
Warm Blood Meal Increases Digestion Rate and Milk Protein Production to Maximize Reproductive Output for the Tsetse Fly, Glossina morsitans


 ,
,  ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Flies and Description of Studies
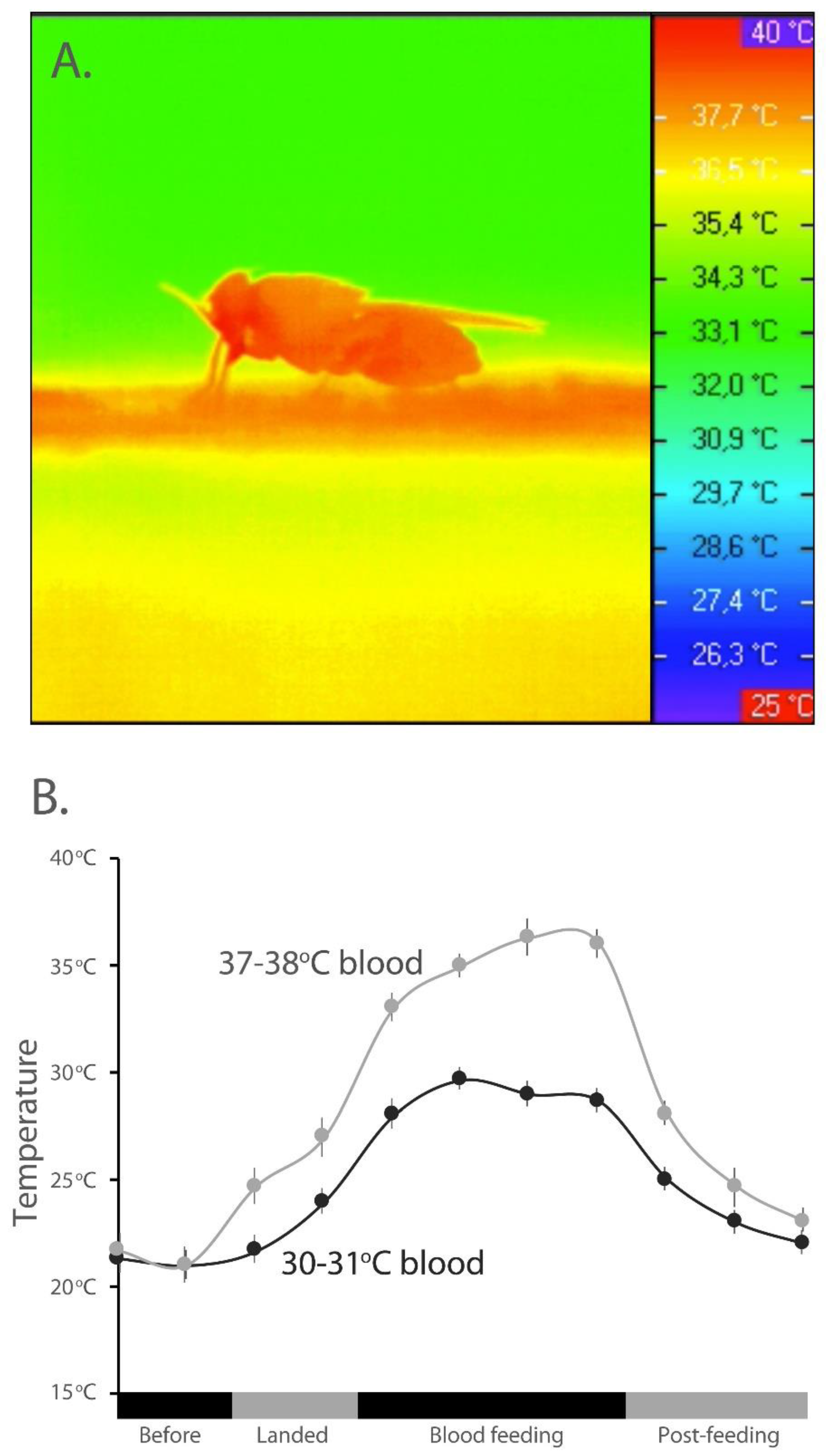
2.2. Thermal Imaging and Contact Thermal Changes during Blood Meal
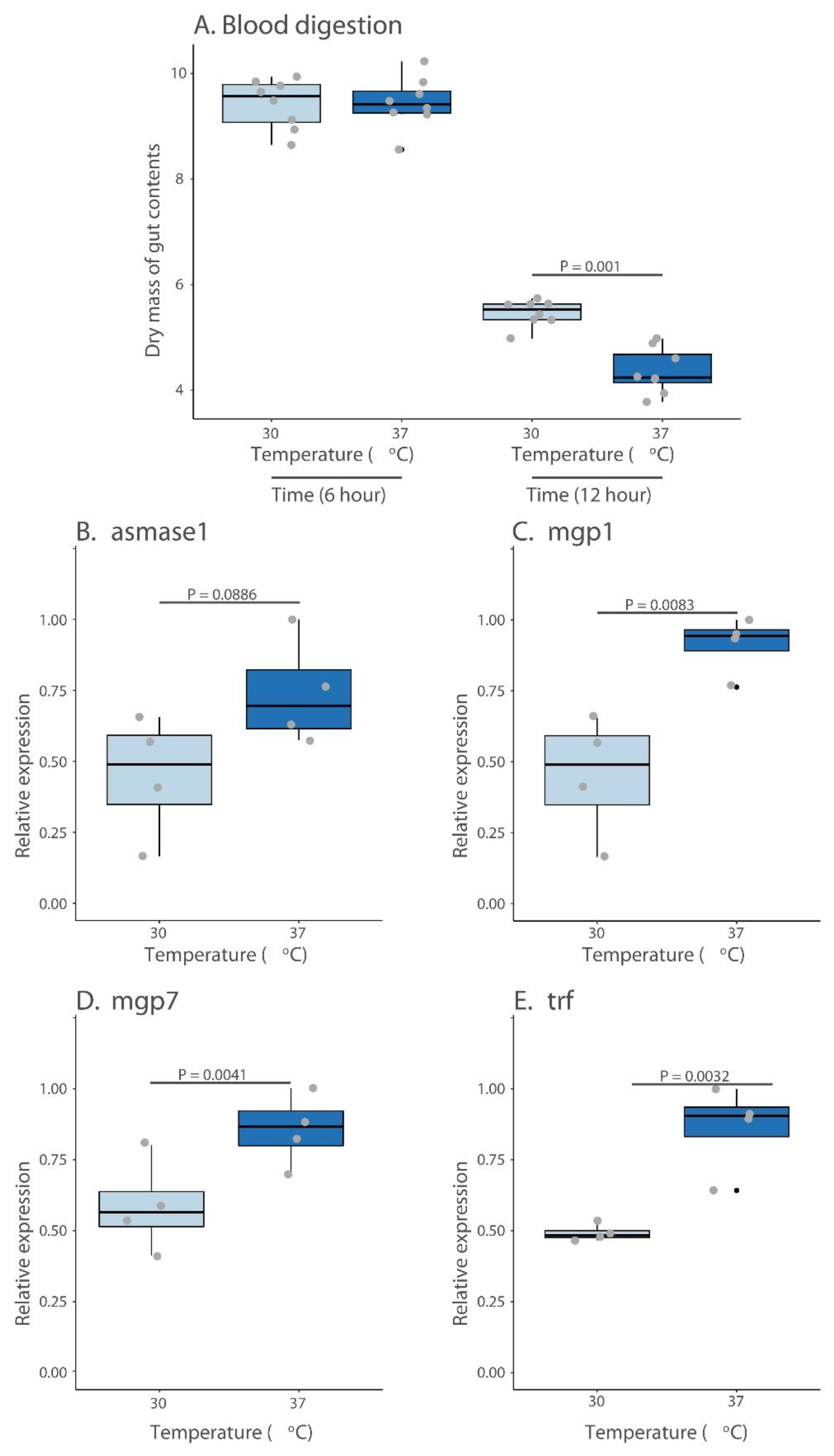
2.3. Blood Digestion
2.4. RNA
2.5. Quantitative PCR
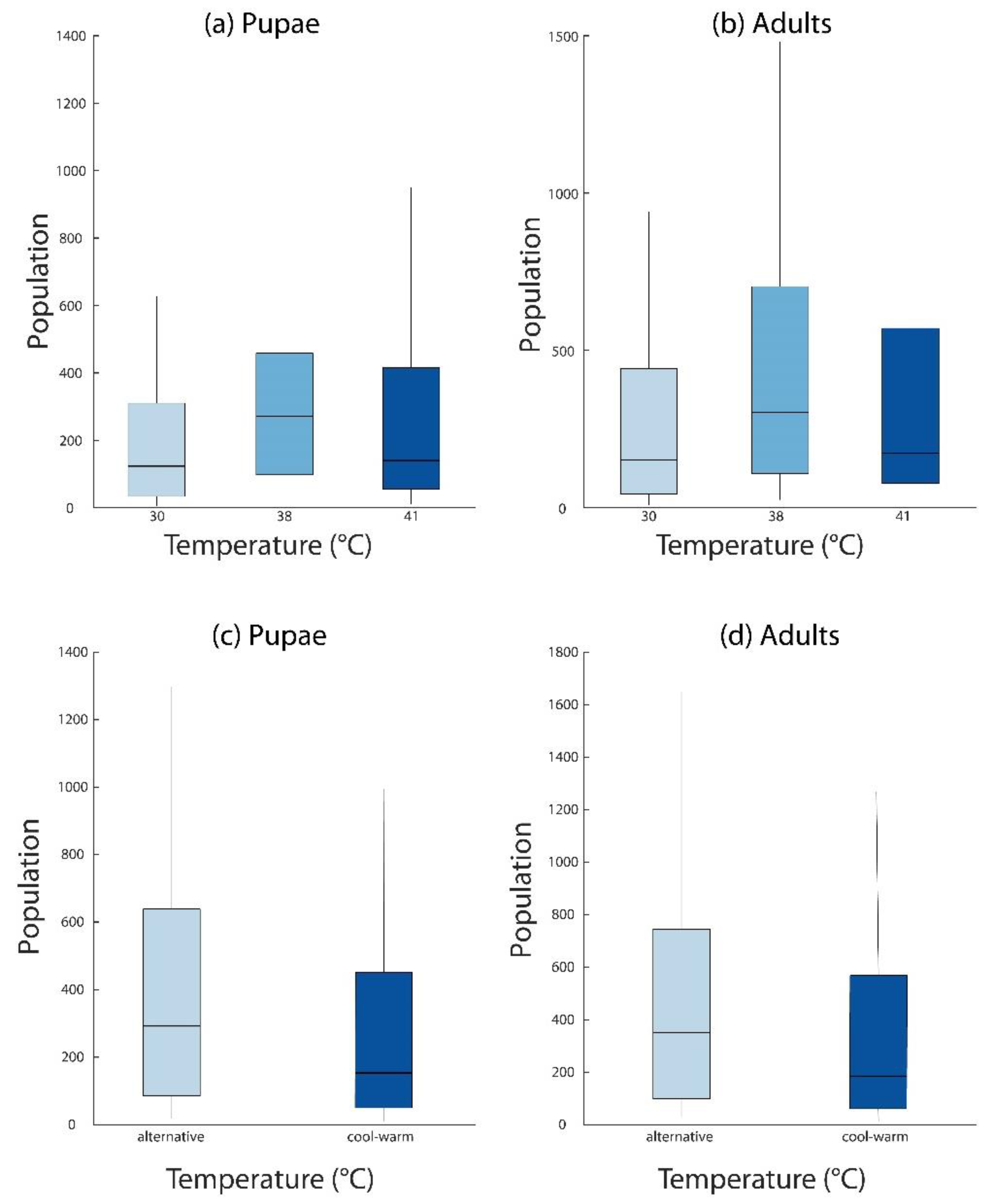
2.6. Population Modeling
3. Results
3.1. Temperature Changes in Tsetse Flies Associated with a Blood Meal
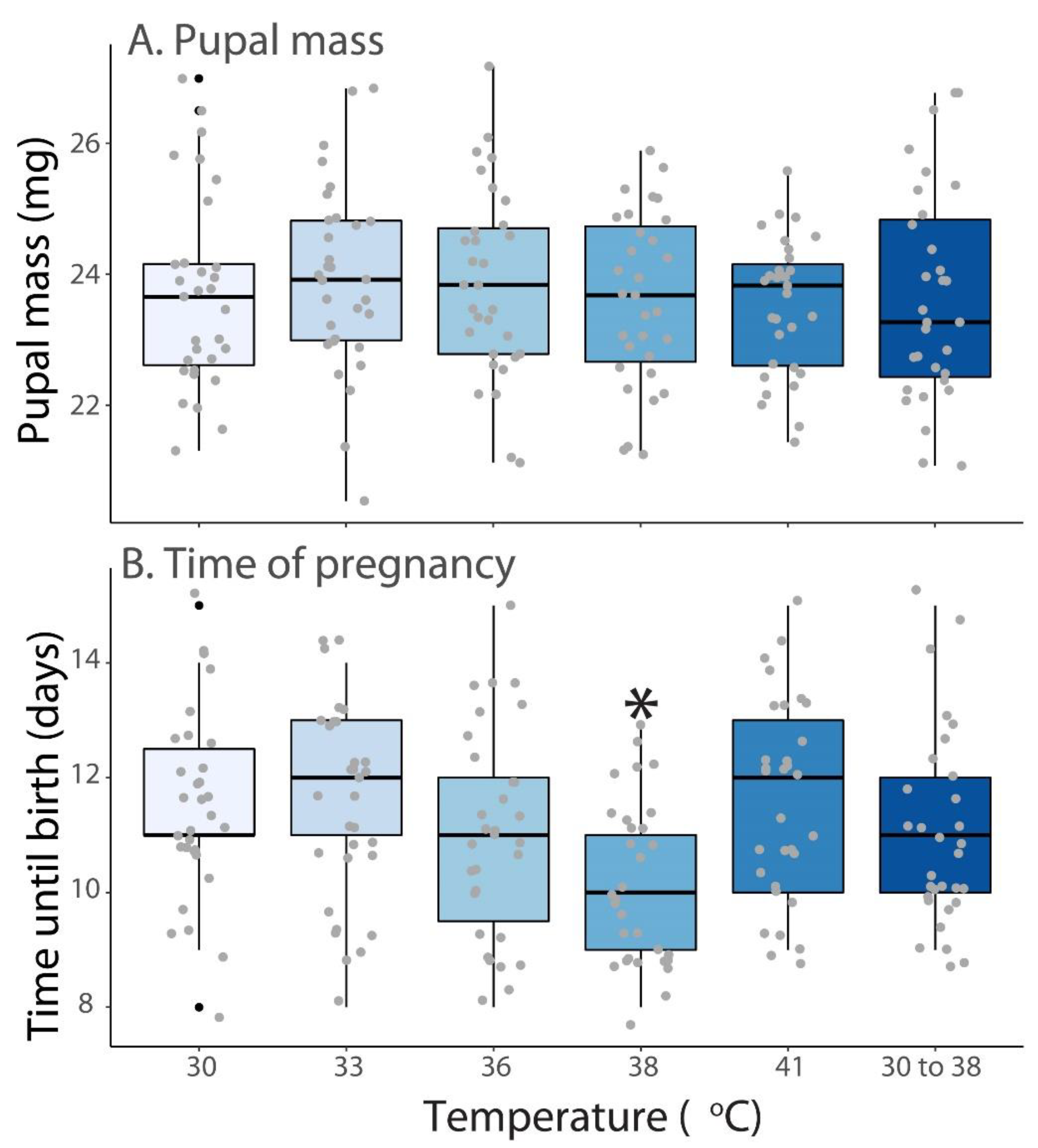
3.2. Pupae Size and Duration of Pregnancy
3.3. Blood Processing and Milk Protein Synthesis
3.4. Population Modeling
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Benoit, J.B.; Attardo, G.M.; Baumann, A.A.; Michalkova, V.; Aksoy, S. Adenotrophic Viviparity in Tsetse Flies: Potential for Population Control and as an Insect Model for Lactation. Annu. Rev. Entomol. 2015, 60, 351–371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meier, R.; Kotrba, M.; Ferrar, P. Ovoviviparity and Viviparity in the Diptera. Biol. Rev. Camb. Philos. Soc. 1999, 74, 199–258. [Google Scholar] [CrossRef]
- Tobe, S.S. Reproductive Physiology of Glossina. Annu. Rev. Entomol. 1978, 23, 283–307. [Google Scholar] [CrossRef] [PubMed]
- Lord, J.S.; Leyland, R.; Haines, L.R.; Barreaux, A.M.G.; Bonsall, M.B.; Torr, S.J.; English, S. Effects of Maternal Age and Stress on Offspring Quality in a Viviparous Fly. Ecol. Lett. 2021, 24, 2113–2122. [Google Scholar] [CrossRef] [PubMed]
- Michalkova, V.; Benoit, J.B.; Attardo, G.M.; Medlock, J.; Aksoy, S. Amelioration of Reproduction-Associated Oxidative Stress in a Viviparous Insect Is Critical to Prevent Reproductive Senescence. PLoS ONE 2014, 9, e87554. [Google Scholar] [CrossRef]
- Cmelik, S.H.W.; Bursell, E.; Slack, E. Composition of the Gut Contents of Third-Instar Tsetse Larvae (Glossina morsitans Westwood). Comp. Biochem. Physiol. 1969, 29, 447–453. [Google Scholar] [CrossRef]
- Benoit, J.B.; Attardo, G.M.; Michalkova, V.; Krause, T.B.; Bohova, J.; Zhang, Q.; Baumann, A.A.; Mireji, P.O.; Takáč, P.; Denlinger, D.L.; et al. A Novel Highly Divergent Protein Family Identified from a Viviparous Insect by RNA-Seq Analysis: A Potential Target for Tsetse Fly-Specific Abortifacients. PLoS Genet. 2014, 10, e1003874. [Google Scholar] [CrossRef] [Green Version]
- Guz, N.; Attardo, G.M.; Wu, Y.; Aksoy, S. Molecular Aspects of Transferrin Expression in the Tsetse Fly (Glossina Morsitans Morsitans). J. Insect Physiol. 2007, 53, 715–723. [Google Scholar] [CrossRef] [Green Version]
- Attardo, G.M.; Guz, N.; Strickler-Dinglasan, P.; Aksoy, S. Molecular Aspects of Viviparous Reproductive Biology of the Tsetse Fly (Glossina Morsitans Morsitans): Regulation of Yolk and Milk Gland Protein Synthesis. J. Insect Physiol. 2006, 52, 1128–1136. [Google Scholar] [CrossRef] [Green Version]
- Attardo, G.M.; Abd-Alla, A.M.M.; Acosta-Serrano, A.; Allen, J.E.; Bateta, R.; Benoit, J.B.; Bourtzis, K.; Caers, J.; Caljon, G.; Christensen, M.B.; et al. Comparative Genomic Analysis of Six Glossina Genomes, Vectors of African Trypanosomes. Genome Biol. 2019, 20, 187. [Google Scholar] [CrossRef]
- International Glossina Genome Initiative Genome Sequence of the Tsetse Fly (Glossina morsitans): Vector of African Trypanosomiasis. Science 2014, 344, 380–386. [CrossRef] [PubMed] [Green Version]
- Benoit, J.B.; Attardo, G.M.; Michalkova, V.; Takác, P.; Bohova, J.; Aksoy, S. Sphingomyelinase Activity in Mother’s Milk Is Essential for Juvenile Development: A Case from Lactating Tsetse Flies. Biol. Reprod. 2012, 87, 17. [Google Scholar] [CrossRef] [PubMed]
- Lahondère, C.; Lazzari, C.R. Mosquitoes Cool down during Blood Feeding to Avoid Overheating. Curr. Biol. 2012, 22, 40–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benoit, J.B.; Lopez-Martinez, G.; Patrick, K.R.; Phillips, Z.P.; Krause, T.B.; Denlinger, D.L. Drinking a Hot Blood Meal Elicits a Protective Heat Shock Response in Mosquitoes. Proc. Natl. Acad. Sci. USA 2011, 108, 8026–8029. [Google Scholar] [CrossRef] [Green Version]
- Lahondère, C.; Insausti, T.C.; Paim, R.M.; Luan, X.; Belev, G.; Pereira, M.H.; Ianowski, J.P.; Lazzari, C.R. Countercurrent Heat Exchange and Thermoregulation during Blood-Feeding in Kissing Bugs. Elife 2017, 6, e26107. [Google Scholar] [CrossRef]
- Benoit, J.B.; Lazzari, C.R.; Denlinger, D.L.; Lahondère, C. Thermoprotective Adaptations Are Critical for Arthropods Feeding on Warm-Blooded Hosts. Curr. Opin. Insect Sci. 2019, 34, 7–11. [Google Scholar] [CrossRef]
- Lahondère, C.; Lazzari, C.R. Thermal Effect of Blood Feeding in the Telmophagous Fly Glossina morsitans morsitans. J. Therm. Biol. 2015, 48, 45–50. [Google Scholar] [CrossRef]
- McCue, M.D.; Boardman, L.; Clusella-Trullas, S.; Kleynhans, E.; Terblanche, J.S. The Speed and Metabolic Cost of Digesting a Blood Meal Depends on Temperature in a Major Disease Vector. J. Exp. Biol. 2016, 219, 1893–1902. [Google Scholar] [CrossRef] [Green Version]
- Moloo, S.K. An Artificial Feeding Technique for Glossina. Parasitology 1971, 63, 507–512. [Google Scholar] [CrossRef]
- Stabentheiner, A. Thermographic Determination of Body Temperatures in Honey Bees and Hornets: Calibration and Applications. Thermology 1987, 2, 563–572. [Google Scholar]
- Langley, P.A.; Pimley, R.W.; Mews, A.R.; Flood, M.E.T. Effect of Diet Composition on Feeding, Digestion, and Reproduction in Glossina Morsitans. J. Insect Physiol. 1978, 24, 233–238. [Google Scholar] [CrossRef]
- Benoit, J.B.; Michalkova, V.; Didion, E.M.; Xiao, Y.; Baumann, A.A.; Attardo, G.M.; Aksoy, S. Rapid Autophagic Regression of the Milk Gland during Involution Is Critical for Maximizing Tsetse Viviparous Reproductive Output. PLoS Negl. Trop. Dis. 2018, 12, e0006204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Benoit, J.B.; Hansen, I.A.; Attardo, G.M.; Michalková, V.; Mireji, P.O.; Bargul, J.L.; Drake, L.L.; Masiga, D.K.; Aksoy, S. Aquaporins Are Critical for Provision of Water during Lactation and Intrauterine Progeny Hydration to Maintain Tsetse Fly Reproductive Success. PLoS Negl. Trop. Dis. 2014, 8, e2517. [Google Scholar] [CrossRef] [PubMed]
- Hu, C.; Rio, R.V.M.; Medlock, J.; Haines, L.R.; Nayduch, D.; Savage, A.F.; Guz, N.; Attardo, G.M.; Pearson, T.W.; Galvani, A.P.; et al. Infections with Immunogenic Trypanosomes Reduce Tsetse Reproductive Fitness: Potential Impact of Different Parasite Strains on Vector Population Structure. PLoS Negl. Trop. Dis. 2008, 2, e192. [Google Scholar] [CrossRef]
- Sterkel, M.; Oliveira, J.H.M.; Bottino-Rojas, V.; Paiva-Silva, G.O.; Oliveira, P.L. The Dose Makes the Poison: Nutritional Overload Determines the Life Traits of Blood-Feeding Arthropods. Trends Parasitol. 2017, 33, 633–644. [Google Scholar] [CrossRef] [PubMed]
- Attardo, G.M.; Benoit, J.B.; Michalkova, V.; Yang, G.; Roller, L.; Bohova, J.; Takáč, P.; Aksoy, S. Analysis of Lipolysis Underlying Lactation in the Tsetse Fly, Glossina morsitans. Insect Biochem. Mol. Biol. 2012, 42, 360–370. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Langley, P.A.; Bursell, E.; Kabayo, J.; Pimley, R.W.; Trewern, M.A.; Marshall, J. Haemolymph Lipid Transport from Fat Body to Uterine Gland in Pregnant Females of Glossina Morsitans. Insect Biochem. 1981, 11, 225–231. [Google Scholar] [CrossRef]
- Houdas, Y.; Ring, E.F.J. Human Body Temperature: Its Measurement and Regulation; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Protsiv, M.; Ley, C.; Lankester, J.; Hastie, T.; Parsonnet, J. Decreasing Human Body Temperature in the United States Since the Industrial Revolution. Elife 2020, 9, e49555. [Google Scholar] [CrossRef]




Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Benoit, J.B.; Lahondère, C.; Attardo, G.M.; Michalkova, V.; Oyen, K.; Xiao, Y.; Aksoy, S. Warm Blood Meal Increases Digestion Rate and Milk Protein Production to Maximize Reproductive Output for the Tsetse Fly, Glossina morsitans. Insects 2022, 13, 997. https://doi.org/10.3390/insects13110997
Benoit JB, Lahondère C, Attardo GM, Michalkova V, Oyen K, Xiao Y, Aksoy S. Warm Blood Meal Increases Digestion Rate and Milk Protein Production to Maximize Reproductive Output for the Tsetse Fly, Glossina morsitans. Insects. 2022; 13(11):997. https://doi.org/10.3390/insects13110997
Chicago/Turabian StyleBenoit, Joshua B., Chloé Lahondère, Geoffrey M. Attardo, Veronika Michalkova, Kennan Oyen, Yanyu Xiao, and Serap Aksoy. 2022. "Warm Blood Meal Increases Digestion Rate and Milk Protein Production to Maximize Reproductive Output for the Tsetse Fly, Glossina morsitans" Insects 13, no. 11: 997. https://doi.org/10.3390/insects13110997