
Copper Exposure Affects Anti-Predatory Behaviour and Acetylcholinesterase Levels in Culex pipiens (Diptera, Culicidae)
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials & Methods
2.1. Prey Collection and Initial Rearing
2.2. Preparation of Copper Solutions
2.3. Predator Rearing and Preparation of Predator Cues and Predator Water
2.4. Experiment 1: Anti-Predatory Behaviour Tests
2.5. Experiment 2: Direct Predation Effect
2.6. Experiment 3: AChE Activity Assessment
2.7. Statistical Analysis
3. Results
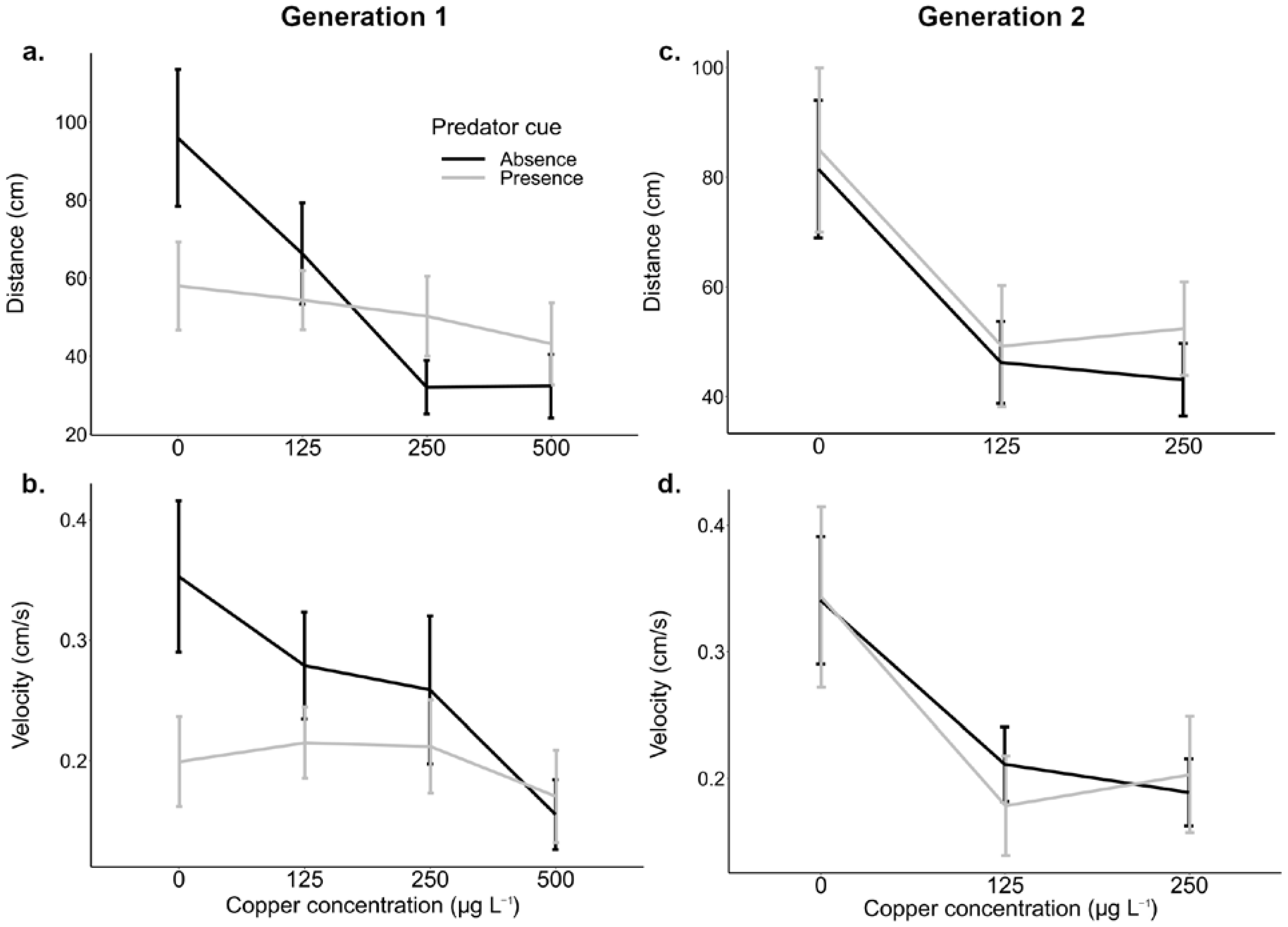
3.1. Effects of Copper and Predation Cues on Behaviour
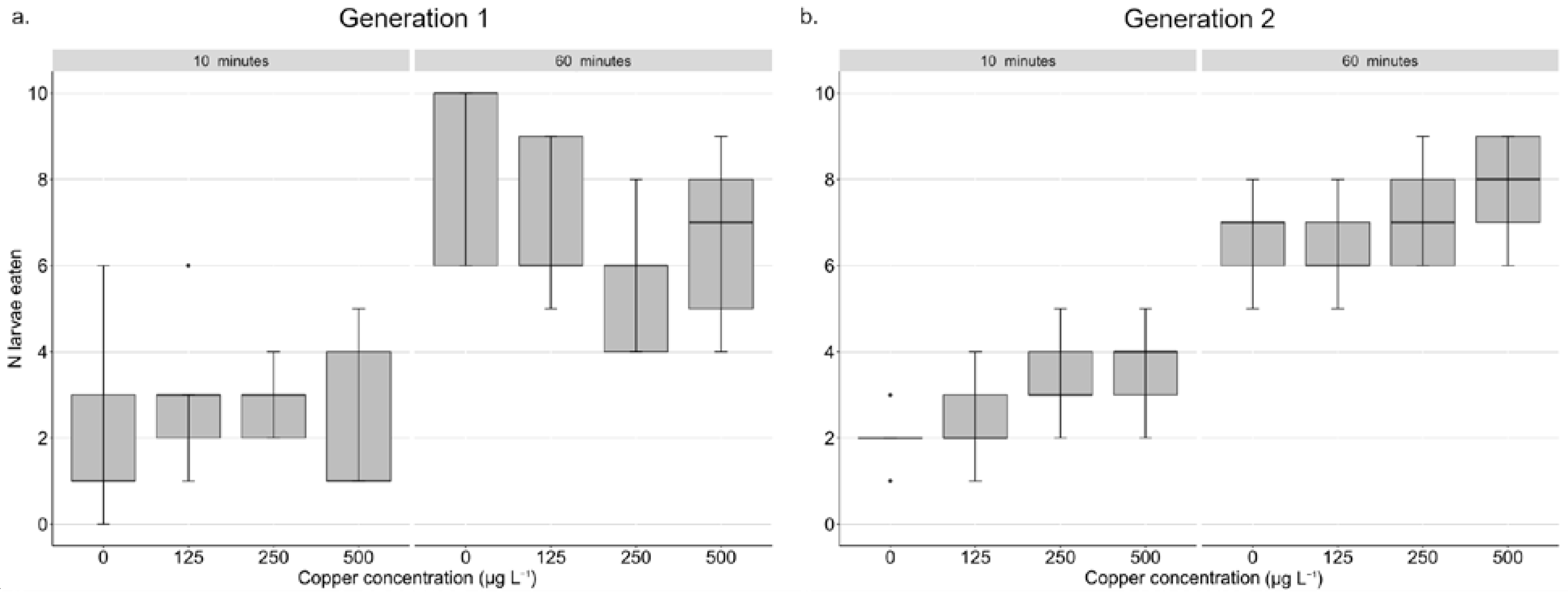
3.2. Effect of Copper on Escaping Behaviour
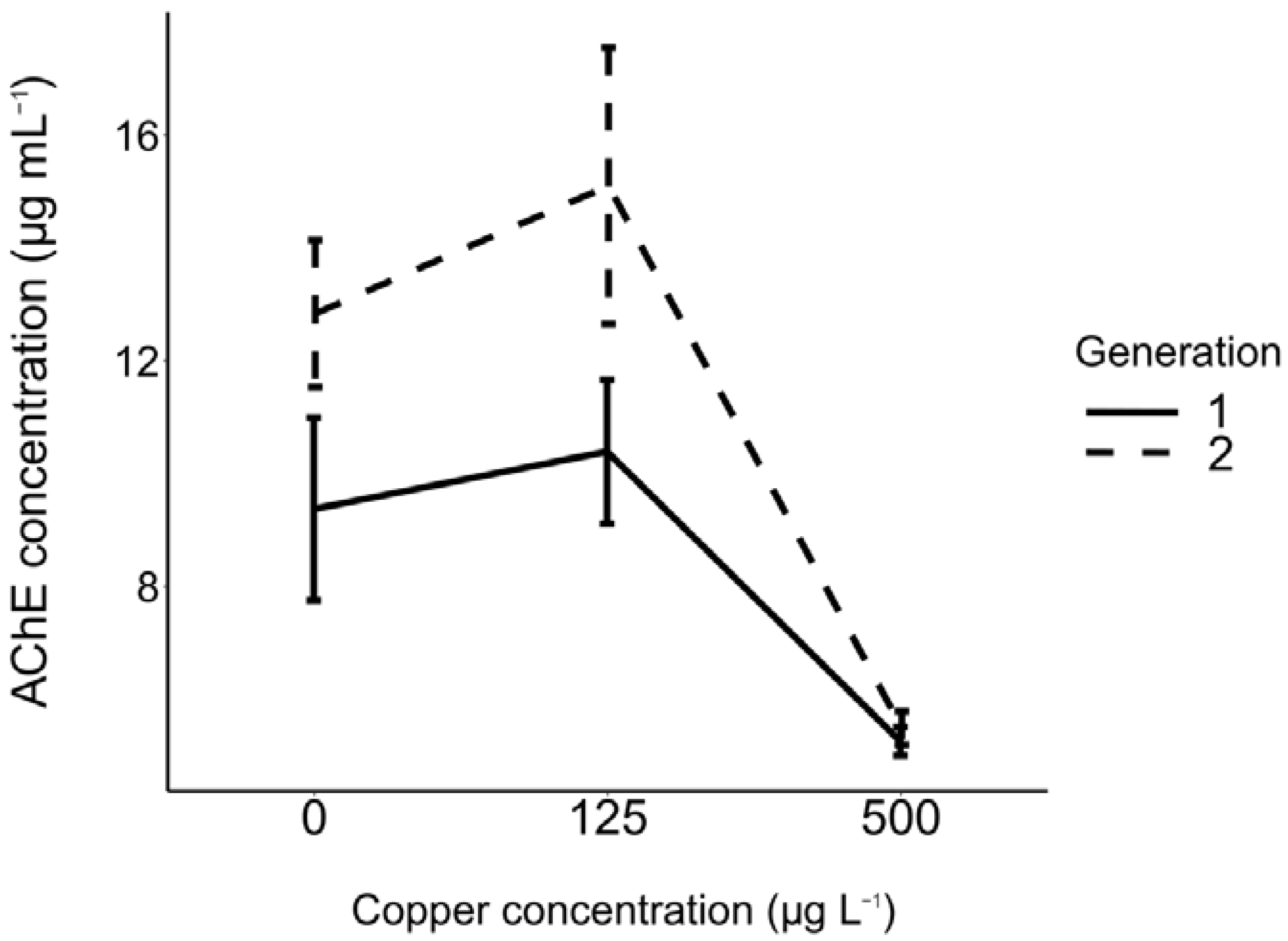
3.3. Effects of Copper on AChE Concentration
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Atafar, Z.; Mesdaghinia, A.; Nouri, J.; Homaee, M.; Yunesian, M.; Ahmadimoghaddam, M.; Mahvi, A.H. Effect of fertilizer application on soil heavy metal concentration. Environ. Monit. Assess. 2010, 160, 83–89. [Google Scholar] [CrossRef]
- Rajaganapathy, V.; Xavier, F.; Sreekumar, D.; Mandal, P.K. Heavy metal contamination in soil, water and fodder and their presence in livestock and products: A review. J. Environ. Sci. Technol. 2011, 4, 234–249. [Google Scholar] [CrossRef] [Green Version]
- Farombi, E.O.; Adelowo, O.A.; Ajimoko, Y.R. Biomarkers of oxidative stress and heavy metal levels as indicators of environmental pollution in African cat fish (Clarias gariepinus) from Nigeria Ogun River. Int. J. Environ. Res. Public Health 2007, 4, 158–165. [Google Scholar] [CrossRef] [Green Version]
- Benson, N.U.; Asuquo, F.E.; Williams, A.B.; Essien, J.P.; Ekong, C.I.; Akpabio, O.; Olajire, A.A. Source evaluation and trace metal contamination in benthic sediments from equatorial ecosystems using multivariate statistical techniques. PLoS ONE 2016, 11, 453–495. [Google Scholar] [CrossRef] [Green Version]
- Sildanchandra, W.; Crane, M. Influence of sexual dimorphism in Chironomus riparius Meigen on toxic effects of cadmium. Environ. Toxicol. Chem. An. Int. J. 2000, 19, 2309–2313. [Google Scholar] [CrossRef]
- Goswami, S.; Ahad, R.I.A.; Syiem, M.B. Expression of copper toxicity in the rice-field cyanobacterium anabaena oryzae ind4. Eur. Asian J. Biosci. 2019, 13, 57–67. [Google Scholar]
- Abdel-Tawwab, M.; Mousa, M.A.A.; Abbass, F.E. Growth performance and physiological response of African catfish, Clarias gariepinus (B.) fed organic selenium prior to the exposure to environmental copper toxicity. Aquaculture 2007, 272, 335–345. [Google Scholar] [CrossRef]
- U.S. EPA. Revised Lead and Copper Monitoring and Reporting Guidance for Public Water Systems; Office of Water, EPA 816-R-10-004; U.S. EPA: Washington, DC, USA, 2010.
- Brix, K.V.; DeForest, D.K.; Adams, W.J. The sensitivity of aquatic insects to divalent metals: A comparative analysis of laboratory and field data. Sci. Total Environ. 2011, 409, 4187–4197. [Google Scholar] [CrossRef]
- Clements, W.H.; Cadmus, P.; Brinkman, S.F. Responses of aquatic insects to Cu and Zn in stream microcosms: Understanding differences between single species tests and field responses. Environ. Sci. Technol. 2013, 47, 7506–7513. [Google Scholar] [CrossRef]
- Reza, M.; Ilmiawati, C. Laboratory testing of low concentration (<1 ppm) of copper to prolong mosquito pupation and adult emergence time: An alternative method to delay mosquito life cycle. PLoS ONE 2020, 15, e0226859. [Google Scholar] [CrossRef]
- Frasco, M.F.; Fournier, D.; Carvalho, F.; Guilhermino, L. Do metals inhibit acetylcholinesterase (AchE)? Implementation of assay conditions for the use of AchE activity as a biomarker of metal toxicity. Biomarkers 2005, 10, 360–375. [Google Scholar] [CrossRef]
- Boukadida, K.; Banni, M.; Romero-Ramirez, A.; Clerandeau, C.; Gourves, P.Y.; Cachot, J. Metal contamination and heat stress impair swimming behavior and acetylcholinesterase activity in embryo-larval stages of the Mediterranean mussel, Mytilus galloprovincialis. Mar. Environ. Res. 2022, 179, 105677. [Google Scholar] [CrossRef]
- Kim, Y.H.; Lee, S.H. Which acetylcholinesterase functions as the main catalytic enzyme in the Class Insecta? Insect Biochem. Mol. Biol. 2013, 43, 47–53. [Google Scholar] [CrossRef]
- Engdahl, C. Selective Inhibition of Acetylcholinesterase 1 from Disease-Transmitting Mosquitoes Design and Development of New Insecticides for Vector Control. Ph.D. Thesis, Umea University, Umea, Sweden, 2017. [Google Scholar]
- Rumrill, C.T.; Scott, D.E.; Lance, S.L. Effects of metal and predator stressors in larval southern toads (Anaxyrus terrestris). Ecotoxicology 2016, 25, 1278–1286. [Google Scholar] [CrossRef]
- Troncoso, L.; Galleguillos, R.; Larrain, A. Effects of copper on the fitness of the Chilean scallop Argopecten purpuratus (Mollusca: Bivalvia). Hydrobiologia 2000, 420, 185–189. [Google Scholar] [CrossRef]
- Mebane, C.A.; Schmidt, T.S.; Miller, J.L.; Balistrieri, L.S. Bioaccumulation and Toxicity of Cadmium, Copper, Nickel, and Zinc and Their Mixtures to Aquatic Insect Communities. Environ. Toxicol. Chem. 2020, 39, 812–833. [Google Scholar] [CrossRef] [Green Version]
- Haba, Y.; McBride, L. Origin and status of Culex pipiens mosquito ecotypes. Curr. Biol. 2022, 32, R237–R246. [Google Scholar] [CrossRef]
- Vinogradova, E.B. Culex Pipiens Pipiens Mosquitoes: Taxonomy, Distribution, Ecology, Physiology, Genetics, Applied Importance and Control; Pensoft Publishers: Sofia, Bulgaria, 2000. [Google Scholar]
- Turell, M.J.; O’Guinn, M.L.; Dohm, D.J.; Jones, J.W. Vector competence of North American mosquitoes (Diptera: Culicidae) for West Nile virus. J. Med. Entomol. 2001, 38, 130–134. [Google Scholar] [CrossRef]
- Gad, A.M.; Hammad, R.E.; Farid, H.A. Uptake and development of Wuchereria bancrofti in Culex pipiens L. and Aedes caspius pallas. J. Egypt Soc. Parasitol. 1996, 26, 305–314. [Google Scholar]
- Farid, H.A.; Hammad, R.E.; Hassan, M.M.; Morsy, Z.S.; Kamal, I.H.; Weil, G.J.; Ramzy, R.M. Detection of Wuchereria bancrofti in mosquitoes by the polymerase chain reaction: A potentially useful tool for large-scale control programmes. Trans. R. Soc. Trop. Med. Hyg. 2001, 95, 29–32. [Google Scholar] [CrossRef]
- Mackenzie, J.S.; Barrett, A.D.T.; Deubel, V. The Japanese Encephalitis Serological Group of Flaviviruses: A Brief Introduction to the Group. In Mackenzie JS, Barrett ADT, Deubel V; Encephalitis, J., Viruses, W.N., Eds.; Springer: Berlin/Heidelberg, Germany, 2002; pp. 1–10. [Google Scholar] [CrossRef] [Green Version]
- Kramer, L.D.; Ebel, G.D. Dynamics of flavivirus infection in mosquitoes. Adv. Virus. Res. 2003, 60, 187–232. [Google Scholar]
- Farajollahi, A.; Fonseca, D.M.; Kramer, L.D.; Marm Kilpatrick, A. “Bird biting” mosquitoes and human disease: A review of the role of Culex pipiens complex mosquitoes in epidemiology. Infect. Genet. Evol. 2011, 11, 1577–1585. [Google Scholar] [CrossRef]
- Ghorbani, F.; Vatandoost, H.; Hanafi-Bojd, A.A.; Abai, M.R.; Nikookar, H.; Enayati, A.A. High resistance of vector of West Nile Virus, Culex pipiens Linnaeus (Diptera: Culicidae) to different insecticides recommended by WHO in northern Iran. J. Arthropod Borne Dis. 2018, 12, 24–30. [Google Scholar]
- Al-Sarar, A.S. Insecticide resistance of Culex pipiens (L.) populations (Diptera: Culicidae) from Riyadh city, Saudi Arabia: Status and overcome. Saudi. J. Biol. Sci. 2010, 17, 95–100. [Google Scholar] [CrossRef] [Green Version]
- Reza, M.; Yamamoto, D.S.; Matsuoka, H. Low-concentration copper solution jeopardizes larval movement and ability to survive predation: New insight into malaria eradication via vector control. Med. Entomol. Zool. 2012, 63, 217–222. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, M.C.; Trowell, J.J.; Brown, G.E.; Chivers, D.P. The role of learning in the development of threat-sensitive predator avoidance by fathead minnows. Anim. Behav. 2005, 70, 777–784. [Google Scholar] [CrossRef]
- Kesavaraju, B.; Juliano, S.A. Differential behavioral responses to water-borne cues to predation in two container-dwelling mosquitoes. Ann. Entomol. Soc. Am. 2004, 97, 194–201. [Google Scholar] [CrossRef]
- Juliano, S.A.; Gravel, M.E. Predation and the evolution of prey behavior: An experiment with tree hole mosquitoes. Behav. Ecol. 2002, 13, 301–311. [Google Scholar] [CrossRef]
- Awasthi, A.K.; Molinero, J.C.; Wu, C.H.; Tsai, K.H.; King, C.C.; Hwang, J.S. Behavioral changes in mosquito larvae induced by copepods predation. Hydrobiologia 2015, 749, 113–123. [Google Scholar] [CrossRef]
- Ferrari, M.C.O.; Messier, F.; Chivers, D.P. Threat-sensitive learning of predators by larval mosquitoes Culex restuans. Behav. Ecol. Sociobiol. 2008, 62, 1079–1083. [Google Scholar] [CrossRef]
- Wisenden, B.D. Olfactory assessment of predation risk in the aquatic environment. Philos. Trans. R. Soc. B Biol. Sci. 2000, 355, 1205–1208. [Google Scholar] [CrossRef] [Green Version]
- Ferrari, M.C.O.; Wisenden, B.D.; Chivers, D.P. Chemical ecology of predator-prey interactions in aquatic ecosystems: A review and prospectus. Can. J. Zool. 2010, 88, 698–724. [Google Scholar] [CrossRef]
- Schoeppner, N.M.; Relyea, R.A. Interpreting the smells of predation: How alarm cues and kairomones induce different prey defences. Funct. Ecol. 2009, 23, 1114–1121. [Google Scholar] [CrossRef]
- Weber, C.I. Methods for Measuring the Acute Toxicity of Effluents and Receiving Waters to Freshwater and Marine Organisms; Environmental Monitoring Systems Laboratory: Cincinnati, OH, USA, 1991. [Google Scholar]
- Potera, C. Fullerenes and Fish Brains Copper in Drinking Water Lead in Mexican Children Pottery Use Slows Reductions in Blood. Environ. Health. Perspect 2004, 112, 568–569. [Google Scholar]
- Houmann, C. Functional Characterization of Green Sorption Media and Scaling of Pilot Studies for Copper Removal in Stormwater Runoff. Master’s Thesis, University of Central Florida, Orlando, FL, USA, 2015. [Google Scholar]
- Magnusson, A.; Skaug, H.; Nielsen, A.; Berg, C.; Kristensen, K.; Maechler, M.; Skaug, H.; Nielsen, A.; Berg, C.; van Bentham, K.; et al. glmmTMB: Generalized Linear Mixed Models Using Template Model Builder. R Package Version 0.1. 3. Cran R Proj. 2017. Available online: https://rdrr.io/cran/glmmTMB/ (accessed on 11 December 2017).
- Weisberg, S.; Fox, J. An R Companion to Applied Regression; Sage Publications: Thousand Oaks, CA, USA, 2010. [Google Scholar]
- Russell, L. Emmeans: Estimated Marginal Means, Aka Least-Squares Means; R Package Version 1.4. 3.01; The University of Iowa Iowa City: Iowa City, IA, USA, 2019. [Google Scholar]
- Fox, J.; Monette, G.; Chalmers, P. Package ‘Heplots’ R Topics Documented: 2022. Available online: https://mirror.las.iastate.edu/CRAN/web/packages/heplots/heplots.pdf (accessed on 20 October 2022).
- Amer, N.R.; Lawler, S.P.; Zohdy, N.M.; Younes, A.; ElSayed, W.M.; Connon, R.E. Effect of long-term exposure to copper on survival and development of two successive generations of Culex pipiens (Diptera, Culicidae). Ecotoxicology 2021, 30, 351–360. [Google Scholar] [CrossRef]
- Simbeya, C.K.; Csuzdi, C.E.; Dew, W.A.; Pyle, G.G. Electroantennogram measurement of the olfactory response of Daphnia spp. and its impairment by waterborne copper. Ecotoxicol. Environ. Saf. 2012, 82, 80–84. [Google Scholar] [CrossRef]
- Thomas, O.R.B.; Barbee, N.C.; Hassell, K.L.; Swearer, S.E. Smell no evil: Copper disrupts the alarm chemical response in a diadromous fish, Galaxias maculatus. Environ. Toxicol. Chem. 2016, 35, 2209–2214. [Google Scholar] [CrossRef]
- Wisenden, B.D.; Pollock, M.S.; Tremaine, R.J.; Webb, J.M.; Wismer, M.E.; Chivers, D.P. Synergistic interactions between chemical alarm cues and the presence of conspecific and heterospecific fish shoals. Behav. Ecol. Sociobiol. 2003, 54, 485–490. [Google Scholar] [CrossRef]
- Roberts, D. Mosquito larvae change their feeding behavior in response to kairomones from some predators. J. Med. Entomol. 2014, 51, 368–374. [Google Scholar] [CrossRef] [Green Version]
- Roux, O.; Diabaté, A.; Simard, F. Larvae of cryptic species of Anopheles gambiae respond differently to cues of predation risk. Freshw. Biol. 2013, 58, 1178–1189. [Google Scholar] [CrossRef]
- Mireji, P.O.; Keating, J.; Hassanali, A.; Mbogo, C.M.; Muturi, M.N.; Githure, J.I.; Beier, J.C. Biological cost of tolerance to heavy metals in the mosquito Anopheles gambiae. Med. Vet. Entomol. 2010, 24, 101–107. [Google Scholar] [CrossRef] [Green Version]
- Bonnard, M.; Roméo, M.; Amiard-Triquet, C. Effects of copper on the burrowing behavior of estuarine and coastal invertebrates, the polychaete Nereis diversicolor and the bivalve Scrobicularia plana. Hum. Ecol. Risk Assess. 2009, 15, 11–26. [Google Scholar] [CrossRef]
- McIntyre, J.K.; Baldwin, D.H.; Beauchamp, D.A.; Scholz, N.L. Low-level copper exposures increase visibility and vulnerability of juvenile coho salmon to cutthroat trout predators. Ecol. Appl. 2012, 22, 1460–1471. [Google Scholar] [CrossRef]
- Clements, W.H.; Cherry, D.S.; Cairns, J. The influence of copper exposure on predator-prey interactions in aquatic insect communities. Freshw. Biol. 1989, 21, 483–488. [Google Scholar] [CrossRef]
- Baldwin, D.H.; Tatara, C.P.; Scholz, N.L. Copper-induced olfactory toxicity in salmon and steelhead: Extrapolation across species and rearing environments. Aquat. Toxicol. 2011, 101, 295–297. [Google Scholar] [CrossRef]
- Green, W.W.; Mirza, R.S.; Wood, C.; Pyle, G.G. Copper binding dynamics and olfactory impairment in fathead minnows (Pimephales promelas). Environ. Sci. Technol. 2010, 44, 1431–1437. [Google Scholar] [CrossRef]
- Sandahl, J.F.; Baldwin, D.H.; Jenkins, J.J.; Scholz, N.L. A sensory system at the interface between urban stormwater runoff and salmon survival. Environ. Sci. Technol. 2007, 41, 2998–3004. [Google Scholar] [CrossRef]
- Hayden, M.T.; Reeves, M.K.; Holyoak, M.; Perdue, M.; King, A.L.; Tobin, S.C. Thrice as easy to catch! Copper and temperature modulate predator-prey interactions in larval dragonflies and anurans. Ecosphere 2015, 6, 1–17. [Google Scholar] [CrossRef] [Green Version]
- Lacoursière, J.O.; Dahl, C.; Widahl, L.E. Use of the continuity principle to evaluate water processing rate of suspension-feeding mosquito larvae. J. Am. Mosq. Control Assoc. 1999, 15, 228–237. [Google Scholar]
- Tilton, F.A.; Bammler, T.K.; Gallagher, E.P. Swimming impairment and acetylcholinesterase inhibition in zebrafish exposed to copper or chlorpyrifos separately, or as mixtures. Comp. Biochem. Physiol. C Toxicol. Pharmacol. 2011, 153, 9–16. [Google Scholar] [CrossRef] [Green Version]
- Carlier, P.R.; Anderson, T.D.; Wong, D.M.; Hsu, D.C.; Hartsel, J.; Ma, M.; Wong, E.A.; Choudhury, R.; Lam, P.C.-H.; Totrov, M.M.; et al. Towards a species-selective acetylcholinesterase inhibitor to control the mosquito vector of malaria, Anopheles gambiae. Chem. Biol. Interact. 2008, 175, 368–375. [Google Scholar] [CrossRef]
- Hamza-Chaffai, A.; Roméo, M.; Gnassia-Barelli, M.; El Abed, A. Effect of Copper and Lindane on Some Biomarkers Measured in the Clam Ruditapes decussatus. Bull Environ. Contam. Toxicol. 1998, 61, 397–404. [Google Scholar] [CrossRef]
- Buchwalter, D.B.; Cain, D.J.; Clements, W.H.; Luoma, S.N. Using biodynamic models to reconcile differences between laboratory toxicity tests and field biomonitoring with aquatic insects. Environ. Sci. Technol. 2008, 42, 3117. [Google Scholar] [CrossRef] [Green Version]
- Tunçsoy, M.; Erdem, C. Copper Accumulation in Tissues of Oreochromis niloticus Exposed to Copper Oxide Nanoparticles and Copper Sulphate with Their Effect on Antioxidant Enzyme Activities in Liver. Water Air Soil Pollut. 2018, 229, 269. [Google Scholar] [CrossRef]
- Araya, M.; Olivares, M.; Pizarro, F. Copper in human health. Int. J. Environ. Health 2007, 1, 608–620. [Google Scholar] [CrossRef]
- Mastin, B.J.; Rodgers, J.H. Toxicity and bioavailability of copper herbicides (clearigate, cutrine-plus, and copper sulfate) to freshwater animals. Arch. Environ. Contam. Toxicol. 2000, 39, 445–451. [Google Scholar] [CrossRef]
- Stauber, J.L.; Davies, C.M. Use and limitations of microbial bioassays for assessing copper bioavailability in the aquatic environment. Environ. Rev. 2000, 8, 255–301. [Google Scholar] [CrossRef]
- De Oliveira-Filho, E.C.; Matos Lopes, R.; Roma Paumgartten, F.J. Comparative study on the susceptibility of freshwater species to copper-based pesticides. Chemosphere 2004, 56, 369–374. [Google Scholar] [CrossRef] [Green Version]
- Becker, N.; Oo, T.T.; Schork, N. Metallic copper spray—A new control technique to combat invasive container-inhabiting mosquitoes. Parasites Vectors 2015, 8, 575. [Google Scholar] [CrossRef] [Green Version]
- O’Meara, G.F.; Evans, L.F.; Gettman, A.D. Reduced mosquito production in cemetery vases with copper liners. J. Am. Mosq. Control Assoc. 1992, 8, 419–420. Available online: http://europepmc.org/abstract/MED/1361942 (accessed on 1 December 1992).
- Minal, S.P.; Prakash, S. Cu-Zn and Ag-Cu bimetallic nanoparticles as larvicide to control malaria parasite vector: A comparative analysis. In Proceedings of the IEEE Reg 10 Humanit Technol Conf 2016, R10-HTC 2016-Proc, Agra, India, 24 April 2017. [Google Scholar] [CrossRef]
- Sharon, E.A.; Velayutham, K.; Ramanibai, R. Biosynthesis of Copper Nanoparticles using Artocarpus heterophyllus against Dengue Vector Aedes aegypti. Int. J. Life-Sci. Sci Res. 2018, 4, 1872–1879. [Google Scholar] [CrossRef]
- Mogren, C.L.; Trumble, J.T. The impacts of metals and metalloids on insect behavior. Entomol. Exp. Appl. 2010, 135, 1–17. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Generation 1 | Generation 2 | |||||
|---|---|---|---|---|---|---|
| Distance | Velocity | Distance | Velocity | |||
| Variables | Df | p | p | Df | p | p |
| Length of larvae | 1 | <0.001 (23.0) *** | <0.001 (17.3) *** | 1 | <0.001 (56.4) *** | <0.001 (50.2) *** |
| Copper concentration | 3 | 0.001 (15.4) ** | 0.002 (15.0) ** | 2 | <0.001 (60.6) *** | <0.001 (53.7) *** |
| Predator cue | 1 | 0.595 (0.28) | 0.027 (4.87) * | 1 | 0.797 (0.07) | 0.134 (2.24) |
| Copper concentration × predator cue | 3 | 0.007 (12.0) ** | 0.140 (5.48) | 2 | 0.382 (1.92) | 0.562 (1.15) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Amer, N.R.; Lawler, S.P.; Zohdy, N.M.; Younes, A.; ElSayed, W.M.; Wos, G.; Abdelrazek, S.; Omer, H.; Connon, R.E. Copper Exposure Affects Anti-Predatory Behaviour and Acetylcholinesterase Levels in Culex pipiens (Diptera, Culicidae). Insects 2022, 13, 1151. https://doi.org/10.3390/insects13121151
Amer NR, Lawler SP, Zohdy NM, Younes A, ElSayed WM, Wos G, Abdelrazek S, Omer H, Connon RE. Copper Exposure Affects Anti-Predatory Behaviour and Acetylcholinesterase Levels in Culex pipiens (Diptera, Culicidae). Insects. 2022; 13(12):1151. https://doi.org/10.3390/insects13121151
Chicago/Turabian StyleAmer, Nermeen R., Sharon P. Lawler, Nawal M. Zohdy, Aly Younes, Wael M. ElSayed, Guillaume Wos, Samah Abdelrazek, Hind Omer, and Richard E. Connon. 2022. "Copper Exposure Affects Anti-Predatory Behaviour and Acetylcholinesterase Levels in Culex pipiens (Diptera, Culicidae)" Insects 13, no. 12: 1151. https://doi.org/10.3390/insects13121151