
Evaluation of Modern Techniques for Species Identification of Lutzia Mosquitoes (Diptera: Culicidae) in Thailand: Geometric Morphometrics and DNA Barcoding

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
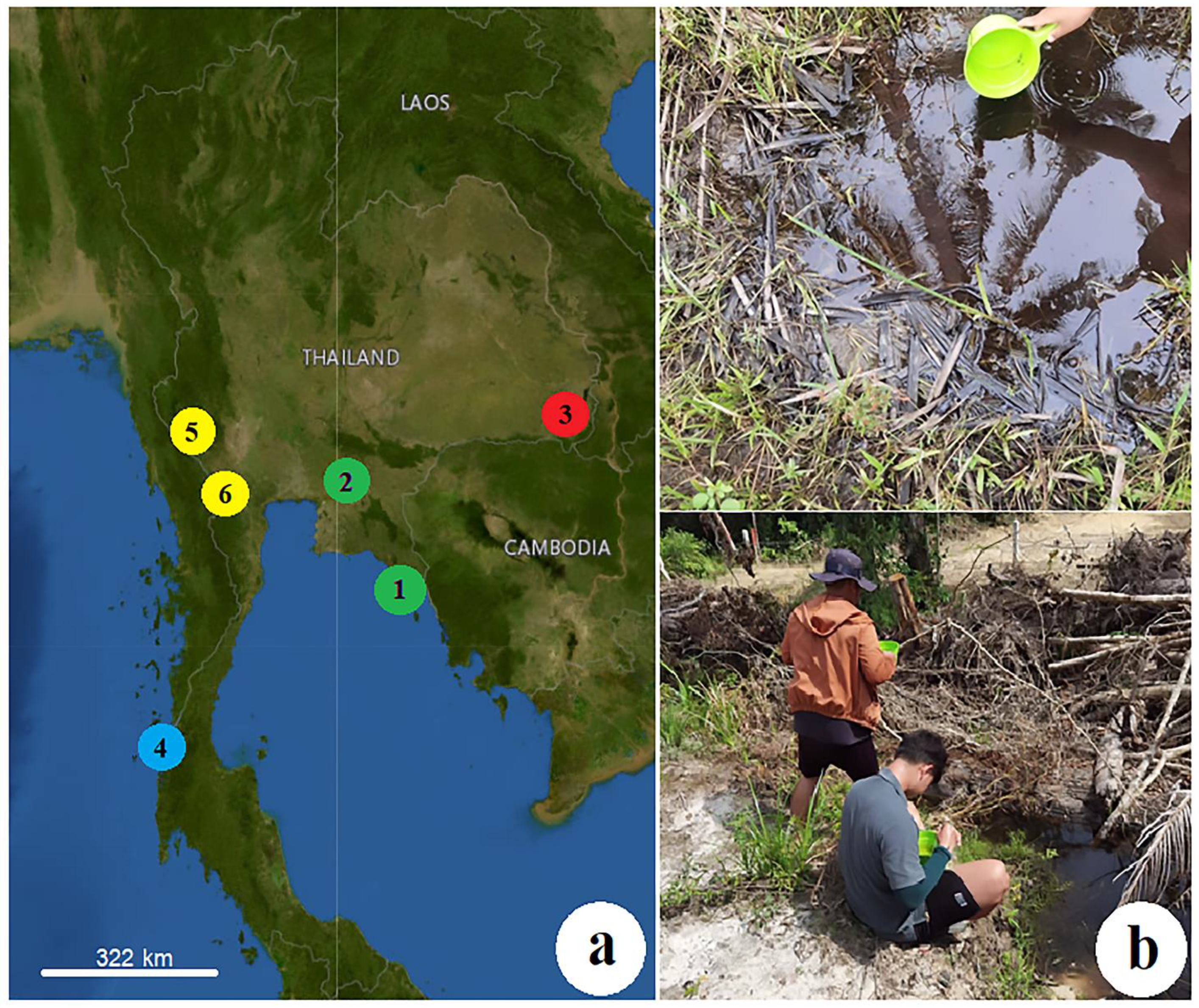
2.1. Mosquito Collection
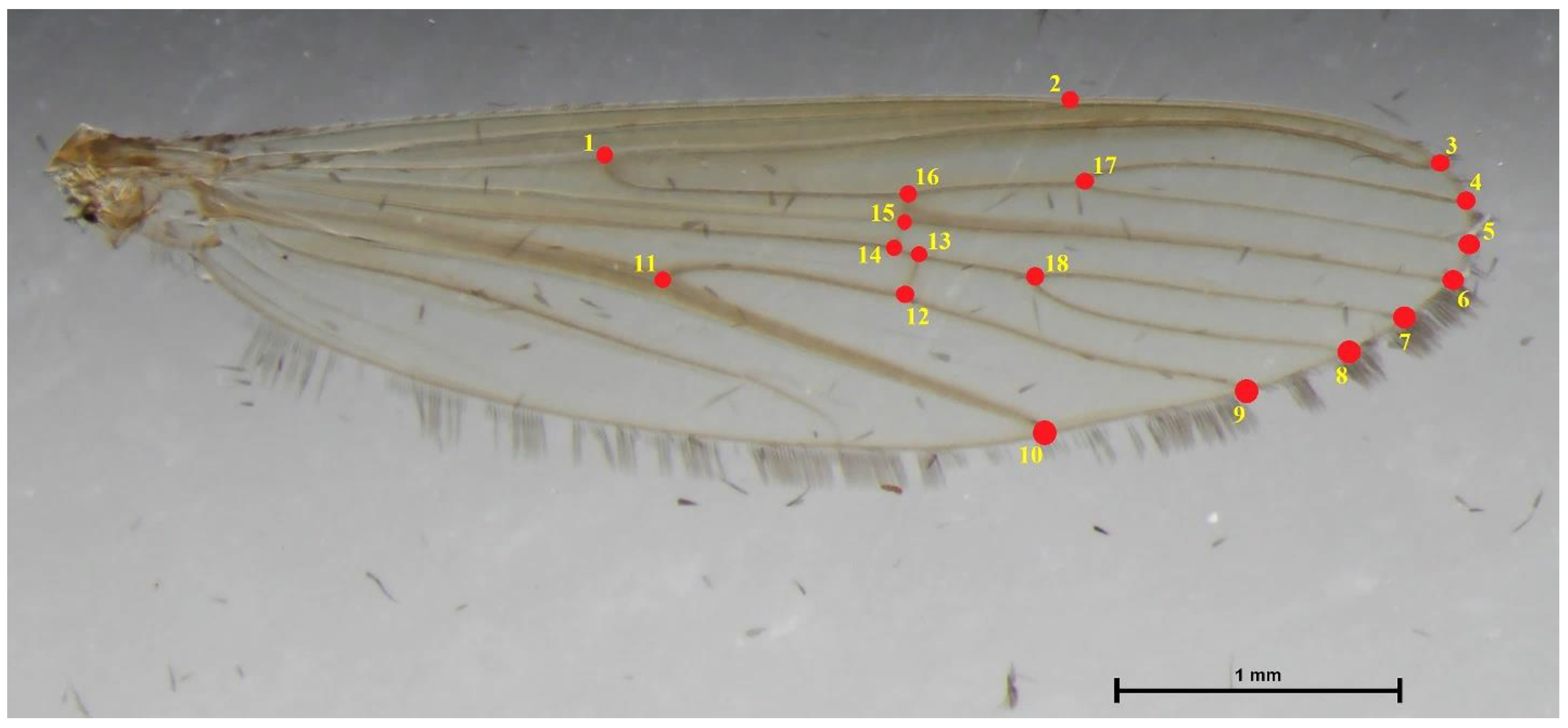
2.2. Geometric Morphometrics
2.3. DNA Extraction, PCR Amplification, and DNA Sequencing
2.4. Software
3. Results
3.1. Lutzia Species
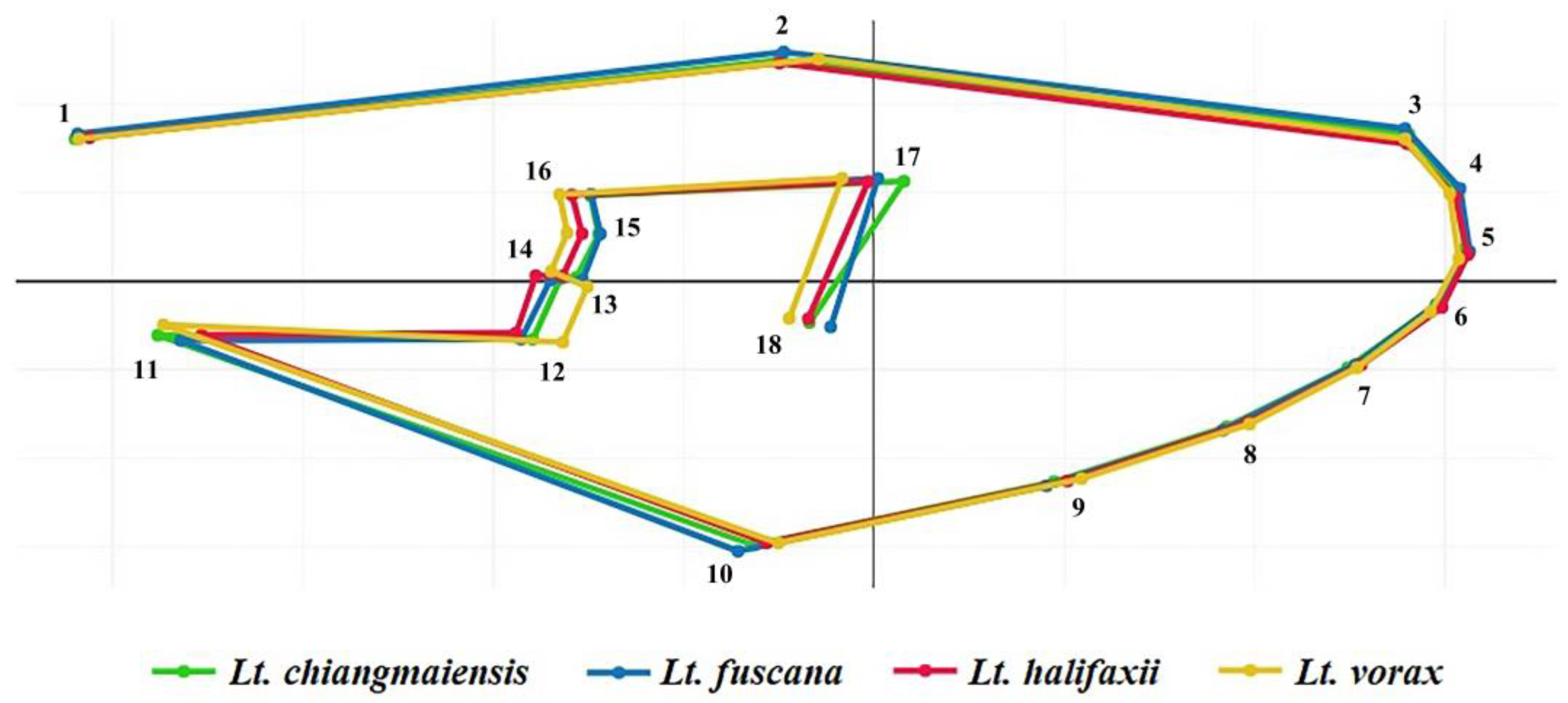
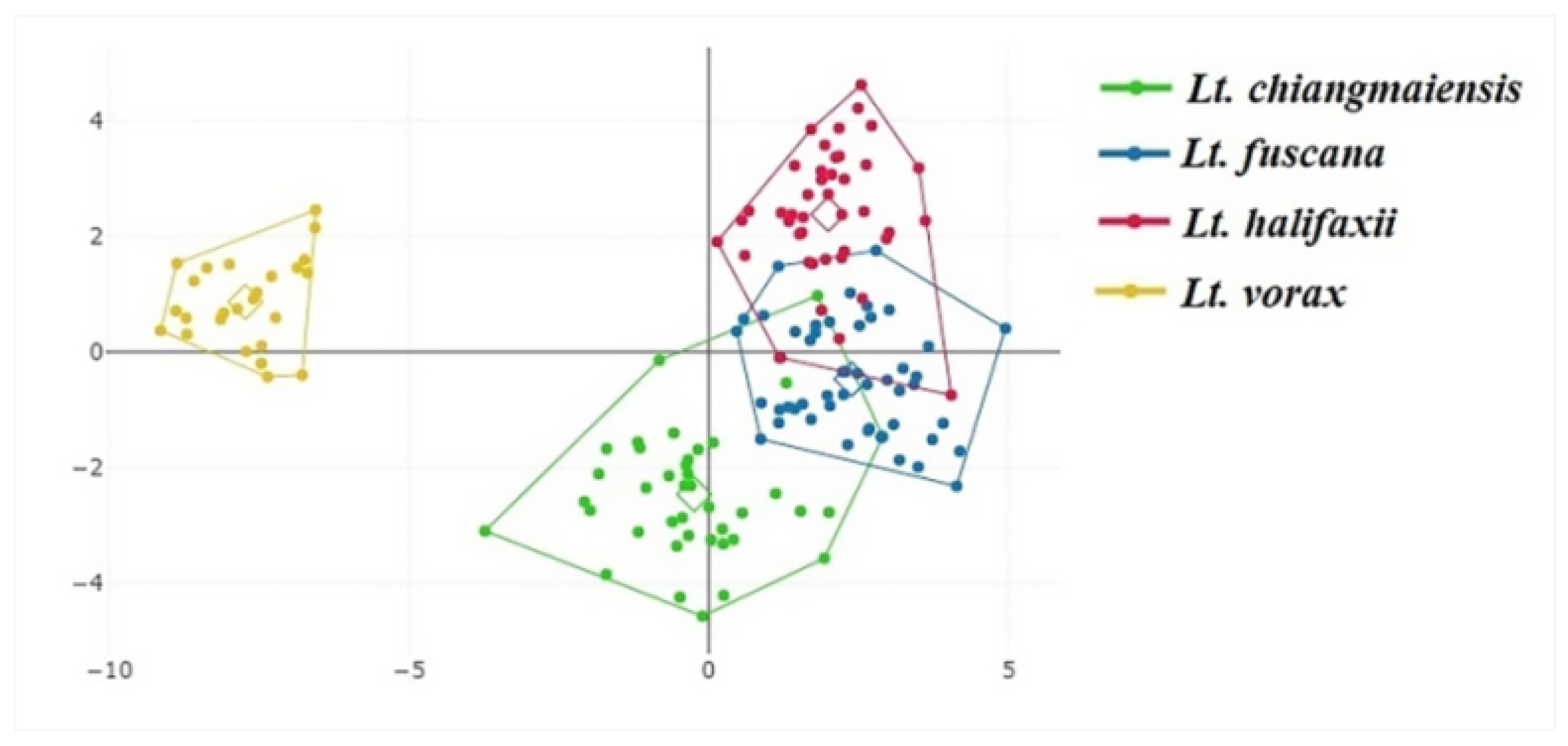
3.2. Wing Geometric Morphometrics
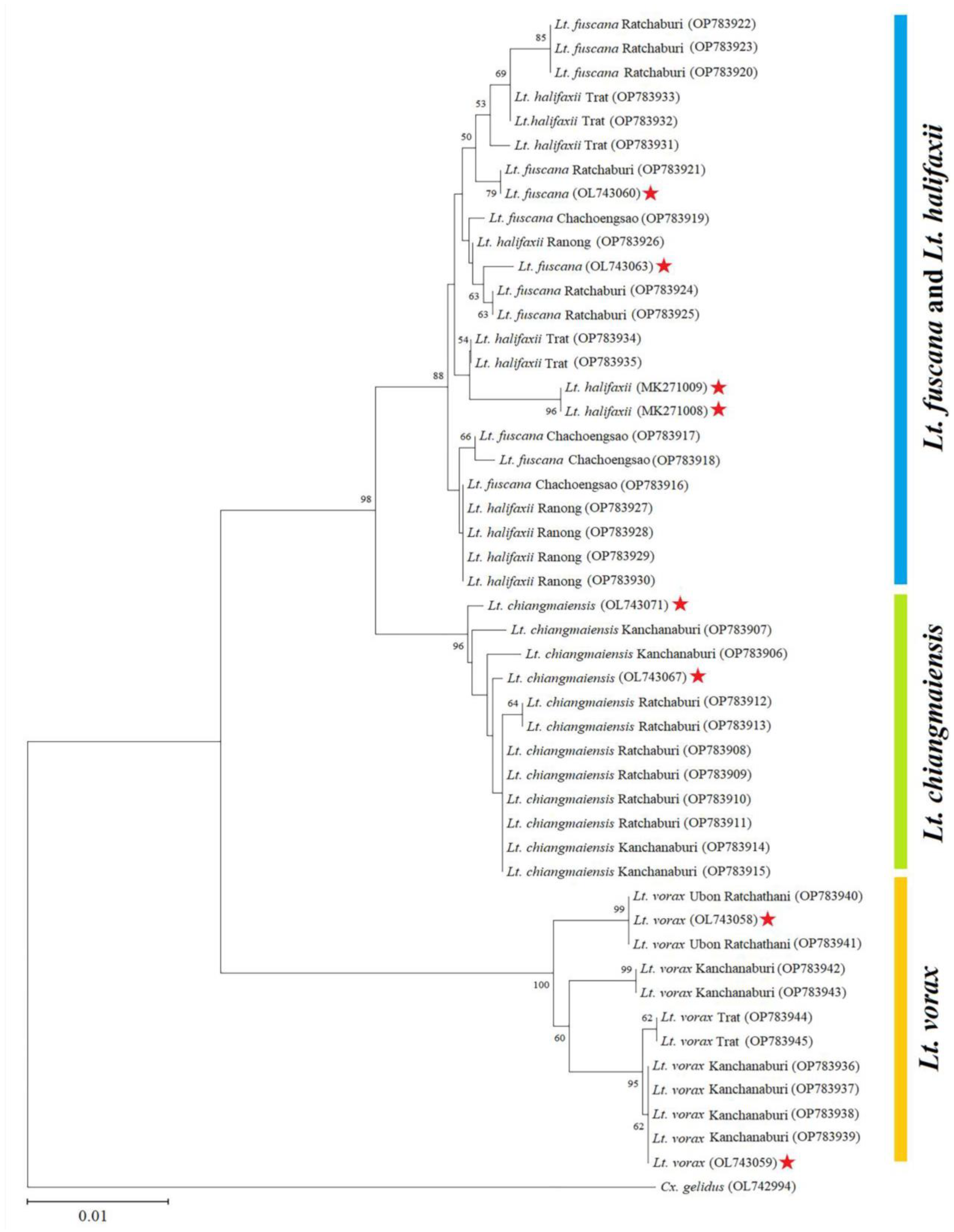
3.3. Barcode Sequences
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Harbach, R.E. Mosquito Taxonomic Inventory. Available online: https://mosquito-taxonomic-inventory.myspecies.info/simpletaxonomy/term/6192 (accessed on 8 October 2022).
- Harbach, R.E.; Kitching, I.J.; Culverwell, C.L.; Dubois, J.; Linton, Y.M. Phylogeny of Mosquitoes of Tribe Culicini (Diptera: Culicidae) Based on Morphological Diversity. Zool. Scr. 2012, 41, 499–514. [Google Scholar] [CrossRef]
- Chaiphongpachara, T.; Changbunjong, T.; Laojun, S.; Nutepsu, T.; Suwandittakul, N.; Kuntawong, K.; Sumruayphol, S.; Ruangsittichai, J. Mitochondrial DNA Barcoding of Mosquito Species (Diptera: Culicidae) in Thailand. PLoS ONE 2022, 17, e0275090. [Google Scholar] [CrossRef] [PubMed]
- Phanitchakun, T.; Namgay, R.; Miyagi, I.; Tsuda, Y.; Walton, C.; Harbach, R.E.; Somboon, P. Morphological and Molecular Evidence for a New Species of Lutzia (Diptera: Culicidae: Culicini) from Thailand. Acta Trop. 2019, 191, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Rattanarithikul, R.; Harbach, R.E.; Harrison, B.A.; Panthusiri, P.; Jones, J.W.; Coleman, R.E. Illustrated Keys to the Mosquitoes of Thailand. II. Genera Culex and Lutzia. Southeast Asian J. Trop. Med. Public Health 2005, 36, 2. [Google Scholar]
- Surendran, S.N.; Jude, P.J.; Thavaranjit, A.C.; Eswaramohan, T.; Vinobaba, M.; Ramasamy, R. Predatory Efficacy of Culex (Lutzia) fuscanus on Mosquito Vectors of Human Diseases in Sri Lanka. J. Am. Mosq. Control Assoc. 2013, 29, 168–170. [Google Scholar] [CrossRef]
- Somboon, P.; Harbach, R.E. Lutzia (Metalutzia) Chiangmaiensis n. sp. (Diptera: Culicidae), Formal Name for the Chiang Mai (CM) Form of the Genus Lutzia in Thailand. J. Med. Entomol. 2019, 56, 1270–1274. [Google Scholar] [CrossRef]
- Sumruayphol, S.; Apiwathnasorn, C.; Ruangsittichai, J.; Sriwichai, P.; Attrapadung, S.; Samung, Y.; Dujardin, J.P. DNA Barcoding and Wing Morphometrics to Distinguish Three Aedes Vectors in Thailand. Acta Trop. 2016, 159, 1–10. [Google Scholar] [CrossRef]
- Sumruayphol, S.; Chaiphongpachara, T.; Samung, Y.; Ruangsittichai, J.; Cui, L.; Zhong, D.; Sattabongkot, J.; Sriwichai, P. Seasonal Dynamics and Molecular Differentiation of Three Natural Anopheles Species (Diptera: Culicidae) of the Maculatus Group (Neocellia Series) in Malaria Hotspot Villages of Thailand. Parasit Vectors 2020, 13, 574. [Google Scholar] [CrossRef]
- Oliveira-Christe, R.; Marrelli, M.T. Using Geometric Morphometric Analysis of Wings to Identify Mosquito Species from the Subgenus Microculex (Diptera: Culicidae). J. Vector Ecol. 2021, 46, 221–225. [Google Scholar] [CrossRef]
- de Souza, A.L.D.S.; Multini, L.C.; Marrelli, M.T.; Wilke, A.B.B. Wing Geometric Morphometrics for Identification of Mosquito Species (Diptera: Culicidae) of Neglected Epidemiological Importance. Acta Trop. 2020, 211, 105593. [Google Scholar] [CrossRef]
- Dujardin, J.P. Morphometrics Applied to Medical Entomology. Infect. Genet. Evol. 2008, 8, 875–890. [Google Scholar] [CrossRef] [PubMed]
- Simões, R.F.; Wilke, A.B.B.; Chagas, C.R.F.; de Menezes, R.M.T.; Suesdek, L.; Multini, L.C.; Silva, F.S.; Grech, M.G.; Marrelli, M.T.; Kirchgatter, K. Wing Geometric Morphometrics as a Tool for the Identification of Culex Subgenus Mosquitoes of Culex (Diptera: Culicidae). Insects 2020, 11, 567. [Google Scholar] [CrossRef] [PubMed]
- Bellin, N.; Calzolari, M.; Callegari, E.; Bonilauri, P.; Grisendi, A.; Dottori, M.; Rossi, V. Geometric Morphometrics and Machine Learning as Tools for the Identification of Sibling Mosquito Species of the Maculipennis Complex (Anopheles). Infect. Genet. Evol. 2021, 95, 105034. [Google Scholar] [CrossRef]
- Boumaza, M.; Merabti, B.; Adjami, Y.; Ouakid, M.L. Geometric Morphometric Wing Analysis of Avian Malaria Vector, Culiseta longiareolata, from Two Locations in Algeria. Insects 2022, 13, 1031. [Google Scholar] [CrossRef] [PubMed]
- Petersen, V.; Santana, M.; Alves, J.M.P.; Suesdek, L. Genetic and Morphological Polymorphisms of Aedes scapularis (Diptera: Culicidae), Vector of Filariae and Arboviruses. Infect. Genet. Evol. 2022, 97, 105193. [Google Scholar] [CrossRef]
- DE Mello, C.F.; Carbajal-DE-LA-Fuente, A.L.; Ferreira, V.D.C.; Alencar, J. Influence of Seasonality on Wing Morphological Variability in Populations of Mansonia amazonensis (Theobald) (Diptera: Culicidae). Zootaxa 2022, 5100, 269–279. [Google Scholar] [CrossRef]
- Martinet, J.P.; Ferté, H.; Sientzoff, P.; Krupa, E.; Mathieu, B.; Depaquit, J. Wing Morphometrics of Aedes Mosquitoes from North-Eastern France. Insects 2021, 12, 341. [Google Scholar] [CrossRef]
- Demirci, B.; Bedir, H.; Akiner, M.M. Landmark-Based Geometric Morphometric Analysis of Wing Size and Wing Shape among Aedes albopictus (Skuse, 1894) Populations in Turkey. J. Vector Ecol. 2021, 46, 103–111. [Google Scholar] [CrossRef]
- Morales Vargas, R.; Tsunoda, T.; Noda, J.; Bousses, P.; Nguyen, T.Y.; Hasebe, F.; Dujardin, J.P. Shape Relatedness between Geographic Populations of Culex tritaeniorhynchus, the Primary Vector of Japanese Encephalitis Virus: A Landmark Study. Infect. Genet. Evol. 2021, 90, 104764. [Google Scholar] [CrossRef]
- Carvajal, T.M.; Amalin, D.M.; Watanabe, K. Wing Geometry and Genetic Analyses Reveal Contrasting Spatial Structures between Male and Female Aedes aegypti (L.) (Diptera: Culicidae) Populations in Metropolitan Manila, Philippines. Infect. Genet. Evol. 2021, 87, 104676. [Google Scholar] [CrossRef]
- Chaiphongpachara, T.; Laojun, S. Variation over Time in Wing Size and Shape of the Coastal Malaria Vector Anopheles (Cellia) epiroticus Linton and Harbach (Diptera: Culicidae) in Samut Songkhram, Thailand. J. Adv. Vet. Anim. Res. 2019, 6, 208–214. [Google Scholar] [CrossRef]
- Chaiphongpachara, T.; Changbunjong, T.; Sumruayphol, S.; Laojun, S.; Suwandittakul, N.; Kuntawong, K. Geometric Morphometrics versus DNA Barcoding for the Identification of Malaria Vectors Anopheles dirus and An. baimaii in the Thai-Cambodia Border. Sci. Rep. 2022, 12, 13236. [Google Scholar] [CrossRef] [PubMed]
- Chaiphongpachara, T.; Changbunjong, T.; Laojun, S. Geometric Morphometric and Molecular Techniques for Discriminating among Three Cryptic Species of the Anopheles barbirostris Complex (Diptera: Culicidae) in Thailand. Heliyon 2022, 8, e11261. [Google Scholar] [CrossRef] [PubMed]
- Altunsoy, F.; Ercan, I.; Ocakoglu, G. Analysis of Morphometric Characteristics of Different Populations of Tabanus Bromius Linne 1758 (Diptera: Tabanidae). Pak. J. Zool. 2017, 49, 1013–1018. [Google Scholar] [CrossRef]
- Prudhomme, J.; Cassan, C.; Hide, M.; Toty, C.; Rahola, N.; Vergnes, B.; Dujardin, J.P.; Alten, B.; Sereno, D.; Bañuls, A.L. Ecology and Morphological Variations in Wings of Phlebotomus Ariasi (Diptera: Psychodidae) in the Region of Roquedur (Gard, France): A Geometric Morphometrics Approach. Parasit. Vectors 2016, 9, 578. [Google Scholar] [CrossRef] [Green Version]
- Limsopatham, K.; Tanajitaree, C.; Sanit, S.; Sukontason, K.; Somboon, P.; Amendt, J.; Feddern, N.; Sukontason, K.L. Wing Morphometrics as a Tool for the Identification of Forensic Important Lucilia spp. (Diptera: Calliphoridae). Acta Trop. 2022, 226, 106242. [Google Scholar] [CrossRef]
- García-Sánchez, A.M.; Zurita, A.; Cutillas, C. Morphometrics as a Complementary Tool in the Differentiation of Two Cosmopolitan Flea Species: Ctenocephalides felis and Ctenocephalides canis. Insects 2022, 13, 707. [Google Scholar] [CrossRef]
- Chaiphongpachara, T.; Tubsamut, P. Geometric Morphometry of Pupae to Identify Four Medically Important Flies (Order: Diptera) in Thailand. Biodiversitas 2019, 20, 1504–1509. [Google Scholar] [CrossRef] [Green Version]
- Lorenz, C.; Almeida, F.; Almeida-Lopes, F.; Louise, C.; Pereira, S.N.; Petersen, V.; Vidal, P.O.; Virginio, F.; Suesdek, L. Geometric Morphometrics in Mosquitoes: What Has Been Measured? Infect. Genet. Evol. 2017, 54, 205–215. [Google Scholar] [CrossRef]
- Ruangsittichai, J.; Apiwathnasorn, C.; Dujardin, J.P. Interspecific and Sexual Shape Variation in the Filariasis Vectors Mansonia dives and Ma. bonneae. Infect. Genet. Evol. 2011, 11, 2089–2094. [Google Scholar] [CrossRef]
- Buttachon, S.; Arikit, S.; Nuchchanart, W.; Puangmalee, T. Geometric Morphometric Analysis and Molecular Identification of Coconut Mite, Aceria guerreronis Keifer (Acari: Eriophyidae) Collected from Thailand. Insects 2022, 13, 1022. [Google Scholar] [CrossRef]
- Zubrii, N.A.; Filippov, B.Y.; Kondakov, A.V.; Khruleva, O.A.; Rybalov, L.B.; Vikhreva, D.V. DNA Barcoding versus Morphological Variability of Pterostichus brevicornis Brevicornis (Kirby, 1837) (Coleoptera, Carabidae) in the Arctic and Subarctic. Insects 2022, 13, 204. [Google Scholar] [CrossRef] [PubMed]
- Rattanarithikul, R.; Harrison, B.A.; Panthusiri, P.; Coleman, R.E. Illustrated Keys to the Mosquitoes of Thailand. I. Background; Geographic Distribution; Lists of Genera, Subgenera, and Species; and a Key to the Genera. Southeast Asian J. Trop. Med. Public Health 2005, 36, 1. [Google Scholar] [PubMed]
- Demari-Silva, B.; Suesdek, L.; Sallum, M.A.M.; Marrelli, M.T. Wing Geometry of Culex Coronator (Diptera: Culicidae) from South and Southeast Brazil. Parasit. Vectors 2014, 7, 174. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goodall, C.R. Procrustes Methods in the Statistical Analysis of Shape. J. R. Stat. Soc. Ser. B 1991, 53, 285–321. [Google Scholar] [CrossRef]
- Rohlf, F.J. Shape Statistics: Procrustes Superimpositions and Tangent Spaces. J. Classif. 1999, 16, 197–223. [Google Scholar] [CrossRef]
- Kumar, N.P.; Rajavel, A.R.; Natarajan, R.; Jambulingam, P. DNA Barcodes Can Distinguish Species of Indian Mosquitoes (Diptera: Culicidae). J. Med. Entomol. 2007, 44, 1–7. [Google Scholar] [CrossRef]
- Dujardin, S.; Dujardin, J.P. Geometric Morphometrics in the Cloud. Infect. Genet. Evol. 2019, 70, 189–196. [Google Scholar] [CrossRef]
- Hall, T.A. BIOEDIT: A User-Friendly Biological Sequence Alignment Editor and Analysis Program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; Mcgettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X Version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.; Chiang, L.; Hapuarachchi, H.C.; Tan, C.; Pang, S.; Lee, R.; Lee, K.; Ng, L. DNA Barcoding: Complementing Morphological Identification of Mosquito Species in Singapore. Parasit. Vectors 2014, 7, 569. [Google Scholar] [CrossRef] [PubMed]
- Chonephetsarath, S.; Raksakoon, C.; Sumruayphol, S.; Dujardin, J.P.; Potiwat, R. The Unequal Taxonomic Signal of Mosquito Wing Cells. Insects 2021, 12, 376. [Google Scholar] [CrossRef] [PubMed]
- Garzón, M.J.; Grech, M.; Lizuain, A.; Schweigmann, N. Geometric Morphometrics for the Differentiation of Females of the Pipiens Assemblage in Argentina. J. Vector Ecol. 2020, 45, 150–154. [Google Scholar] [CrossRef] [PubMed]
- Sauer, F.G.; Jaworski, L.; Erdbeer, L.; Heitmann, A.; Schmidt-Chanasit, J.; Kiel, E.; Lühken, R. Geometric Morphometric Wing Analysis Represents a Robust Tool to Identify Female Mosquitoes (Diptera: Culicidae) in Germany. Sci. Rep. 2020, 10, 17613. [Google Scholar] [CrossRef] [PubMed]
- Sontigun, N.; Samerjai, C.; Sukontason, K.; Wannasan, A.; Amendt, J.; Tomberlin, J.K.; Sukontason, K.L. Wing Morphometrics as a Tool in Species Identification of Forensically Important Blow Flies of Thailand. Acta Trop. 2019, 190, 312–319. [Google Scholar] [CrossRef] [PubMed]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Province | Total Number of Lutzia Mosquitoes Collected | |||
|---|---|---|---|---|
| Lt. chiangmaiensis | Lt. fuscana | Lt. halifaxii | Lt. vorax | |
| Eastern Thailand | ||||
| Trat | 8 | 13 | 32 | 6 |
| Chachoengsao | 5 | 30 | – | 4 |
| Northeastern Thailand | ||||
| Ubon Ratchathani | 5 | – | – | 4 |
| Southern Thailand | ||||
| Ranong | – | – | 15 | – |
| Western Thailand | ||||
| Kanchanaburi | 15 | – | – | 5 |
| Ratchaburi | 12 | 15 | – | 10 |
| Total | 45 | 58 | 47 | 29 |
| Lutzia Species | Pairwise Mahalanobis Distance | |||
|---|---|---|---|---|
| Lt. chiangmaiensis | Lt. fuscana | Lt. halifaxii | Lt. vorax | |
| Lt. chiangmaiensis | 0.00 | |||
| Lt. fuscana | 4.55 * | 0.00 | ||
| Lt. halifaxii | 5.34 * | 4.05 * | 0.00 | |
| Lt. vorax | 8.45 * | 10.25 * | 9.99 * | 0.00 |
| Lutzia Species | Classified as | Total (Individuals) | Correct Identifications (%) | |||
|---|---|---|---|---|---|---|
| Lt. chiangmaiensis (Individuals) | Lt. fuscana (Individuals) | Lt. halifaxii (Individuals) | Lt. vorax (Individuals) | |||
| Lt. chiangmaiensis | 37 | 1 | 2 | 0 | 40 | 92.50% |
| Lt. fuscana | 0 | 47 | 3 | 0 | 50 | 94% |
| Lt. halifaxii | 1 | 2 | 39 | 0 | 42 | 92.86% |
| Lt. vorax | 0 | 0 | 0 | 25 | 25 | 100% |
| Lutzia Species | Average Percentage Genetic Divergences (Min–Max) | |||
|---|---|---|---|---|
| Lt. chiangmaiensis | Lt. fuscana | Lt. halifaxii | Lt. vorax | |
| Lt. chiangmaiensis | 0.19% (0.00–0.71) | |||
| Lt. fuscana | 1.86% (1.43–2.45) | 0.60% (0.00–1.14) | ||
| Lt. halifaxii | 1.70% (1.57–2.16) | 0.48% (0.00–0.85) | 0.35% (0.00–0.57) | |
| Lt. vorax | 5.11% (4.81–5.44) | 4.96% (4.22–5.60) | 4.74% (4.38–5.29) | 0.76% (0.00–1.43) |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Laojun, S.; Changbunjong, T.; Chaiphongpachara, T. Evaluation of Modern Techniques for Species Identification of Lutzia Mosquitoes (Diptera: Culicidae) in Thailand: Geometric Morphometrics and DNA Barcoding. Insects 2023, 14, 78. https://doi.org/10.3390/insects14010078
Laojun S, Changbunjong T, Chaiphongpachara T. Evaluation of Modern Techniques for Species Identification of Lutzia Mosquitoes (Diptera: Culicidae) in Thailand: Geometric Morphometrics and DNA Barcoding. Insects. 2023; 14(1):78. https://doi.org/10.3390/insects14010078
Chicago/Turabian StyleLaojun, Sedthapong, Tanasak Changbunjong, and Tanawat Chaiphongpachara. 2023. "Evaluation of Modern Techniques for Species Identification of Lutzia Mosquitoes (Diptera: Culicidae) in Thailand: Geometric Morphometrics and DNA Barcoding" Insects 14, no. 1: 78. https://doi.org/10.3390/insects14010078