Germ Cell Isolation and Cryopreservation from Reproductive Organs of Brown Mealworm


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Mealworm Rearing and Diet
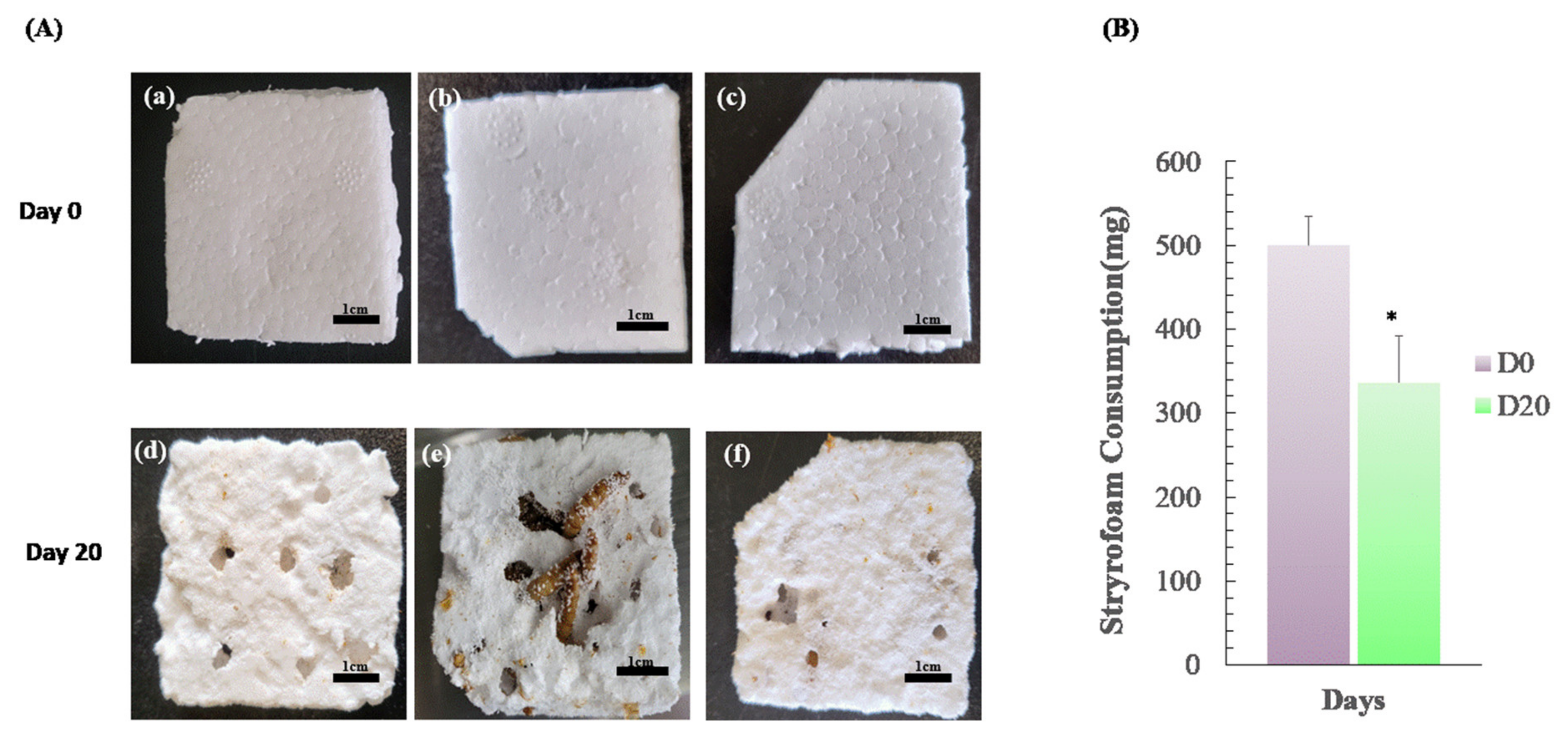
2.2. Styrofoam-Feeding Tests
2.3. Isolation of Testes, Ovaries and Germ Cells from Male and Female Adult Mealworms
2.4. Histology of Testes and Ovaries from Male or Female Adult Mealworms
2.5. Comparison of Ovary Viability of the Female Adult Mealworm between Slow-Freezing and Vitrificaiton
2.6. Ovarioles Isolation from Ovaries of Adult Female Mealworm Using Enzymatic and Mechanical Methods
2.7. Vitrification and Viability of Alginate Encapsulated Ovarioles
2.8. Statistical Analysis
3. Results and Discussion
3.1. Changes in Weight of Mealworms after Feeding on Styrofoam
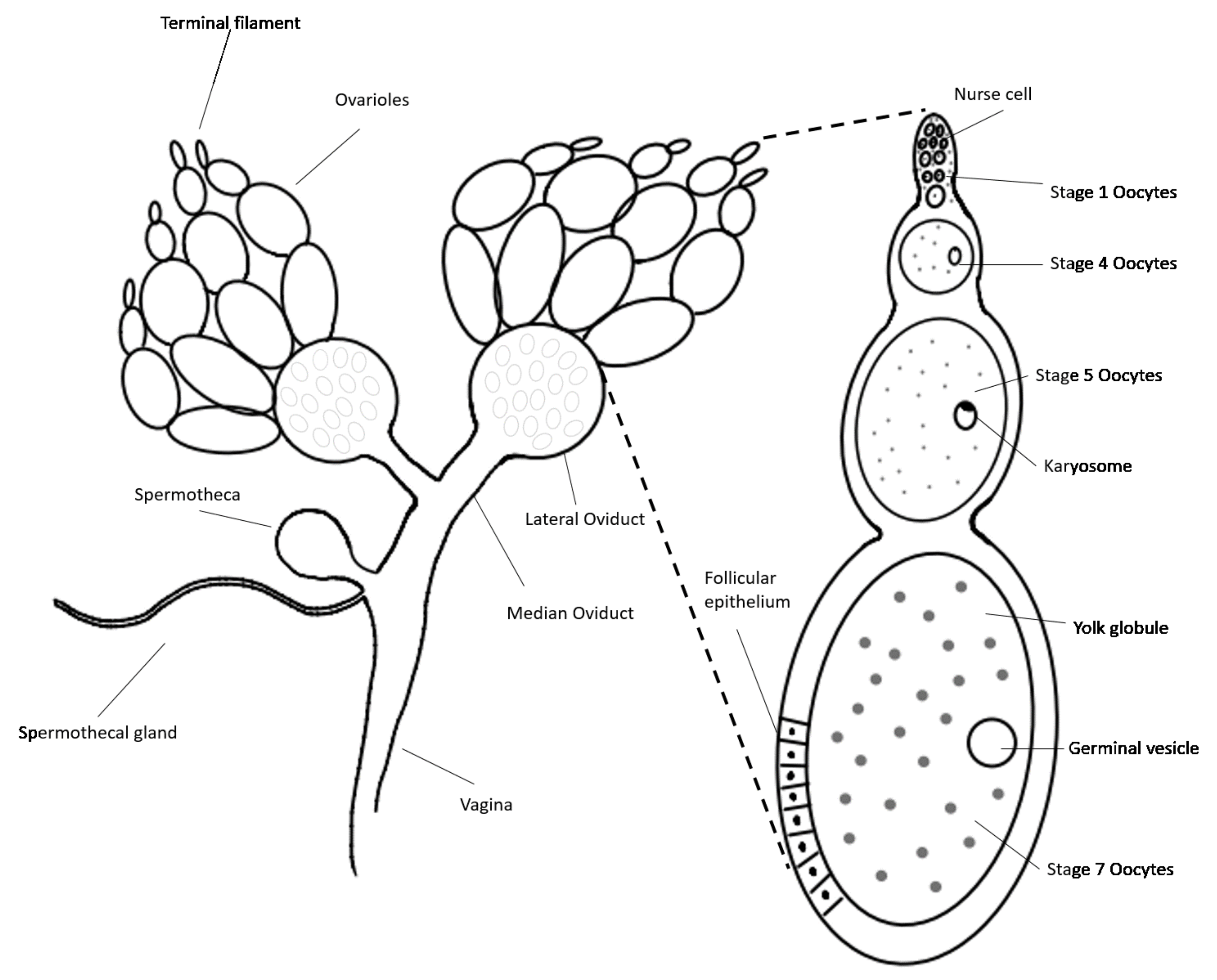
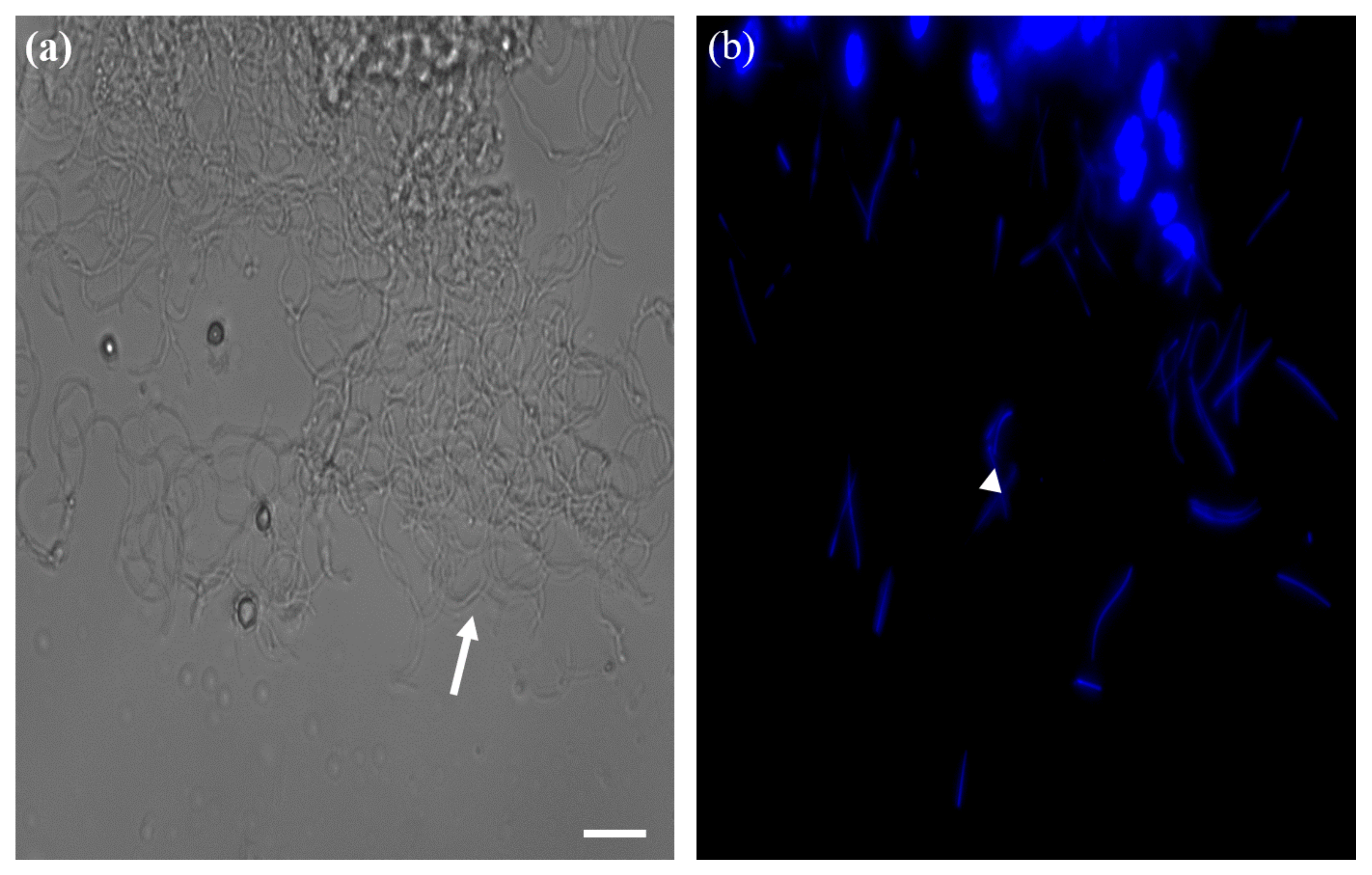
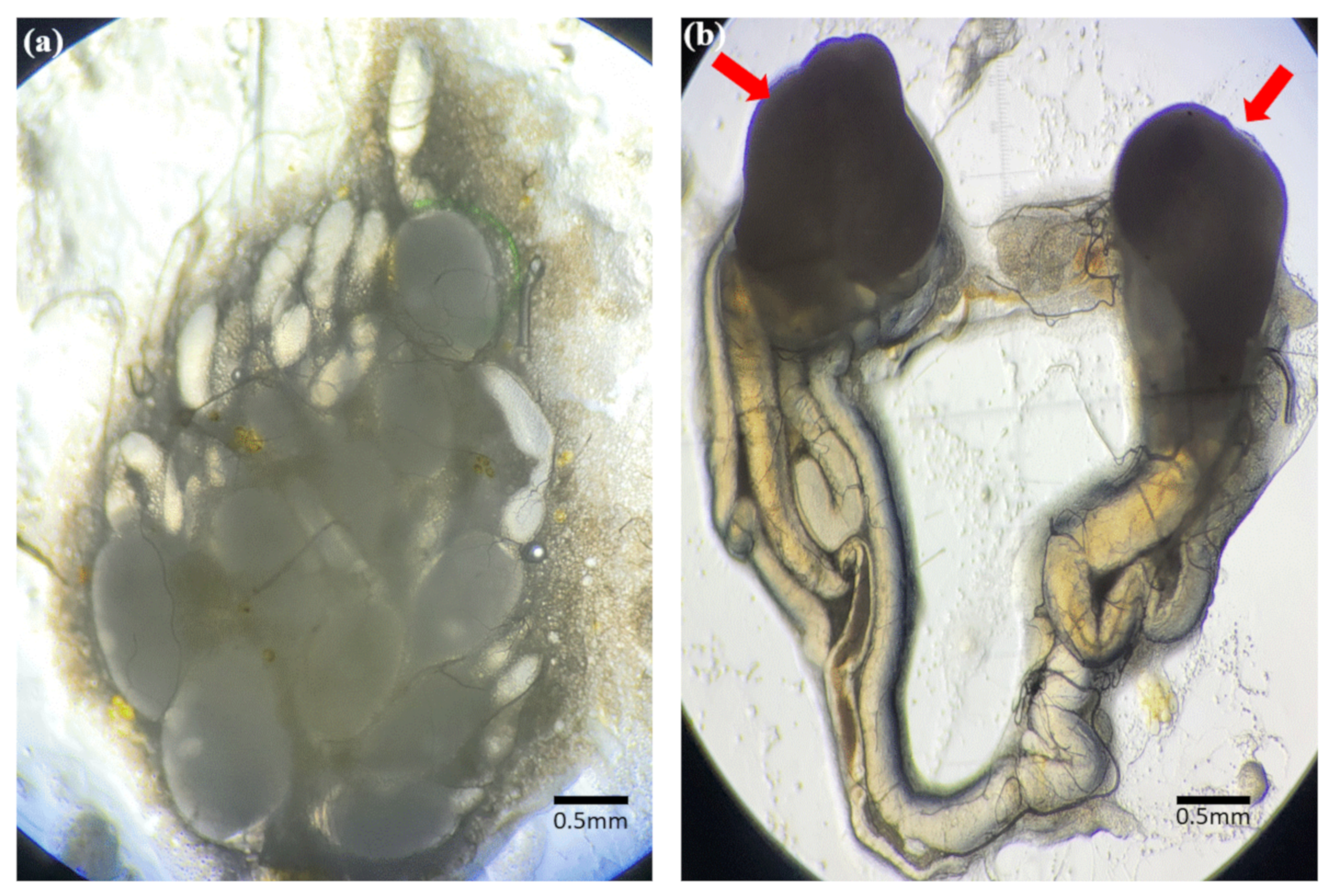
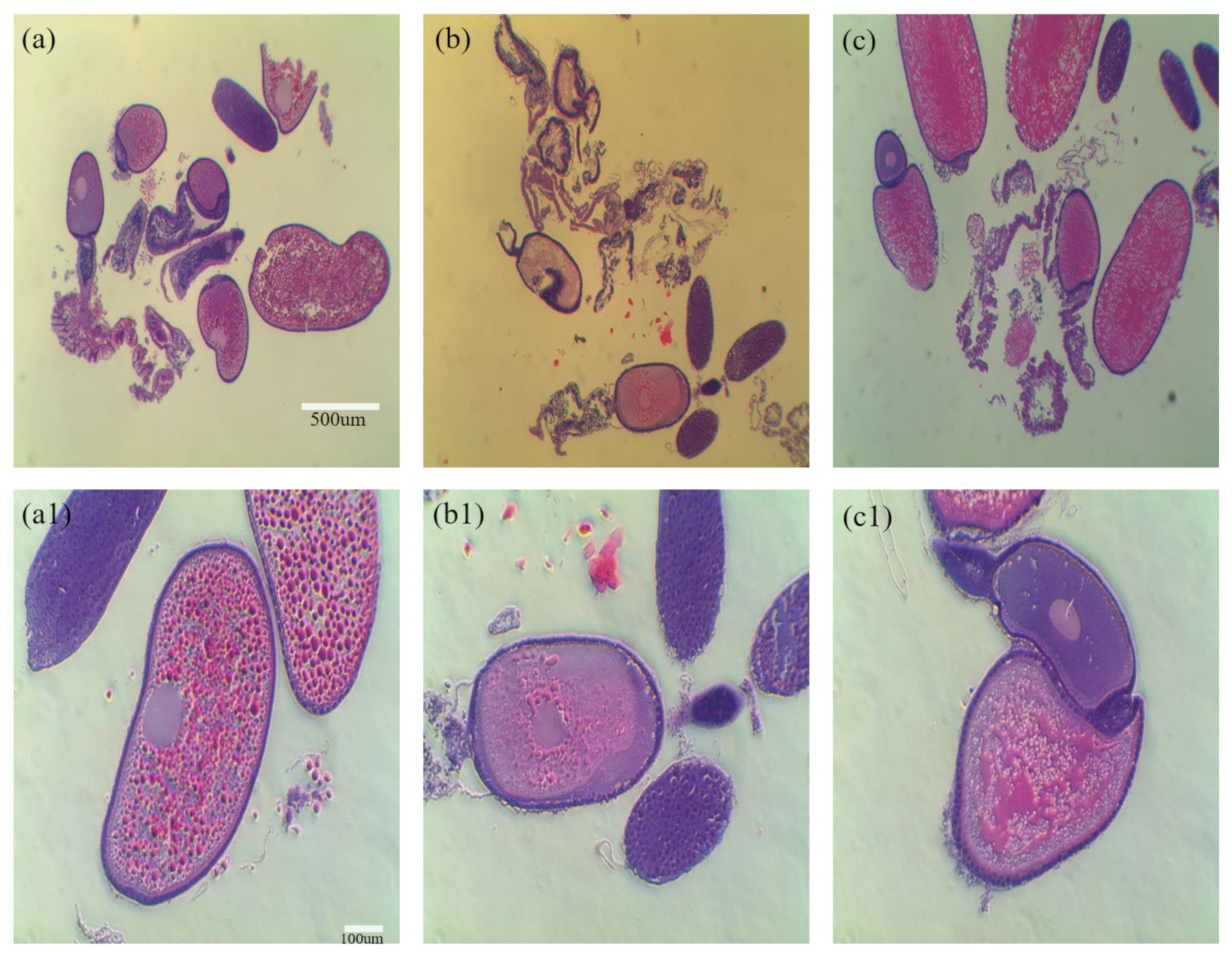
3.2. Identification of Reproductive Organs and Germ Cells of Male and Female Adult Brown Mealworms (aBMs)
3.3. Comparison between Slow-Freezing and Vitrification for Adult Brown Mealworms (aBMs)
3.4. Comparison of Enzymatic and Mechanical Methods for Ovarioles Isolation from the Ovaries of Female Adult Brown Mealworms (aBMs)
3.5. Effects of Vitrification on the Viability of Alginate-Encapsulated Ovarioles
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Kim, S.Y.; Park, B.J.; Lee, Y.B.; Yoon, H.J.; Lee, K.Y.; Kim, N.J. Growth characteristics of mealworm Tenebrio molitor. J. Seric. Entomol. Sci. 2015, 53, 1–5. [Google Scholar]
- Kim, H.S.; Shon, Y.J.; Park, S.J.; Kim, W.J.; Kang, H.J.; Yun, E.Y.; Kim, M.H. Change in Dietary Intake and Nutritional Status Using Mealworms as Hospital Meal in Postoperative Patients. J. Korean Diet. Assoc. 2016, 22, 292–309. [Google Scholar] [CrossRef] [Green Version]
- Yu, Y.; Jun, Y.; Wu, W.M.; Zhao, J.; Song, Y.; Gao, L.; Yang, R.; Lei, J. Biodegradation and Mineralization of Polystryrene by Plastic-Eating Mealworms: Part 2. Role of Gut Microorganisms. Environ. Sci. Technol. 2015, 49, 12087–12093. [Google Scholar]
- Yu, Y. Biodegradation and mineralization of polystyrene by plastic-eating mealworms part 1. Environ. Sci. Technol. 2015, 49, 12080–12086. [Google Scholar]
- Yu, Y.; Lin, H.; Xiaoxi, L.; Wang, J.; Jin, G. Nitrogen Fixation and Diazotrophic Community in Plastic-Eating Mealworms Tenebrio molitor L. Microb. Ecol. 2022, 1–13. [Google Scholar]
- Noelia, P.P.; Ramón, P.; Ana, R.; Ana, V.; Eugenio, L.C.; Aurora, G.D.; Humberto, Q.; Armando, C. Long-term exhaustion of the inbreeding load in Drosophila melanogaster. Heredity 2021, 127, 373–383. [Google Scholar]
- Harrison, K.A.; Magrath, M.J.L.; Yen, J.D.L.; Pavlova, A.; Murray, N.; Quin, B.; Menkhorst, P.; Miller, K.A.; Cartwright, K.; Sunnucks, P. Lifetime Fitness Costs of Inbreeding and Being Inbred in a Critically Endangered Bird. Curr. Biol. 2019, 29, 2711–2717. [Google Scholar]
- Aneta, K.U.; Rybak, J.; Wróbel, M.; Leluk, K.A.; Mirończuk, A.M. Comprehensive assessment of microbiome diversity in Tenebrio molitor fed with polystyrene waste. Environ. Pollut. 2020, 262, 1142812. [Google Scholar]
- Piotr, B.; Kinga, P.; Anna, P.; Andrzej, P.; Barbara, G.; Andrzej, B. Biodegradation of Different Types of Plastics by Tenebrio molitor Insect. Polymers 2021, 13, 3508. [Google Scholar]
- Borges, E.N.; Silva, R.C.; Futino, D.O.; Rocha-Junior, C.M.C.; Amorim, C.A.; Báo, S.N.; Lucci, C.M. Cryopreservation of swine ovarian tissue: Effect of different cryoprotectants on the structural preservation of preantral follicle oocytes. Cryobiology 2009, 59, 195–200. [Google Scholar] [CrossRef] [Green Version]
- Leonel EC, R.; Corral, A.; Risco, R.; Camboni, A.; Taboga, S.R.; Kilbride, P. Stepped vitrification technique for human ovarian tissue cryopreservation. Sci. Rep. 2019, 9, 20008. [Google Scholar] [CrossRef] [Green Version]
- Zahra, D.; Mohsen, N.; Shahla, Z. Cryopreservation of Bufotes viridis embryos by vitrification. Cryobiology 2017, 75, 60–67. [Google Scholar]
- Upton, R.; Clulow, S.; Calatayud, N.E.; Colyvas, K.; Seeto, R.G.; Wong, L.A. Generation of reproductively mature offspring from the endangered green and golden bell frog Litoria aurea using cryopreserved spermatozoa. Reprod. Fertil. Dev. 2021, 33, 562–572. [Google Scholar] [CrossRef]
- Campbell, L.; Cafe, S.L.; Upton, R.; Doody, J.S.; Nixon, B.; Clulow, J.; Clulow, S. A model protocol for the cryopreservation and recovery of motile lizard sperm using the phosphodiesterase inhibitor caffeine. Conserv. Physiol. 2020, 22, 8. [Google Scholar] [CrossRef]
- Campbell, L.; John, C.; Doody, J.S.; Clulow, S. Optimal cooling rates for sperm cryopreservation in a threatened lizard conform to two-factor hypothesis of cryo-injury. Cryobiology 2021, 103, 101–106. [Google Scholar] [CrossRef]
- He, X. Methodological Advances in the Culture, Manipulation and Utilization of Embryonic Stem Cells for Basic and Practical Applications; Atwood, C., Ed.; Tech Publisher: Vienna, Austria, 2011; p. 113. [Google Scholar]
- Karlsson, O.M.; Cravalho, E.G.; Toner, M. A model of diffusion-limited ice growth inside biological cells during freezing. J. Appl. Phys. 1994, 75, 4442. [Google Scholar] [CrossRef]
- Zhao, G.; Takamatsu, H.; He, X. The effect of solution nonideality on modeling transmembrane water transport and diffusion-limited intracellular ice formation during cryopreservation. J. Appl. Phys. 2014, 115, 144701. [Google Scholar] [CrossRef] [Green Version]
- Zhang, W.; Yang, G.; Zhang, A.; Xu, L.X.; He, X. Preferential vitrification of water in small alginate microcapsules significantly augments cell cryopreservation by vitrification. Biomed. Microdevices 2010, 12, 89–96. [Google Scholar] [CrossRef]
- Choi, J.K.; Agarwal, P.; He, X. In Vitro Culture of Early Secondary Preantral Follicles in Hanging Drop of Ovarian Cell-Conditioned Medium to Obtain MII Oocytes from Outbred Deer Mice. Tissue Eng. Part A 2013, 19, 2626–2637. [Google Scholar] [CrossRef]
- Wafa, B.; Jessica, K.; Jérôme, B.; Nicolas, B. Encapsulation of Cells in a Collagen Matrix Surrounded by an Alginate Hydrogel Shell for 3D Cell Culture. ACS Biomater. Sci. Eng. 2022, 8, 2700–2708. [Google Scholar]
- Aaron, G.M.; Gregory, S.K. Alginate modification improves long-term survival and function of transplanted encapsulated islets. Tissue Eng. Part A 2009, 15, 1301–1309. [Google Scholar]
- Huang, H.; Choi, J.K.; Rao, W.; Zhao, S.; Agarwal, P.; Zhao, G.; He, X. Alginate Hydrogel Microencapsulation Inhibits Devitrification and Enables Large-Volume Low-CPA Cell Vitrification. Adv. Funct. Mater. 2015, 25, 6839–6850. [Google Scholar] [CrossRef]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Group | No. of Total Ovarioles (%) | No. of Ruptured Ovarioles (%) | No. of Retrieved Ovarioles (%) |
|---|---|---|---|
| Vitrification without alginate capsule | 13 (100) | 7 (53.8) | 6 (46.2) |
| Vitrification with alginate capsule | 24 (100) | 11 (45.8) | 13 (54.2) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Byeun, D.G.; Moon, B.-S.; Lee, S.; Choi, J.K. Germ Cell Isolation and Cryopreservation from Reproductive Organs of Brown Mealworm. Insects 2022, 13, 1108. https://doi.org/10.3390/insects13121108
Byeun DG, Moon B-S, Lee S, Choi JK. Germ Cell Isolation and Cryopreservation from Reproductive Organs of Brown Mealworm. Insects. 2022; 13(12):1108. https://doi.org/10.3390/insects13121108
Chicago/Turabian StyleByeun, Do Gyeung, Byoung-San Moon, Seungki Lee, and Jung Kyu Choi. 2022. "Germ Cell Isolation and Cryopreservation from Reproductive Organs of Brown Mealworm" Insects 13, no. 12: 1108. https://doi.org/10.3390/insects13121108