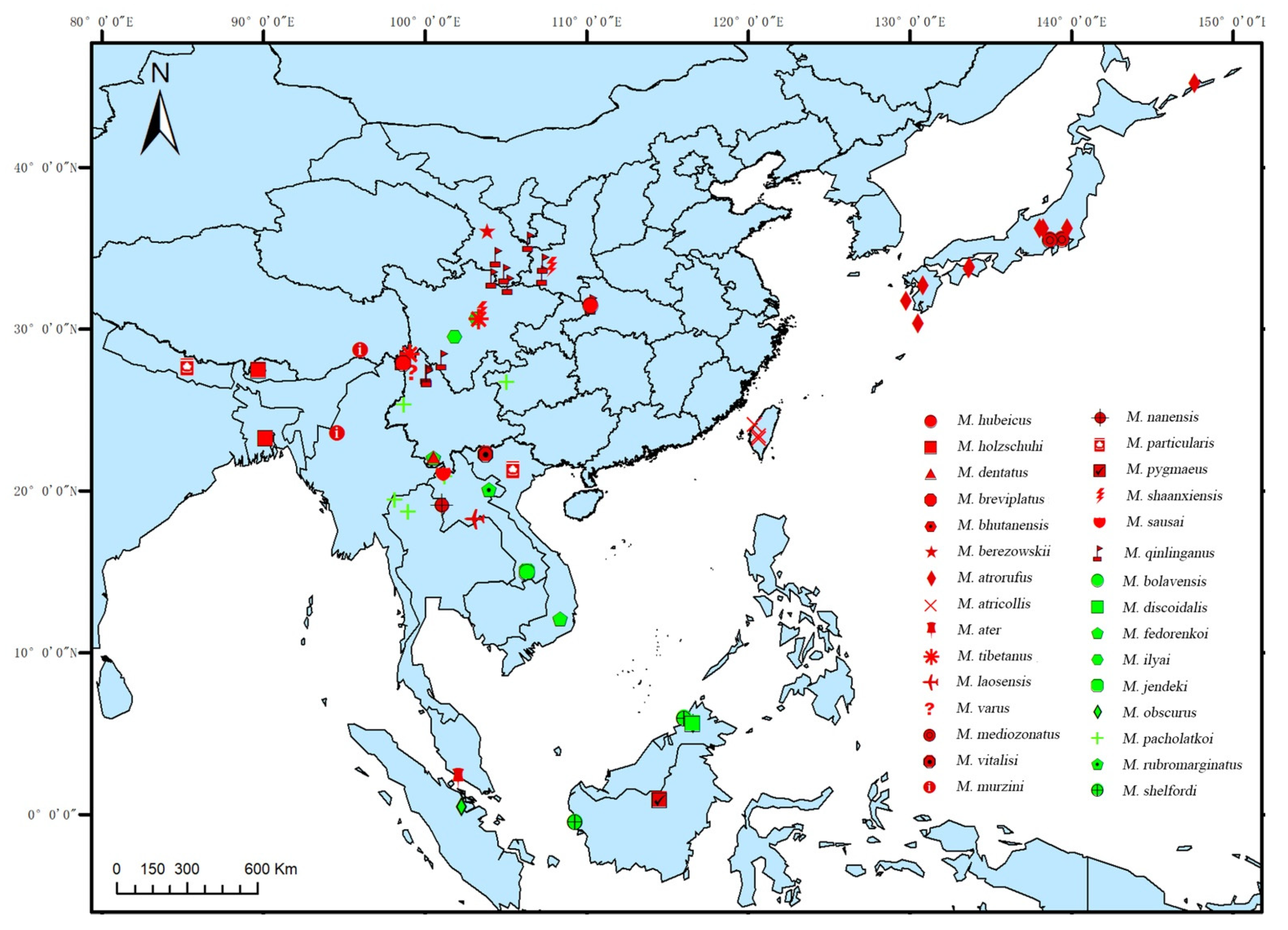
3.3. Distribution Pattern
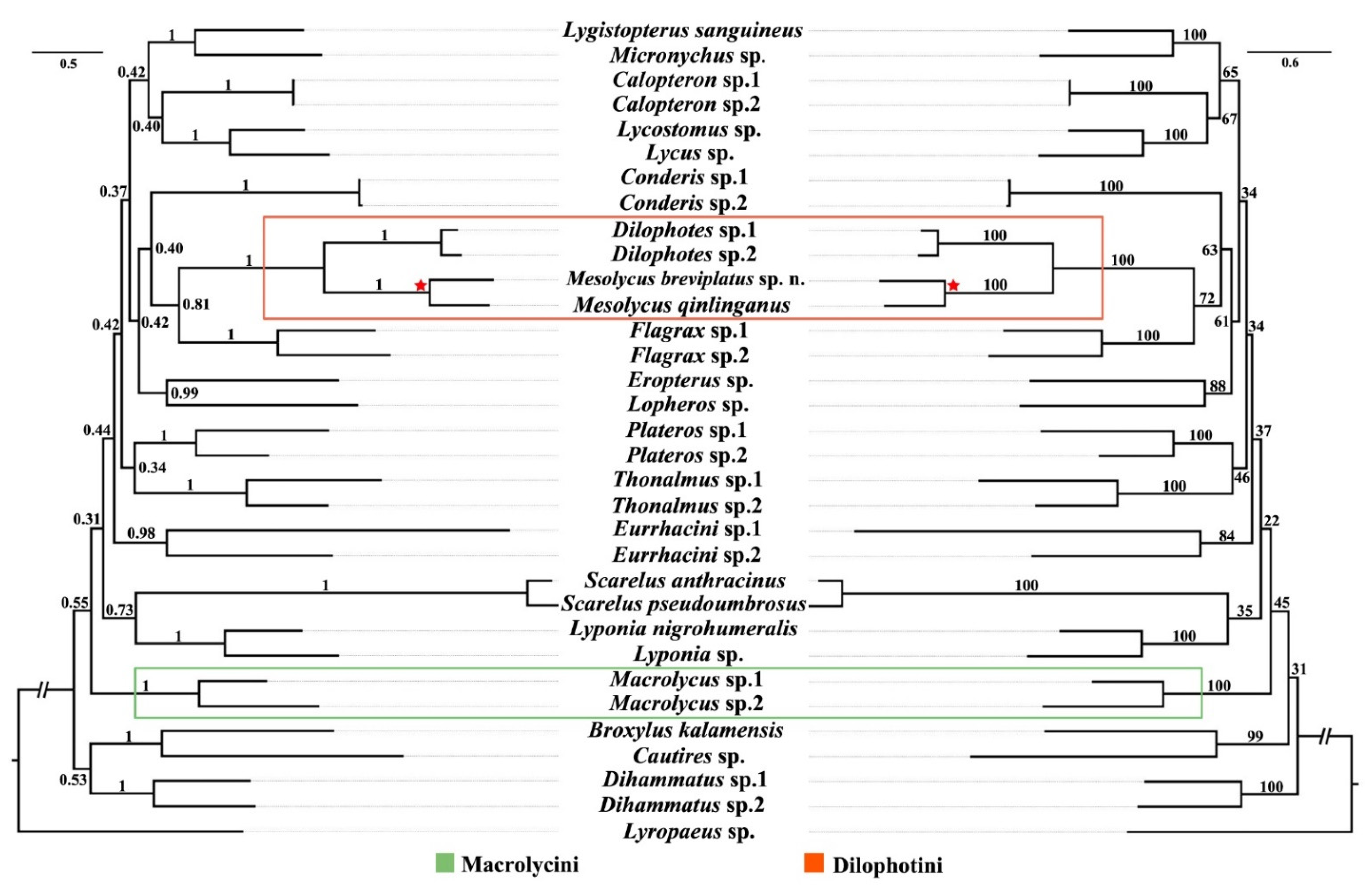
3.4. Phylogenetic Analyses
4. Discussion
4.1. Separate Status of the New Species
| Key to the Males of Mesolycus Species | |
| 2 | |
| 20 |
| 3 |
| 6 |
| M. mediozonatus (Nakane, 1955) |
| 4 |
| M. holzschuhi (Bic, 2002) |
| 5 |
| M. shaanxiensis sp. n. |
| M. hubeicus Kazantsev, 2004 |
| 7 |
| 11 |
| M. varus sp. n. |
| 8 |
| 9 |
| 10 |
| M. sausai (Bic, 2002) |
| M. bhutanensis (Bic, 2002) |
| M. vitalisi (Pic, 1923) |
| M. breviplatus sp. n. |
| 12 |
| 15 |
| 13 |
| 14 |
| M. murzini Kazantsev, 2004 |
| M. atrorufus (Kiesenwetter, 1879) |
| M. nanensis Kazantsev, 2004 |
| M. dentatus sp. n. |
| 16 |
| 17 |
| M. berezowskii (Kazantsev, 2000) |
| M. laosensis (Bic, 2002) |
| M. particularis (Pic, 1928) comb. n. |
| 18 |
| M. atricollis (Pic, 1926) comb. n. |
| 19 |
| M. qinlinganus (Kazantsev, 2000) |
| M. tibetanus (Kazantsev, 2000) |
| M. shelfordi (Bourgeois, 1906) |
| 21 |
| 22 |
| 26 |
| M. obscurus (Pic, 1912) |
| 23 |
| M. ilyai (Kazantsev, 2000) |
| 24 |
| M. pacholatkoi (Bic, 2002) comb. n. |
| 25 |
| M. fedorenkoi Kazantsev, 2013 |
| M. rubromarginatus Kazantsev, 2013 |
| M. discoidalis (Pic, 1912) |
| 27 |
| M. jendeki (Bic, 2002) |
| M. bolavensis(Bic, 2002) |
4.2. Species Diversity of Chinese Mesolycus
4.3. Phylogenetic Position of Mesolycus
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bocak, L.; Bocakova, M. Phylogeny and classification of the family Lycidae (Insecta: Coleoptera). Ann. Zool. 2008, 58, 695–720. [Google Scholar] [CrossRef]
- Motyka, M.; Kampova, L.; Bocak, L. Phylogeny and evolution of Müllerian mimicry in aposematic Dilophotes: Evidence foradvergence and size–constraintsin evolution of mimetic sexual dimorphism. Sci. Rep. 2018, 8, 3744. [Google Scholar] [CrossRef] [PubMed]
- Bocak, L.; Yagi, T. Evolution of mimicry patterns in Metriorrhynchus (Coleoptera: Lycidae): The history of dispersal and speciation in South East Asia. Evolution 2010, 64, 39–52. [Google Scholar] [CrossRef] [PubMed]
- Malohlava, V.; Bocak, L. Evidence of extreme habitat stability in a southeast Asian biodiversity hotspot based on the evolutionary analysis of neotenic net–winged beetles. Mol. Ecol. 2010, 19, 4800–4811. [Google Scholar] [CrossRef]
- Sklenarova, K.; Chesters, D.; Bocak, L. Phylogeography of poorly dispersing net–winged beetles: A role of drifting India in the origin of Afrotropical and Oriental fauna. PLoS ONE 2013, 8, e67957. [Google Scholar] [CrossRef] [Green Version]
- Nakane, T. Lycidae (Insecta: Coleoptera). In Fauna Japonica; Academic Press of Japan: Tokyo, Japan, 1969; 224 p. [Google Scholar]
- Bocak, L.; Matsuda, K. Review of the immature stages of the family Lycidae (Insetcta: Coleoptera). J. Nat. Hist. 2003, 37, 1463–1507. [Google Scholar] [CrossRef]
- Kazantsev, S.V. On Dilophotes Waterhouse, 1879 and Mimodilophotes Pic, 1939 (Coleoptera, Lycidae). Russ. Entomol. J. 2000, 9, 329–332. [Google Scholar]
- Kazantsev, S.V. Contribution to the knowledge of Macrolycini with description of Calcaeron, new genus (Coleoptera, Lycidae). Zootaxa 2004, 493, 1–32. [Google Scholar] [CrossRef]
- Bic, V. Review of the genus Dilophotes Waterhouse (Coleoptera: Lycidae) of the Palaearctic region and Indochina. Zootaxa 2002, 59, 1–26. [Google Scholar] [CrossRef]
- Li, Y.; Bocak, L.; Pang, H. New species of Macrolycus Waterhouse, 1878 from China and Laos, with a check–list of the genus (Coleoptera: Lycidae). Zootaxa 2012, 3232, 44–61. [Google Scholar] [CrossRef]
- Bocak, L.; Bocakova, M. Taxonomic notes on East–Asiatic Lycidae (Coleoptera). Acta Entomol. Bohemoslov. 1988, 85, 444–446. [Google Scholar]
- Kazantsev, S.V. New net–winged beetles (Coleoptera: Lycidae) from Indochina, with synonymic and taxonomic notes. Cauc. Entomol. Bull. 2013, 9, 247–252. [Google Scholar] [CrossRef]
- Kleine, R. Lycidae. In Coleopterorum Catalogus; Pars 123; Junk, W., Schenkling, S., Eds.; W. Junk: Berlin, Germany, 1933; 145 p. [Google Scholar]
- Kleine, R. Neue Indische Lycidae nebst faunistischen Bemerkungen. Indian For. Rec. 1928, 13, 221–268. [Google Scholar]
- Bocak, L.; Bocakova, M. Revision of the supergeneric classification of the family Lycidae (Coleoptera). Pol. Pismo Entomol. 1990, 59, 623–676. [Google Scholar]
- Kazantsev, S.V. Morphology of Lycidae with some considerations on evolution of the Coleoptera. Elytron 2005, 17, 49–226. [Google Scholar]
- Masek, M.; Motyka, M.; Kusy, D.; Bocek, M.; Li, Y.; Bocak, L. Molecular Phylogeny, Diversity and Zoogeography of Net–Winged Beetles (Coleoptera: Lycidae). Insects 2018, 9, 154. [Google Scholar] [CrossRef] [Green Version]
- Peng, Y.; Leung, H.C.; Yiu, S.M.; Chin, F.Y. IDBA–UD: A de novo assembler for single–cell and metagenomic sequencing data with highly uneven depth. Bioinformatics 2012, 28, 1420–1428. [Google Scholar] [CrossRef] [Green Version]
- Hebert, P.D.N.; Ratnasingham, S.; de Waard, J.R. Barcoding animal life: Cytochrome c oxidase subunit I divergences among closely related species. Proceedings of the Royal Society of London. Ser. B Biol. Sci. 2003, 270, 96–99. [Google Scholar] [CrossRef] [Green Version]
- Kearse, M.; Moir, R.; Wilson, A.; Stones–Havas, S.; Cheung, M.; Sturrock, S.; Buxyon, S.; Cooper, S.; Duran, C.; Thierer, T.; et al. Geneious Basic: An integrated and extendable desktop software platform for the organization and analysis of sequence data. Bioinformatics 2012, 28, 1647–1649. [Google Scholar] [CrossRef] [Green Version]
- Uribe, J.E.; Gutiérrez–Rodríguez, J. The complete mitogenome of the trilobite beetle, Platerodrilus sp. (Elateroidea: Lycidae). Mitochondrial DNA Part B 2016, 1, 658–659. [Google Scholar] [CrossRef] [Green Version]
- Bernt, M.; Donath, A.; Jühling, F.; Externbrink, F.; Florentz, C.; Fritzsch, G.; Pütz, J.; Middendorf, M.; Stadler, P.F. MITOS: Improved de novo metazoan mitochondrial genome annotation. Mol. Phylogenetics Evol. 2013, 69, 313–319. [Google Scholar] [CrossRef] [PubMed]
- Motyka, M.; Masek, M.; Bocak, L. Congruence between morphology and molecular phylogeny: The reclassification of Calochromini (Coleoptera: Lycidae) and their dispersal history. Zool. J. Linn. Soc. 2017, 180, 147–165. [Google Scholar] [CrossRef]
- Bocakova, M.; Bocak, L.; Hunt, T.; Teraväinen, M.; Vogler, A.P. Molecular phylogenetics of Elateriformia (Coleoptera): Evolution of bioluminescence and neoteny. Cladistics 2007, 23, 477–496. [Google Scholar] [CrossRef]
- Sklenarova, K.; Kubecek, V.; Bocak, L. Subtribal classification of Metriorrhynchini (Insecta, Coleoptera, Lycidae): An integrative approach using molecular phylogeny and morphology of adults and larvae. Arthropod Syst. Phylogeny 2014, 72, 37–54. [Google Scholar]
- Levkanicova, Z.; Bocak, L. Identification of net-winged beetle larvae (Coleoptera: Lycidae) using three mtDNA fragments: A comparison of their utility. Syst. Entomol. 2009, 34, 210–221. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Zhang, D.; Gao, F.; Jakovlić, I.; Zou, H.; Zhang, J.; Li, W.X.; Wang, G.T. PhyloSuite: An integrated and scalable desktop platform for streamlined molecular sequence data management and evolutionary phylogenetics studies. Mol. Ecol. Resour. 2020, 20, 348–355. [Google Scholar] [CrossRef]
- Capella–Gutiérrez, S.; Silla–Martínez, J.M.; Gabaldón, T. TrimAI: A tool for automated alignment trimming in large–scale phylogenetic analyses. Bioinformatics 2009, 25, 1972–1973. [Google Scholar] [CrossRef] [Green Version]
- Lanfear, R.; Calcott, B.; Ho, S.Y.; Guindon, S. PartitionFinder: Combined selection of partitioning schemes and substitution models for phylogenetic analyses. Mol. Biol. Evol. 2012, 29, 1695–1701. [Google Scholar] [CrossRef] [Green Version]
- Nguyen, L.T.; Schmidt, H.A.; Von–Haeseler, A.; Minh, B.Q. IQ–TREE: A fast and effective stochastic algorithm for estimating maximum–likelihood phylogenies. Mol. Biol. Evol. 2015, 32, 268–274. [Google Scholar] [CrossRef]
- Ronquist, F.; Huelsenbeck, J.P. MrBayes 3: Bayesian phylogenetic inference under mixed models. Bioinformatics 2003, 19, 1572–1574. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rambaut, A. FigTree v1.4.3 Software; Institute of Evolutionary Biology, University of Edinburgh: Edinburgh, UK, 2016. [Google Scholar]
- ICZN [International Commission on Zoological Nomenclature]. International Code of Zoological Nomenclature, 4th ed.; The International Trust for Zoological Nomenclature: London, UK, 1999. [Google Scholar]
- Kiesenwetter, H. Coleoptera Japoniae collecta a Domino Lewis et aliis. Dtsch. Entomol. Zaitschrift 1879, 23, 305–320. [Google Scholar] [CrossRef]
- Gorham, H.S. Revision of the genera and species of Malacoderm Coleoptera of the Japanese fauna. Trans. R. Entomol. Soc. Lond. 1883, 31, 393–411. [Google Scholar] [CrossRef]
- Kleine, R. Bestimmungstabelle der Lycidae. In Bestimmungstabellen der europäischen Coleopteren; 123 Heft; Edmund Reitter: Troppau, Czech Republic, 1942; 90p. [Google Scholar]
- Lewis, G. On the lamellicorn Coleoptera of Japan, and notices of others. Ann. Mag. Nat. Hist. 1895, 16, 374–408. [Google Scholar] [CrossRef] [Green Version]
- Bocakova, M.; Bocak, L. Lycidae. In Catalogue of Palaearctic Coleoptera; Elateroidea—Derodontoidea—Bostrichoidea—Lymexyloidea—Cleroidea—Cucujoidea; Löbl, I., Smetana, A., Eds.; Apollo Books: Stenstrup, Demarks, 2007; Volume 4, pp. 211–224. [Google Scholar]
- Matsuda, K. Identification manual of the lycid–beetles in Japan, IV. Sayabane N. S. 2012, 7, 1–6. [Google Scholar]
- Pic, M.H. Sauter’s Formosa Ausbeute. Mordellidae, Lycidae, Cantharidae. Entomol. Mitt. 1926, 15, 67–69. [Google Scholar]
- Pic, M. Nouveautés diverses. Mélanges Exot. Entomol. 1928, 52, 1–32. [Google Scholar]
- Li, D.; Aspöck, H.; Aspöck, U.; Liu, X.-Y. Mining the Species Diversity of Lacewings: New Species of the Pleasing Lacewing Genus Dilar Rambur, 1838 (Neuroptera, Dilaridae) from the Oriental Region. Insects 2021, 12, 451. [Google Scholar] [CrossRef]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, H.; Du, R.; Kang, Y.; Ge, X.; Yang, X.; Yang, Y. Unexpected High Species Diversity of Mesolycus Gorham (Coleoptera, Lycidae) from China, with a Preliminary Investigation on Its Phylogenetic Position Based on Multiple Genes. Insects 2022, 13, 1171. https://doi.org/10.3390/insects13121171
Liu H, Du R, Kang Y, Ge X, Yang X, Yang Y. Unexpected High Species Diversity of Mesolycus Gorham (Coleoptera, Lycidae) from China, with a Preliminary Investigation on Its Phylogenetic Position Based on Multiple Genes. Insects. 2022; 13(12):1171. https://doi.org/10.3390/insects13121171
Chicago/Turabian StyleLiu, Haoyu, Ruolan Du, Ya Kang, Xueying Ge, Xingke Yang, and Yuxia Yang. 2022. "Unexpected High Species Diversity of Mesolycus Gorham (Coleoptera, Lycidae) from China, with a Preliminary Investigation on Its Phylogenetic Position Based on Multiple Genes" Insects 13, no. 12: 1171. https://doi.org/10.3390/insects13121171