


RNAi-Mediated Manipulation of Cuticle Coloration Genes in Lygus hesperus Knight (Hemiptera: Miridae)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Insects
2.2. Bioinformatics
2.3. PCR
2.4. RNAi-Mediated Knockdown
3. Results
3.1. Identification and Bioinformatic Analyses
3.2. RT-PCR Cloning and Expression Profiling
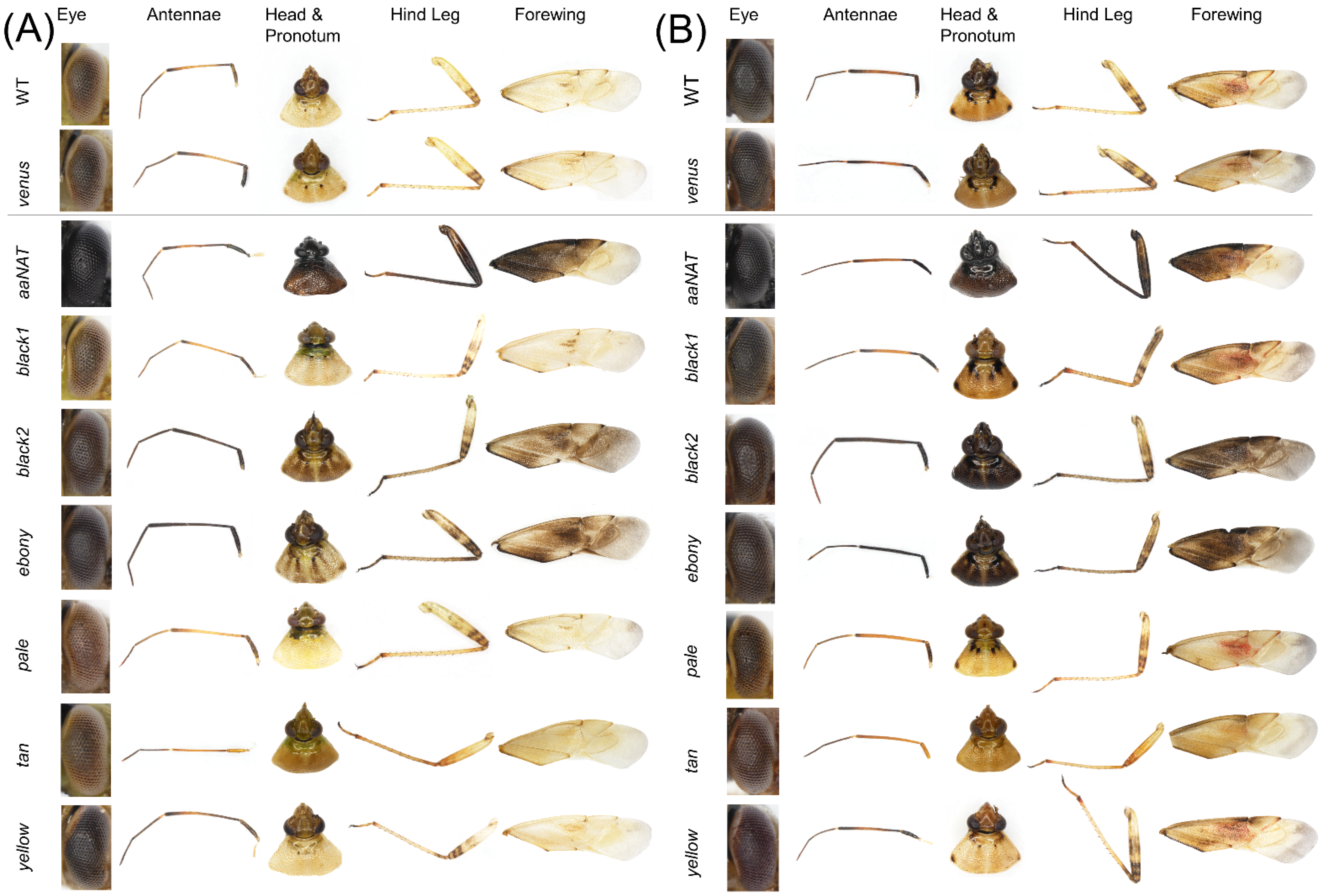
3.3. RNAi-Mediated Knockdown
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Scott, D.R. An annotated listing of host plants of Lygus hesperus Knight. Bull. Entomol. Soc. Am. 1977, 23, 19–22. [Google Scholar] [CrossRef]
- Wheeler, A.G. Biology of the Plant Bugs (Hemiptera: Miridae): Pests, Predators, Opportunists; Comstock Publishing Associates: Ithaca, NY, USA, 2001; p. 562. [Google Scholar]
- Naranjo, S.; Ellsworth, P.; Dierig, D. Impact of Lygus spp. (Hemiptera: Miridae) on damage, yield and quality of lesquerella (Physaria fendleri), a potential new oil-seed crop. J. Econ. Entomol. 2011, 104, 1575–1583. [Google Scholar] [CrossRef] [PubMed]
- Ritter, R.A.; Lenssen, A.; Blodgett, S.; Taper, M.A. Regional assemblages of Lygus (Heteroptera: Miridae) in Montana canola fields. J. Kans. Entomol. Soc. 2010, 83, 297–305. [Google Scholar] [CrossRef]
- Strand, L.L. Integrated Pest Management for Strawberries, 2nd ed.; University of California Agriculture and Natural Resources Publications: Oakland, CA, USA, 2008; p. 3351. [Google Scholar]
- Hull, J.J.; Geib, S.M.; Fabrick, J.A.; Brent, C.S. Sequencing and de novo assembly of the western tarnished plant bug (Lygus hesperus) transcriptome. PLoS ONE 2013, 8, e55105. [Google Scholar] [CrossRef] [PubMed]
- Hull, J.J.; Chaney, K.; Geib, S.M.; Fabrick, J.A.; Brent, C.S.; Walsh, D.; Lavine, L.C. Transcriptome-based identification of ABC transporters in the western tarnished plant bug Lygus hesperus. PLoS ONE 2014, 9, e113046. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tassone, E.E.; Geib, S.M.; Hall, B.; Fabrick, J.A.; Brent, C.S.; Hull, J.J. De novo construction of an expanded transcriptome assembly for the western tarnished plant bug, Lygus hesperus. GigaScience 2016, 5, 6. [Google Scholar] [CrossRef] [Green Version]
- Van Ekert, E.; Wang, M.; Miao, Y.G.; Brent, C.S.; Hull, J.J. RNA interference-mediated knockdown of the Halloween gene spookiest (CYP307B1) impedes adult eclosion in the western tarnished plant bug, Lygus hesperus. Insect Mol. Biol. 2016, 25, 550–565. [Google Scholar] [CrossRef]
- Brent, C.S.; Hull, J.J. RNA interference-mediated knockdown of eye coloration genes in the western tarnished plant bug (Lygus hesperus Knight). Arch. Insect Biochem. Physiol. 2019, 100, e21527. [Google Scholar] [CrossRef] [PubMed]
- Hull, J.J.; Yang, Y.-W.; Miyasaki, K.; Brent, C.S. TRPA1 modulates noxious odor responses in Lygus hesperus. J. Insect Physiol. 2020, 122, 104038. [Google Scholar] [CrossRef]
- Hull, J.J.; Gross, R.J.; Brent, C.S.; Christie, A.E. Filling in the gaps: A reevaluation of the Lygus hesperus peptidome using an expanded de novo assembled transcriptome and molecular cloning. Gen. Comp. Endocrinol. 2021, 303, 113708. [Google Scholar] [CrossRef]
- Hull, J.J.; Brent, C.S.; Choi, M.-Y.; Mikó, Z.; Fodor, J.; Fónagy, A. Molecular and functional characterization of pyrokinin-like peptides in the western tarnished plant bug Lygus hesperus (Hemiptera: Miridae). Insects 2021, 12, 914. [Google Scholar] [CrossRef] [PubMed]
- Heu, C.C.; Gross, R.J.; Le, K.P.; LeRoy, D.M.; Fang, B.; Hull, J.J.; Brent, C.S.; Fabrick, J.A. CRISPR-mediated knockout of cardinal and cinnabar eye pigmentation genes in the western tarnished plant bug. Sci. Rep. 2022, 12, 4917. [Google Scholar] [CrossRef] [PubMed]
- Zhan, S.; Guo, Q.; Li, M.; Li, M.; Li, J.; Miao, X.; Huang, Y. Disruption of an N-acetyltransferase gene in the silkworm reveals a novel role in pigmentation. Development 2010, 137, 4083–4090. [Google Scholar] [CrossRef] [Green Version]
- Osanai-Futahashi, M.; Ohde, T.; Hirata, J.; Uchino, K.; Futahashi, R.; Tamura, T.; Niimi, T.; Sezutsu, H. A visible dominant marker for insect transgenesis. Nat. Commun. 2012, 3, 1295. [Google Scholar] [CrossRef] [Green Version]
- Liu, J.; Lemonds, T.R.; Marden, J.H.; Popadić, A. A pathway analysis of melanin patterning in a hemimetabolous insect. Genetics 2016, 203, 403–413. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, X.L.; Han, W.K.; Ze, L.J.; Peng, Y.C.; Yang, Y.L.; Zhang, J.; Yan, Q.; Dong, S.L. Clustered regularly interspaced short palindromic repeats/CRISPR-associated protein 9 mediated knockout reveals functions of the yellow-y gene in Spodoptera litura. Front. Physiol. 2020, 11, 615391. [Google Scholar] [CrossRef]
- Massey, J.H.; Akiyama, N.; Bien, T.; Dreisewerd, K.; Wittkopp, P.J.; Yew, J.Y.; Takahashi, A. Pleiotropic effects of ebony and tan on pigmentation and cuticular hydrocarbon composition in Drosophila melanogaster. Front. Physiol. 2019, 10, 518. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Ze, L.; Jin, L.; Li, G.-Q. Yellow-b, -c, -d, and -h are required for normal body coloration of Henosepilachna vigintioctopunctata. Arch. Insect Biochem. Physiol. 2021, 109, e21856. [Google Scholar] [CrossRef]
- Ze, L.J.; Jin, L.; Li, G.Q. Silencing of Adc and Ebony causes abnormal darkening of cuticle in Henosepilachna vigintioctopunctata. Front. Physiol. 2022, 13, 829675. [Google Scholar] [CrossRef]
- Chen, J.X.; Li, W.X.; Lyu, J.; Hu, Y.T.; Huang, G.; Zhang, W.Q. CRISPR/Cas9-mediated knockout of the NlCSAD gene results in darker cuticle pigmentation and a reduction in female fecundity in Nilaparvata lugens (Hemiptera: Delphacidae). Comp. Biochem. Physiol. A 2021, 256, 110921. [Google Scholar] [CrossRef]
- Strong, F.E.; Sheldahl, J.A.; Hughs, P.R.; Hussein, E.M.K. Reproductive biology of Lygus hesperus Knight. Hilgardia 1970, 40, 105–147. [Google Scholar] [CrossRef] [Green Version]
- Mueller, S.C.; Summers, C.G.; Goodell, P.B. A Field Key to the Most Common Lygus Species Found in Agronomic Crops of the Central San Joaquin Valley of California; University of California Agriculture and Natural Resources Publications: Oakland, CA, USA, 2003; p. 8104. [Google Scholar]
- Brent, C.S. Classification of diapause status by color phenotype in Lygus hesperus. J. Insect Sci. 2012, 12, 136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wright, T.R. The genetics of biogenic amine metabolism, sclerotization, and melanization in Drosophila melanogaster. Adv. Genet. 1987, 24, 127–222. [Google Scholar] [PubMed]
- Hinaux, H.; Bachem, K.; Battistara, M.; Rossi, M.; Xin, Y.; Jaenichen, R.; Le Pul, Y.; Arnoult, L.; Kobler, J.M.; Grunwald Kadow, I.C.; et al. Revisiting the developmental and cellular role of the pigmentation gene yellow in Drosophila using a tagged allele. Dev. Biol. 2018, 438, 111–123. [Google Scholar] [CrossRef] [PubMed]
- Dai, F.; Qiao, L.; Tong, X.; Cao, C.; Chen, P.; Chen, J.; Lu, C.; Xiang, Z.-H. Mutations of an arylalkylamine-n-acetyl transferase, BM-iAANAT, are responsible for silkworm melanism mutant. J. Biol. Chem. 2010, 285, 19553–19560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, Y.; Li, J.; Zhao, T.; Li, G.; Zhu, Y. A new arylalkylamine N-acetyltransferase in silkworm (Bombyx mori) affects integument pigmentation. Appl. Biochem. Biotechnol. 2015, 175, 3447–3457. [Google Scholar] [CrossRef]
- Ahmed-Braimah, Y.H.; Sweigart, A.L. A single gene causes an interspecific difference in pigmentation in Drosophila. Genetics 2015, 200, 331–342. [Google Scholar] [CrossRef] [Green Version]
- Noh, M.Y.; Koo, B.; Kramer, K.J.; Muthukrishnan, S.; Arakane, Y. Arylalkylamine N-acetyltransferase 1 gene (TcAANAT1) is required for cuticle morphology and pigmentation of the adult red flour beetle, Tribolium castaneum. Insect Biochem. Mol. Biol. 2016, 79, 119–129. [Google Scholar] [CrossRef]
- Masuoka, Y.; Maekawa, K. Ecdysone signaling regulates soldier-specific cuticular pigmentation in the termite Zootermopsis nevadensis. FEBS Lett. 2016, 590, 1694–1703. [Google Scholar] [CrossRef]
- Dai, F.; Qiao, L.; Cao, C.; Liu, X.; Tong, X.; He, S.; Hu, H.; Zhang, L.; Wu, S.; Tan, D.; et al. Aspartate decarboxylase is required for a normal pupa pigmentation pattern in the silkworm, Bombyx mori. Sci. Rep. 2015, 5, 10885. [Google Scholar] [CrossRef]
- Phillips, A.M.; Smart, R.; Strauss, R.; Brembs, B.; Kelly, L.E. The Drosophila black enigma: The molecular and behavioural characterization of the black 1 mutant allele. Gene 2005, 351, 131–142. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kramer, K.J.; Morgan, T.D.; Hopkins, T.L.; Roseland, C.R.; Aso, Y.; Beeman, R.W.; Lookhart, G. Catecholamines and β-alanine in the red flour beetle, Tribolium castaneum: Roles in cuticle sclerotization and melanization. Insect Biochem. 1984, 14, 293–298. [Google Scholar] [CrossRef]
- Arakane, Y.; Lomakin, J.; Beeman, R.W.; Muthukrishnan, S.; Gehrke, S.H.; Kanost, M.R.; Kramer, K.J. Molecular and functional analyses of amino acid decarboxylases involved in cuticle tanning in Tribolium castaneum. J. Biol. Chem. 2009, 284, 16584–16594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Futahashi, R.; Sato, J.; Meng, Y.; Okamoto, S.; Daimon, T.; Yamamoto, K.; Suetsugu, Y.; Narukawa, J.; Takahashi, H.; Banno, Y.; et al. yellow and ebony are the responsible genes for the larval color mutants of the silkworm Bombyx mori. Genetics 2008, 180, 1995–2005. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bridges, C.B.; Morgan, T.H. The Third-Chromosome Group of Mutant Characters of Drosophila melanogaster; Carnegie Institution of Washington: Washington, DC, USA, 1923. [Google Scholar]
- Bi, H.L.; Xu, J.; He, L.; Zhang, Y.; Li, K.; Huang, Y.P. CRISPR/Cas9-mediated ebony knockout results in puparium melanism in Spodoptera litura. Insect Sci. 2019, 26, 1011–1019. [Google Scholar] [CrossRef]
- Wittkopp, P.J.; True, J.R.; Carroll, S.B. Reciprocal functions of the Drosophila yellow and ebony proteins in the development and evolution of pigment patterns. Development 2002, 129, 1849–1858. [Google Scholar] [CrossRef]
- Mun, S.; Noh, M.Y.; Kramer, K.J.; Muthukrishnan, S.; Arakane, Y. Gene functions in adult cuticle pigmentation of the yellow mealworm, Tenebrio molitor. Insect Biochem. Mol. Biol. 2020, 117, 103291. [Google Scholar] [CrossRef]
- Tomoyasu, Y.; Arakane, Y.; Kramer, K.J.; Denell, R.E. Repeated co-options of exoskeleton formation during wing-to-elytron evolution in beetles. Curr. Biol. 2009, 19, 2057–2065. [Google Scholar] [CrossRef] [Green Version]
- Ferguson, L.C.; Maroja, L.; Jiggins, C.D. Convergent, modular expression of ebony and tan in the mimetic wing patterns of Heliconius butterflies. Dev. Genes Evol. 2011, 221, 297–308. [Google Scholar] [CrossRef]
- True, J.R.; Yeh, S.D.; Hovemann, B.T.; Kemme, T.; Meinertzhagen, I.A.; Edwards, T.N.; Liou, S.R.; Han, Q.; Li, J. Drosophila tan encodes a novel hydrolase required in pigmentation and vision. PLoS Genet. 2005, 1, e63. [Google Scholar] [CrossRef]
- Jeong, S.; Rebeiz, M.; Andolfatto, P.; Werner, T.; True, J.; Carroll, S.B. The evolution of gene regulation underlies a morphological difference between two Drosophila sister species. Cell 2008, 132, 783–793. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Qiao, L.; Du, M.; Liang, X.; Hao, Y.; He, X.; Si, F.; Mei, T.; Chen, B. Tyrosine hydroxylase is crucial for maintaining pupal tanning and immunity in Anopheles sinensis. Sci. Rep. 2016, 6, 29835. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, K.S.; Kim, B.Y.; Jin, B.R. Differential regulation of tyrosine hydroxylase in cuticular melanization and innate immunity in the silkworm Bombyx mori. J. Asia-Pac. Entomol. 2015, 18, 765–770. [Google Scholar] [CrossRef]
- Gorman, M.J.; An, C.J.; Kanost, M.R. Characterization of tyrosine hydroxylase from Manduca sexta. Insect Biochem. Mol. Biol. 2007, 37, 1327–1337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, S.; Wang, M.; Li, X. Overexpression of tyrosine hydroxylase and dopa decarboxylase associated with pupal melanization in Spodoptera exigua. Sci. Rep. 2015, 5, 11273. [Google Scholar] [CrossRef] [Green Version]
- Gorman, M.J.; Arakane, Y. Tyrosine hydroxylase is required for cuticle sclerotization and pigmentation in Tribolium castaneum. Insect Biochem. Mol. Biol. 2010, 40, 267–273. [Google Scholar] [CrossRef] [Green Version]
- Li, M.; Bui, M.; Yang, T.; Bowman, C.S.; White, B.J.; Akbari, O.S. Germline Cas9 expression yields highly efficient genome engineering in a major worldwide disease vector, Aedes aegypti. Proc. Nat. Acad. Sci. USA 2017, 114, E10540–E10549. [Google Scholar] [CrossRef] [Green Version]
- Suzuki, T.K.; Koshikawa, S.; Kobayashi, I.; Uchino, K.; Sezutsu, H. Modular cis-regulatory logic of yellow gene expression in silkmoth larvae. Insect Mol. Biol. 2019, 28, 568–577. [Google Scholar] [CrossRef] [Green Version]
- Wang, P.; Ze, L.; Jin, L.; Li, G.-Q. Involvement of Yellow-y in the cuticle pigmentation of the larvae, pupae and adults in Henosepilachna vigintioctopunctata. J. Asia-Pac. Entomol. 2022, 25, 101864. [Google Scholar] [CrossRef]
- Heinze, S.D.; Kohlbrenner, T.; Ippolito, D.; Meccariello, A.; Burger, A.; Mosimann, C.; Saccone, G.; Bopp, D. CRISPR-Cas9 targeted disruption of the yellow ortholog in the housefly identifies the brown body locus. Sci. Rep. 2017, 7, 4582. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, H.U.; Du, J.; Zhang, J.; Shen, J.; Cai, W. Three melanin pathway genes, TH, yellow, and aaNAT, regulate pigmentation in the twin-spotted assassin bug, Platymeris biguttatus (Linnaeus). Int. J. Mol. Sci. 2019, 20, 2728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arakane, Y.; Dittmer, N.T.; Tomoyasu, Y.; Kramer, K.J.; Muthukrishnan, S.; Beeman, R.W.; Kanost, M.R. Identification, mRNA expression and functional analysis of several yellow family genes in Tribolium castaneum. Insect Biochem. Mol. Biol. 2010, 40, 259–266. [Google Scholar] [CrossRef] [PubMed]
- Rylee, J.C.; Siniard, D.J.; Doucette, K.K.; Zentner, G.E.; Zelhof, A.C. Expanding the genetic toolkit of Tribolium castaneum. PLoS ONE 2018, 13, e01895977. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Debolt, J.W. Meridic diet for rearing successive generations of Lygus hesperus. Ann. Entomol. Soc. Am. 1982, 75, 119–122. [Google Scholar] [CrossRef]
- Patana, R. Disposable diet packet for feeding and oviposition of Lygus hesperus (Hemiptera: Miridae). J. Econ. Entomol. 1982, 75, 668–669. [Google Scholar] [CrossRef]
- Basic Local Alignment Search Tool. Available online: http://blast.ncbi.nlm.nih.gov/Blast.cgi (accessed on 13 September 2022).
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef] [Green Version]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Stecher, G.; Tamura, K.; Kumar, S. Molecular Evolutionary Genetics Analysis (MEGA) for macOS. Mol. Biol. Evol. 2020, 37, 1237–1239. [Google Scholar] [CrossRef]
- Le, S.Q.; Gascuel, O. An improved general amino acid replacement matrix. Mol. Biol. Evol. 2008, 25, 1307–1320. [Google Scholar] [CrossRef] [Green Version]
- Saitou, N.; Nei, M. The neighbor-joining method: A new method for reconstructing phylogenetic trees. Mol. Biol. Evol. 1987, 4, 406–425. [Google Scholar]
- Liu, P.Y.; Ding, H.Z.; Christensen, B.M.; Li, J.Y. Cysteine sulfinic acid decarboxylase activity of Aedes aegypti aspartate 1-decarboxylase: The structural basis of its substrate selectivity. Insect Biochem. Mol. Biol. 2012, 42, 396–403. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.M.; Zheng, W.H.; Ye, W.Q.; Wang, C.; Gao, Y.; Cui, W.J.; Zhou, Z. Characterization of cysteine sulfinic acid decarboxylase from Tribolium castaneum and its application in the production of beta-alanine. Appl. Microbiol. Biotechnol. 2019, 103, 9443–9453. [Google Scholar] [CrossRef] [PubMed]
- Saenko, S.V.; Jerónimo, M.A.; Beldade, P. Genetic basis of stage-specific melanism: A putative role for a cysteine sulfinic acid decarboxylase in insect pigmentation. Heredity 2012, 108, 594–601. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arakane, Y.; Muthukrishnan, S.; Beeman, R.W.; Kanost, M.R.; Kramer, K.J. Laccase 2 is the phenoloxidase gene required for beetle cuticle tanning. Proc. Nat. Acad. Sci. USA 2005, 102, 11337–11342. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsugehara, T.; Imai, T.; Takeda, M. Characterization of arylalkylamine N-acetyltransferase from silkmoth (Antheraea pernyi) and pesticidal drug design based on the baculovirus-expressed enzyme. Comp. Biochem. Physiol. C Toxicol. Pharm. 2013, 157, 93–102. [Google Scholar] [CrossRef]
- O’Flynn, B.G.; Lewandowski, E.M.; Prins, K.C.; Suarez, G.; McCaskey, A.N.; Rios-Guzman, N.M.; Anderson, R.L.; Shepherd, B.A.; Gelis, I.; Leahy, J.W.; et al. Characterization of arylalkylamine N-acyltransferase from Tribolium castaneum: An investigation into a potential next-generation insecticide target. ACS Chem. Biol. 2020, 15, 513–523. [Google Scholar] [CrossRef] [PubMed]
- Roseland, C.R.; Kramer, K.J.; Hopkins, T.L. Cuticular strength and pigmentation of rust-red and black strains of Tribolium castaneum. Correlation with catecholamine and beta-alanine content. Insect Biochem. 1987, 17, 21–28. [Google Scholar] [CrossRef]
- Verlinden, H. Dopamine signaling in locusts and other insects. Biology 2018, 97, 40–52. [Google Scholar]
- Brent, C.S.; Miyasaki, K.; Vuong, C.; Miranda, B.; Steele, B.; Brent, K.G.; Nath, R. Regulatory roles of biogenic amines and juvenile hormone in the reproductive behavior of the western tarnished plant bug (Lygus hesperus). J. Comp. Physiol. B 2016, 186, 169–179. [Google Scholar] [CrossRef]
- Zhang, L.; Martin, A.; Perry, M.W.; van der Burg, K.R.; Matsuoka, Y.; Monteiro, A.; Reed, R.D. Genetic basis of melanin pigmentation in butterfly wings. Genetics 2017, 205, 1537–1550. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Brent, C.S.; Heu, C.C.; Gross, R.J.; Fan, B.; Langhorst, D.; Hull, J.J. RNAi-Mediated Manipulation of Cuticle Coloration Genes in Lygus hesperus Knight (Hemiptera: Miridae). Insects 2022, 13, 986. https://doi.org/10.3390/insects13110986
Brent CS, Heu CC, Gross RJ, Fan B, Langhorst D, Hull JJ. RNAi-Mediated Manipulation of Cuticle Coloration Genes in Lygus hesperus Knight (Hemiptera: Miridae). Insects. 2022; 13(11):986. https://doi.org/10.3390/insects13110986
Chicago/Turabian StyleBrent, Colin S., Chan C. Heu, Roni J. Gross, Baochan Fan, Daniel Langhorst, and J. Joe Hull. 2022. "RNAi-Mediated Manipulation of Cuticle Coloration Genes in Lygus hesperus Knight (Hemiptera: Miridae)" Insects 13, no. 11: 986. https://doi.org/10.3390/insects13110986