
BmAbl1 Regulates Silk Protein Synthesis via Glutathione Metabolism in Bombyx mori

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Sample Preparation
2.2. Inhibited BmAbl1 Using Dasatinib
2.3. Measurement of Tyrosine Kinase Activity
2.4. cDNA Synthesis and Quantitative Real-Time PCR
2.5. Plasmid Construction
2.6. Silkworm Germline Transformation
2.7. RNA-Sequencing Analysis
2.8. Phenotypic Measurement of Transgenic Strain
2.9. Measurement Content of Glutathione (GSH)
2.10. Measurement Content of Free Amino Acids
2.11. Western Blot
2.12. Statistical Analysis
3. Results
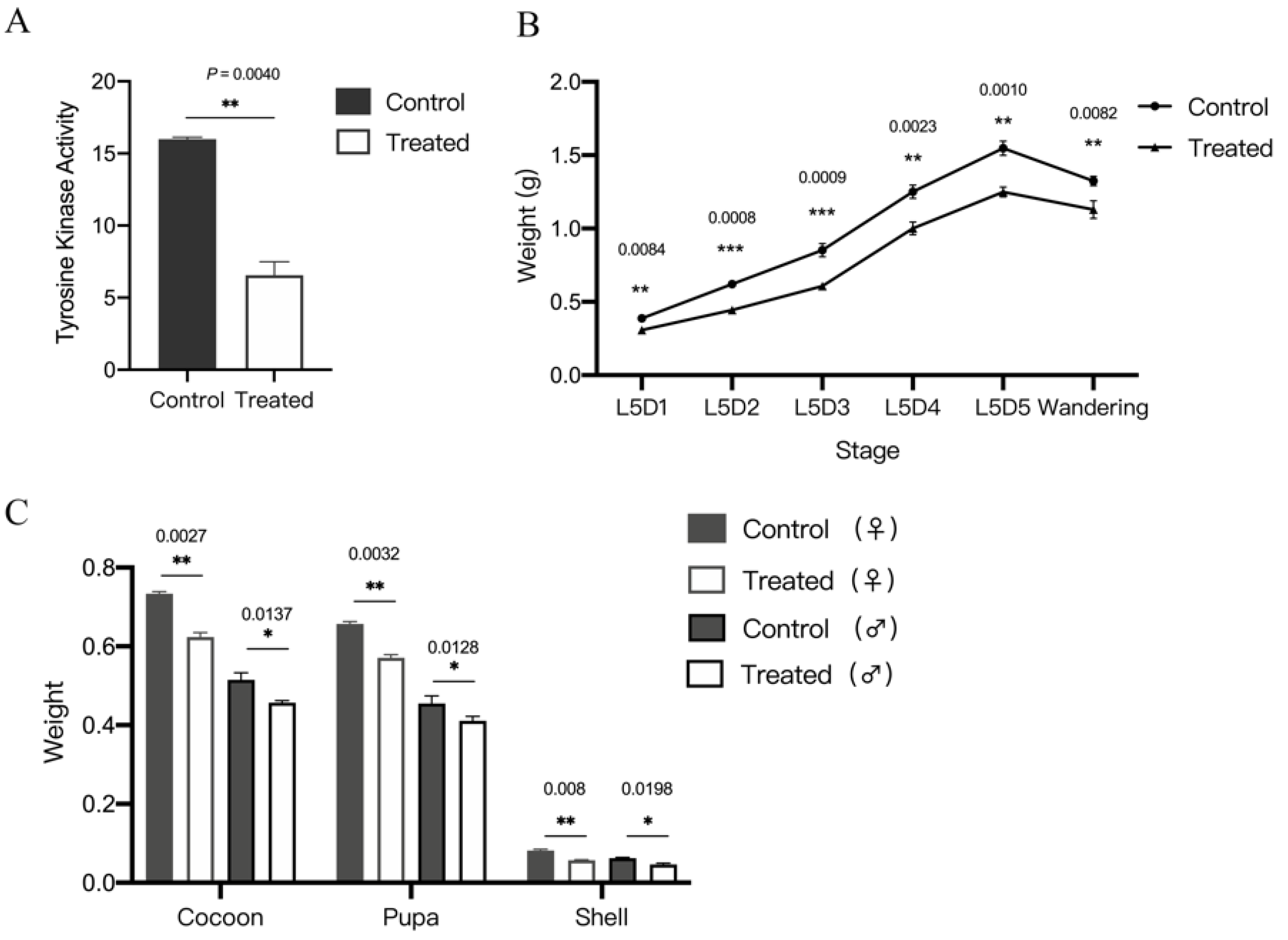
3.1. BmAbl1 Inhibited by Dasatinib Reduce Larva Weight and CSW
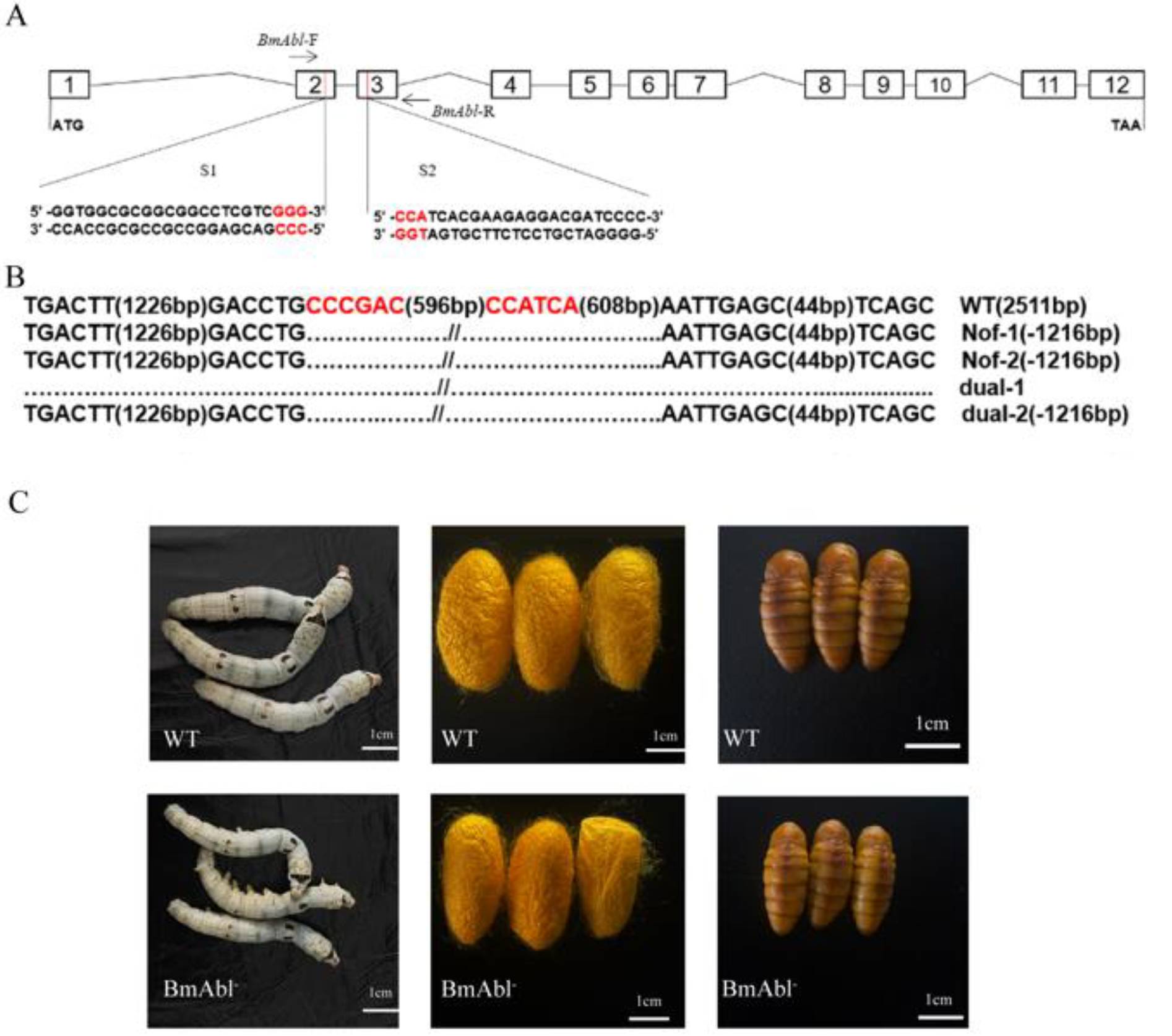
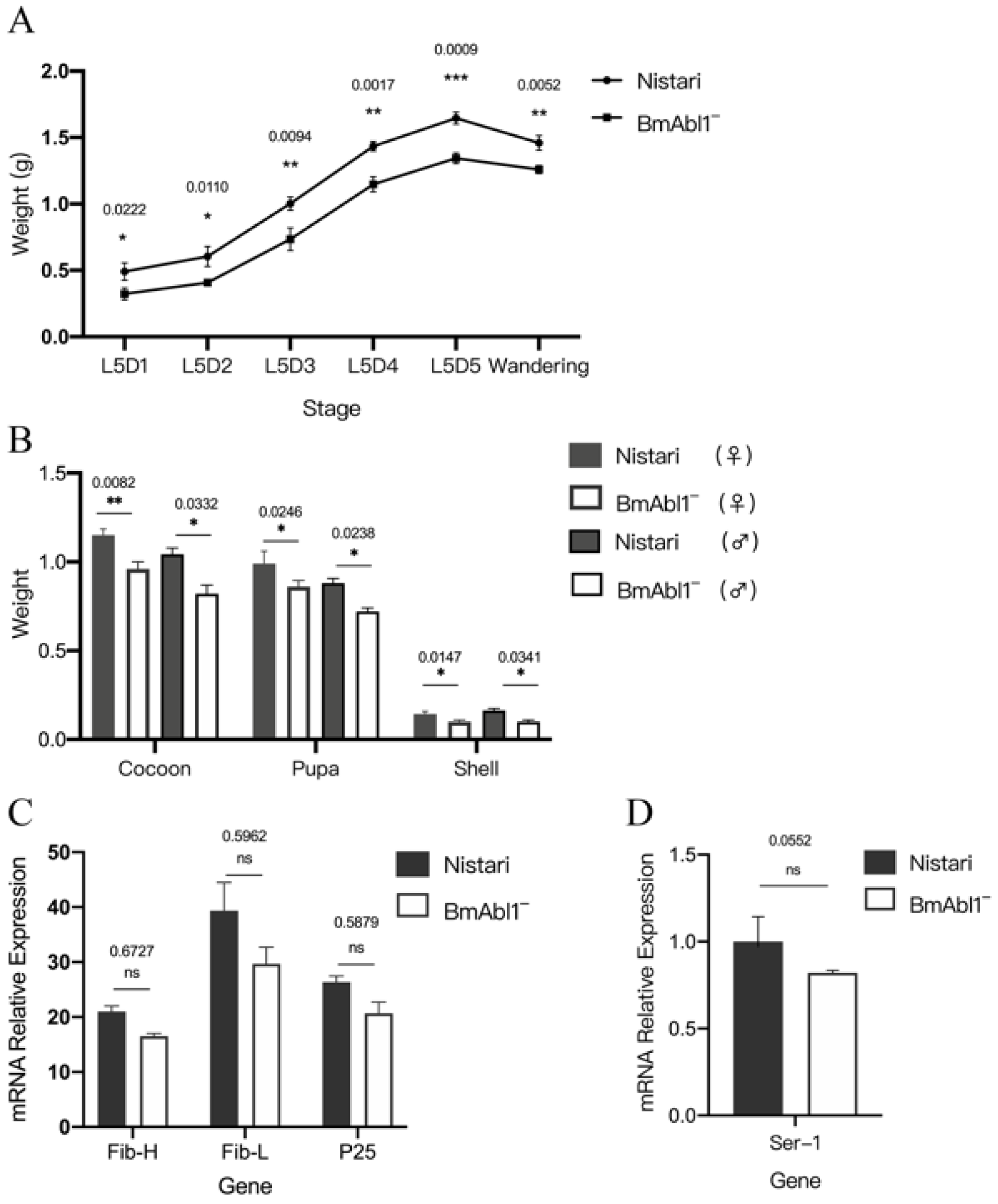
3.2. BmAbl1 Knockout Reduces Larva Weight and CSW
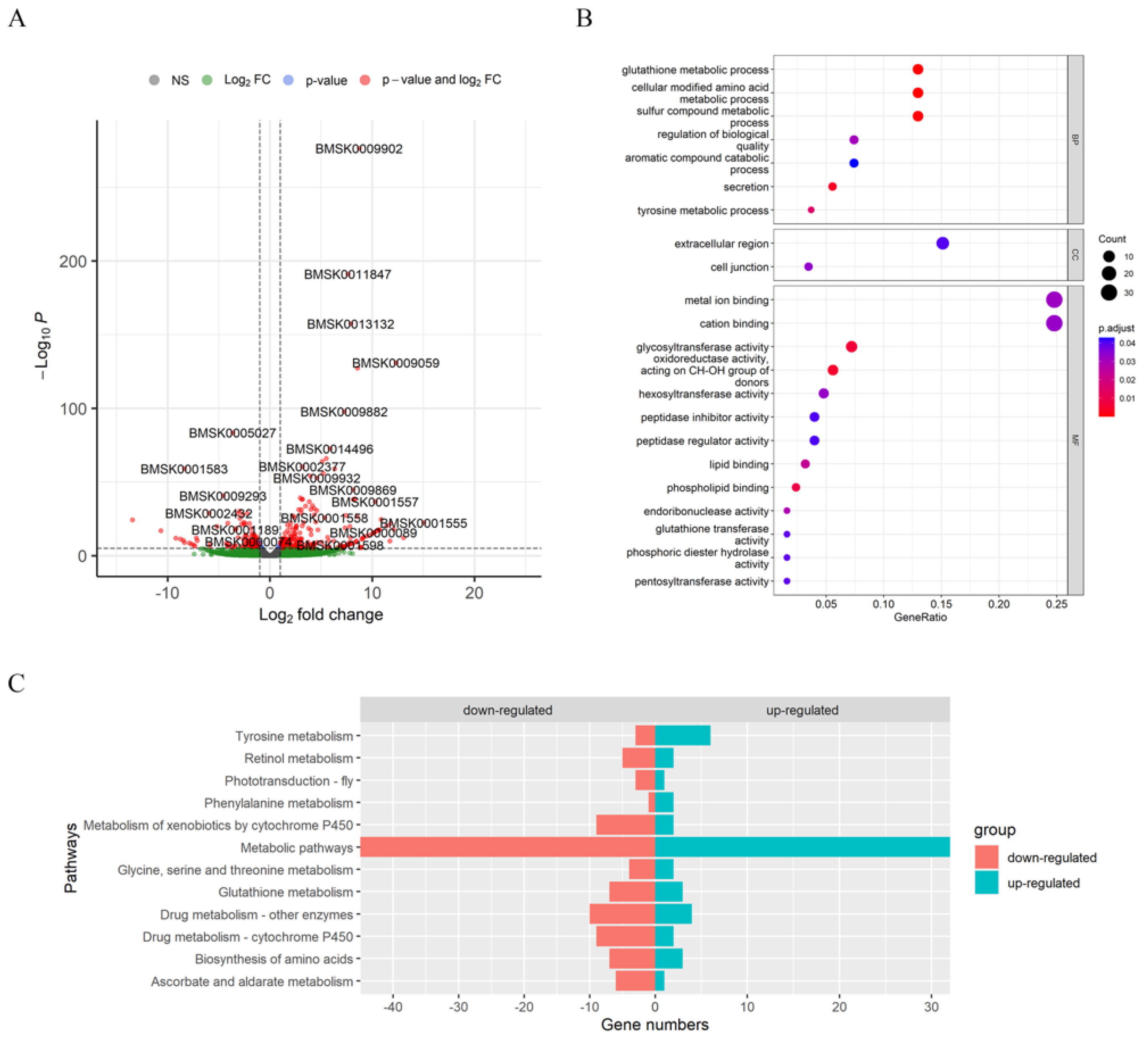
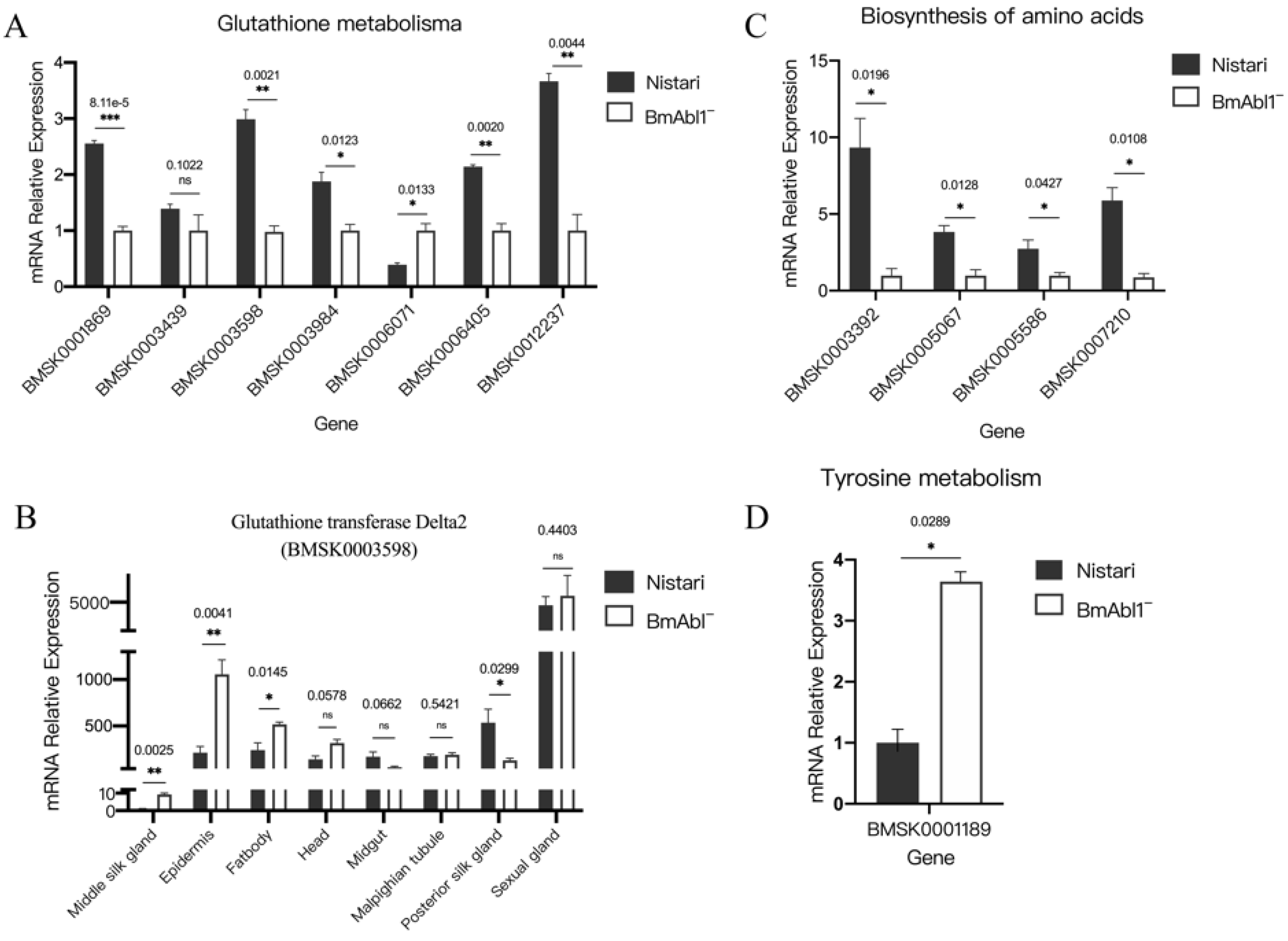
3.3. Glutathione Metabolism and Amino Acid Biosynthesis Were Affected by Knocking Out BmAbl1-
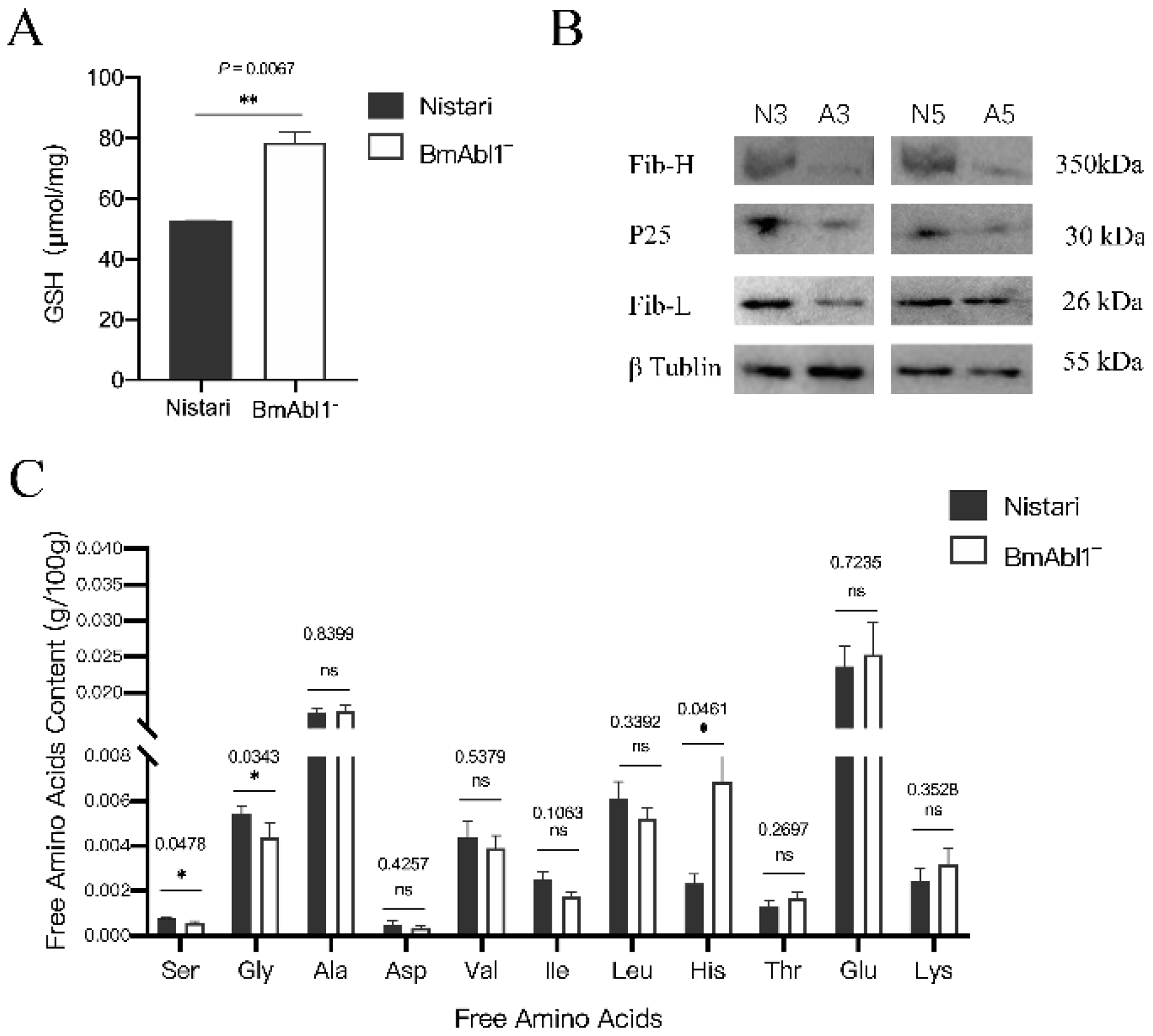
3.4. Abnormal Glutathione Metabolism Reduced Glycine and Serine Content in PSG of BmAbl1-
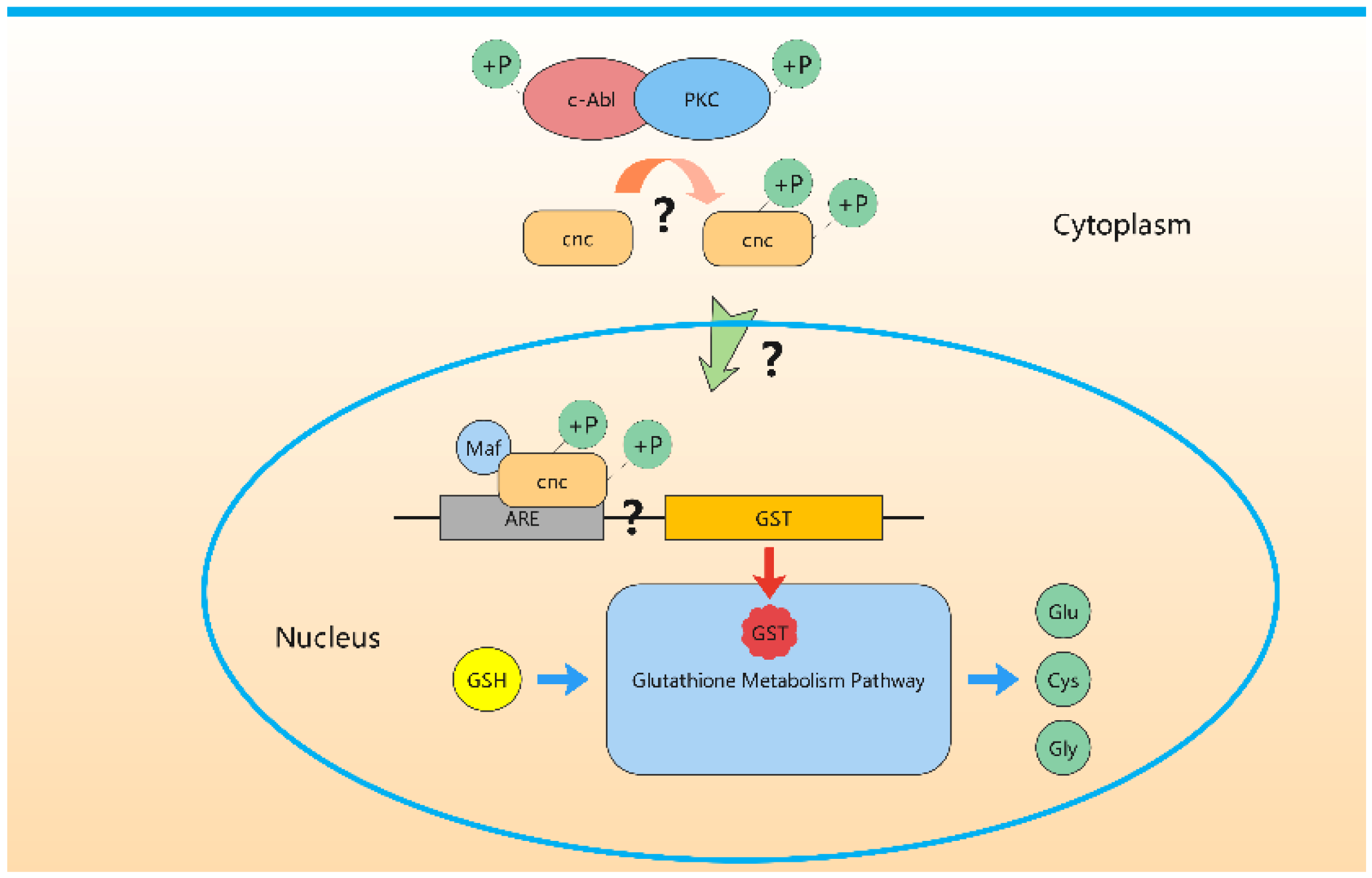
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Tanaka, K.; Kajiyama, N.; Ishikura, K.; Waga, S.; Kikuchi, A.; Ohtomo, K.; Takagi, T.; Mizuno, S. Determination of the site of disulfide linkage between heavy and light chains of silk fibroin produced by Bombyx mori. Biochim. Biophys. Acta (BBA)-Protein Struct. Mol. Enzymol. 1999, 1432, 92–103. [Google Scholar] [CrossRef]
- Yukuhiro, K.; Sezutsu, H.; Yonemura, N. Evolutionary Divergence of Lepidopteran and Trichopteran Fibroins. In Biotechnology of Silk; Springer: Berlin/Heidelberg, Germany, 2014; pp. 25–47. [Google Scholar]
- Inoue, S.; Tanaka, K.; Arisaka, F.; Kimura, S.; Ohtomo, K.; Mizuno, S. Silk fibroin of Bombyx mori is secreted, assembling a high molecular mass elementary unit consisting of H-chain, L-chain, and P25, with a 6: 6: 1 molar ratio. J. Biol. Chem. 2000, 275, 40517–40528. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, C.Z.; Confalonieri, F.; Jacquet, M.; Perasso, R.; Li, Z.G.; Janin, J. Silk fibroin: Structural implications of a remarkable amino acid sequence. Proteins Struct. Funct. Bioinform. 2001, 44, 119–122. [Google Scholar] [CrossRef] [PubMed]
- Bin, L.; Cheng, L.; Zhao, A.-C.; Xiang, Z.-H. Multiple interval mapping for whole cocoon weight and related economically important traits QTL in silkworm (Bombyx mori). Agric. Sci. China 2006, 5, 798–804. [Google Scholar]
- Zhan, S.; Huang, J.; Guo, Q.; Zhao, Y.; Li, W.; Miao, X.; Goldsmith, M.R.; Li, M.; Huang, Y. An integrated genetic linkage map for silkworms with three parental combinations and its application to the mapping of single genes and QTL. BMC Genom. 2009, 10, 389. [Google Scholar] [CrossRef] [Green Version]
- Li, B.; Wang, X.; Hou, C.; Xu, A.; Li, M. Genetic analysis of quantitative trait loci for cocoon and silk production quantity in Bombyx mori (Lepidoptera: Bombycidae). Eur. J. Entomol. 2013, 110, 205. [Google Scholar] [CrossRef] [Green Version]
- Li, C.; Zuo, W.; Tong, X.; Hu, H.; Qiao, L.; Song, J.; Xiong, G.; Gao, R.; Dai, F.; Lu, C. A composite method for mapping quantitative trait loci without interference of female achiasmatic and gender effects in silkworm, Bombyx mori. Anim. Genet. 2015, 46, 426–432. [Google Scholar] [CrossRef]
- He, S.; Tong, X.; Han, M.; Bai, Y.; Dai, F. Genome-wide identification and characterization of tyrosine kinases in the silkworm, Bombyx mori. Int. J. Mol. Sci. 2018, 19, 934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Luan, Y.; Li, C.; Zuo, W.; Hu, H.; Gao, R.; Zhang, B.; Tong, X.; Lu, C.; Dai, F. Gene mapping reveals the association between tyrosine protein kinase Abl1 and the silk yield of Bombyx mori. Anim. Genet. 2021, 52, 342–350. [Google Scholar] [CrossRef] [PubMed]
- Colicelli, J. ABL tyrosine kinases: Evolution of function, regulation, and specificity. Sci. Signal. 2010, 3, re6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bashaw, G.J.; Kidd, T.; Murray, D.; Pawson, T.; Goodman, C.S. Repulsive axon guidance: Abelson and Enabled play opposing roles downstream of the roundabout receptor. Cell 2000, 101, 703–715. [Google Scholar] [CrossRef]
- Yu, H.-H.; Zisch, A.H.; Dodelet, V.C.; Pasquale, E.B. Multiple signaling interactions of Abl and Arg kinases with the EphB2 receptor. Oncogene 2001, 20, 3995–4006. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crowner, D.; Le Gall, M.; Gates, M.A.; Giniger, E. Notch steers Drosophila ISNb motor axons by regulating the Abl signaling pathway. Curr. Biol. 2003, 13, 967–972. [Google Scholar] [CrossRef] [Green Version]
- Giniger, E. A role for Abl in Notch signaling. Neuron 1998, 20, 667–681. [Google Scholar] [CrossRef] [Green Version]
- Pendergast, A.M. The Abl family kinases: Mechanisms of regulation and signaling. Adv. Cancer Res. 2002, 85, 51–100. [Google Scholar]
- Hernández, S.E.; Krishnaswami, M.; Miller, A.L.; Koleske, A.J. How do Abl family kinases regulate cell shape and movement? Trends Cell Biol. 2004, 14, 36–44. [Google Scholar] [CrossRef]
- Rhee, J.; Buchan, T.; Zukerberg, L.; Lilien, J.; Balsamo, J. Cables links Robo-bound Abl kinase to N-cadherin-bound β-catenin to mediate Slit-induced modulation of adhesion and transcription. Nat. Cell Biol. 2007, 9, 883–892. [Google Scholar] [CrossRef]
- Steelman, L.; Pohnert, S.; Shelton, J.; Franklin, R.; Bertrand, F.; McCubrey, J. JAK/STAT, Raf/MEK/ERK, PI3K/Akt and BCR-ABL in cell cycle progression and leukemogenesis. Leukemia 2004, 18, 189–218. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Xu, H.; Zhu, J.; Ma, S.; Liu, Y.; Jiang, R.-J.; Xia, Q.; Li, S. Ras1CA overexpression in the posterior silk gland improves silk yield. Cell Res. 2011, 21, 934–943. [Google Scholar] [CrossRef] [Green Version]
- Ma, L.; Liu, S.; Shi, M.; Chen, X.-X.; Li, S. Ras1CA-upregulated BCPI inhibits cathepsin activity to prevent tissue destruction of the Bombyx posterior silk gland. J. Proteome Res. 2013, 12, 1924–1934. [Google Scholar] [CrossRef]
- Lu, F.; Wei, Z.; Luo, Y.; Guo, H.; Zhang, G.; Xia, Q.; Wang, Y. SilkDB 3.0: Visualizing and exploring multiple levels of data for silkworm. Nucleic Acids Res. 2020, 48, D749–D755. [Google Scholar] [CrossRef] [Green Version]
- Kim, D.; Paggi, J.M.; Park, C.; Bennett, C.; Salzberg, S.L. Graph-based genome alignment and genotyping with HISAT2 and HISAT-genotype. Nat. Biotechnol. 2019, 37, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Anders, S.; Pyl, P.T.; Huber, W. HTSeq—A Python framework to work with high-throughput sequencing data. Bioinformatics 2015, 31, 166–169. [Google Scholar] [CrossRef] [Green Version]
- Yu, G.; Wang, L.-G.; Han, Y.; He, Q.-Y. clusterProfiler: An R package for comparing biological themes among gene clusters. Omics A J. Integr. Biol. 2012, 16, 284–287. [Google Scholar] [CrossRef] [PubMed]
- Xie, C.; Mao, X.; Huang, J.; Ding, Y.; Wu, J.; Dong, S.; Kong, L.; Gao, G.; Li, C.-Y.; Wei, L. KOBAS 2.0: A web server for annotation and identification of enriched pathways and diseases. Nucleic Acids Res. 2011, 39 (Suppl. S2), W316–W322. [Google Scholar] [CrossRef] [Green Version]
- Xu, J.; Dong, Q.; Yu, Y.; Niu, B.; Ji, D.; Li, M.; Huang, Y.; Chen, X.; Tan, A. Mass spider silk production through targeted gene replacement in Bombyx mori. Proc. Natl. Acad. Sci. USA 2018, 115, 8757–8762. [Google Scholar] [CrossRef] [Green Version]
- Hou, S.; Tao, C.; Yang, H.; Cheng, T.; Liu, C. Sage controls silk gland development by regulating Dfd in Bombyx mori. Insect Biochem. Mol. Biol. 2021, 132, 103568. [Google Scholar] [CrossRef] [PubMed]
- Ma, L.; Ma, Q.; Li, X.; Cheng, L.; Li, K.; Li, S. Transcriptomic analysis of differentially expressed genes in the Ras1(CA)-overexpressed and wildtype posterior silk glands. BMC Genom. 2014, 15, 182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, Y.F.; Chen, X.Y.; Zhang, C.D.; Tang, X.F.; Wang, L.; Liu, T.H.; Pan, M.H.; Lu, C. Effects of starvation and hormones on DNA synthesis in silk gland cells of the silkworm, Bombyx mori. Insect Sci. 2016, 23, 569–578. [Google Scholar] [CrossRef]
- Ikeda, H.; Nishi, S.; Sakai, M. Transcription factor Nrf2/MafK regulates rat placental glutathione S-transferase gene during hepatocarcinogenesis. Biochem. J. 2004, 380, 515–521. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, A.; Kang, M.-I.; Watai, Y.; Tong, K.I.; Shibata, T.; Uchida, K.; Yamamoto, M. Oxidative and electrophilic stresses activate Nrf2 through inhibition of ubiquitination activity of Keap1. Mol. Cell. Biol. 2006, 26, 221–229. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Consortium, m.; Roy, S.; Ernst, J.; Kharchenko, P.V.; Kheradpour, P.; Negre, N.; Eaton, M.L.; Landolin, J.M.; Bristow, C.A.; Ma, L. Identification of functional elements and regulatory circuits by Drosophila modENCODE. Science 2010, 330, 1787–1797. [Google Scholar] [CrossRef] [PubMed]
- Misra, J.R.; Horner, M.A.; Lam, G.; Thummel, C.S. Transcriptional regulation of xenobiotic detoxification in Drosophila. Genes Dev. 2011, 25, 1796–1806. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, X.; Wu, F.; Datta, R.; Kharbanda, S.; Kufe, D. Interaction between protein kinase C δ and the c-Abl tyrosine kinase in the cellular response to oxidative stress. J. Biol. Chem. 2000, 275, 7470–7473. [Google Scholar] [CrossRef] [Green Version]
- Huang, H.-C.; Nguyen, T.; Pickett, C.B. Phosphorylation of Nrf2 at Ser-40 by protein kinase C regulates antioxidant response element-mediated transcription. J. Biol. Chem. 2002, 277, 42769–42774. [Google Scholar] [CrossRef]







Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, S.; Sun, L.; Zhang, S.; Sun, X.; Li, M. BmAbl1 Regulates Silk Protein Synthesis via Glutathione Metabolism in Bombyx mori. Insects 2022, 13, 967. https://doi.org/10.3390/insects13110967
Qin S, Sun L, Zhang S, Sun X, Li M. BmAbl1 Regulates Silk Protein Synthesis via Glutathione Metabolism in Bombyx mori. Insects. 2022; 13(11):967. https://doi.org/10.3390/insects13110967
Chicago/Turabian StyleQin, Sheng, Lingling Sun, Shu Zhang, Xia Sun, and Muwang Li. 2022. "BmAbl1 Regulates Silk Protein Synthesis via Glutathione Metabolism in Bombyx mori" Insects 13, no. 11: 967. https://doi.org/10.3390/insects13110967