

Community of Bark and Ambrosia Beetles (Coleoptera: Curculionidae: Scolytinae and Platypodinae) in Agricultural and Forest Ecosystems with Laurel Wilt
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Lures
2.2. Trap Design
2.3. Field Tests
2.4. Statistical Analysis
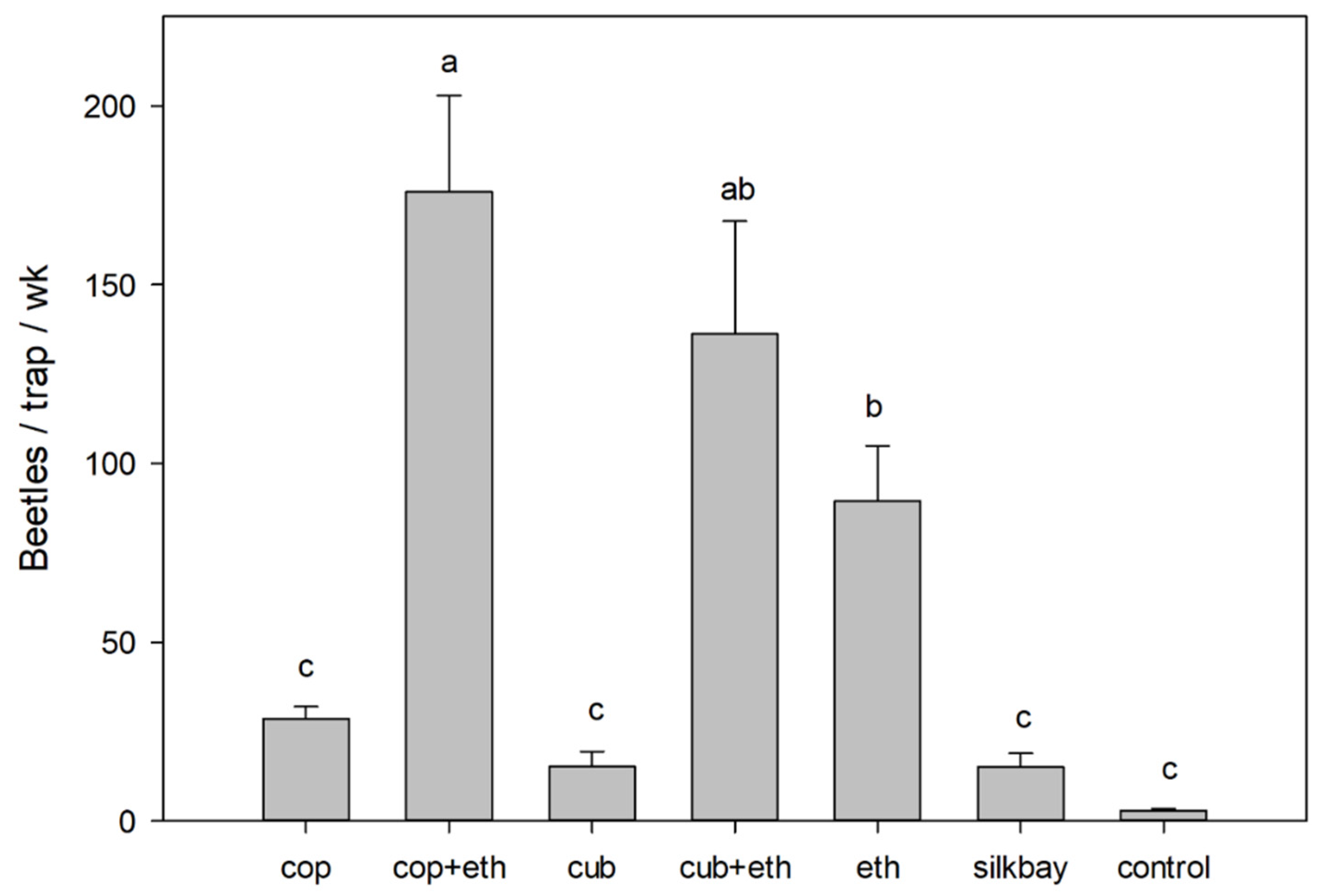
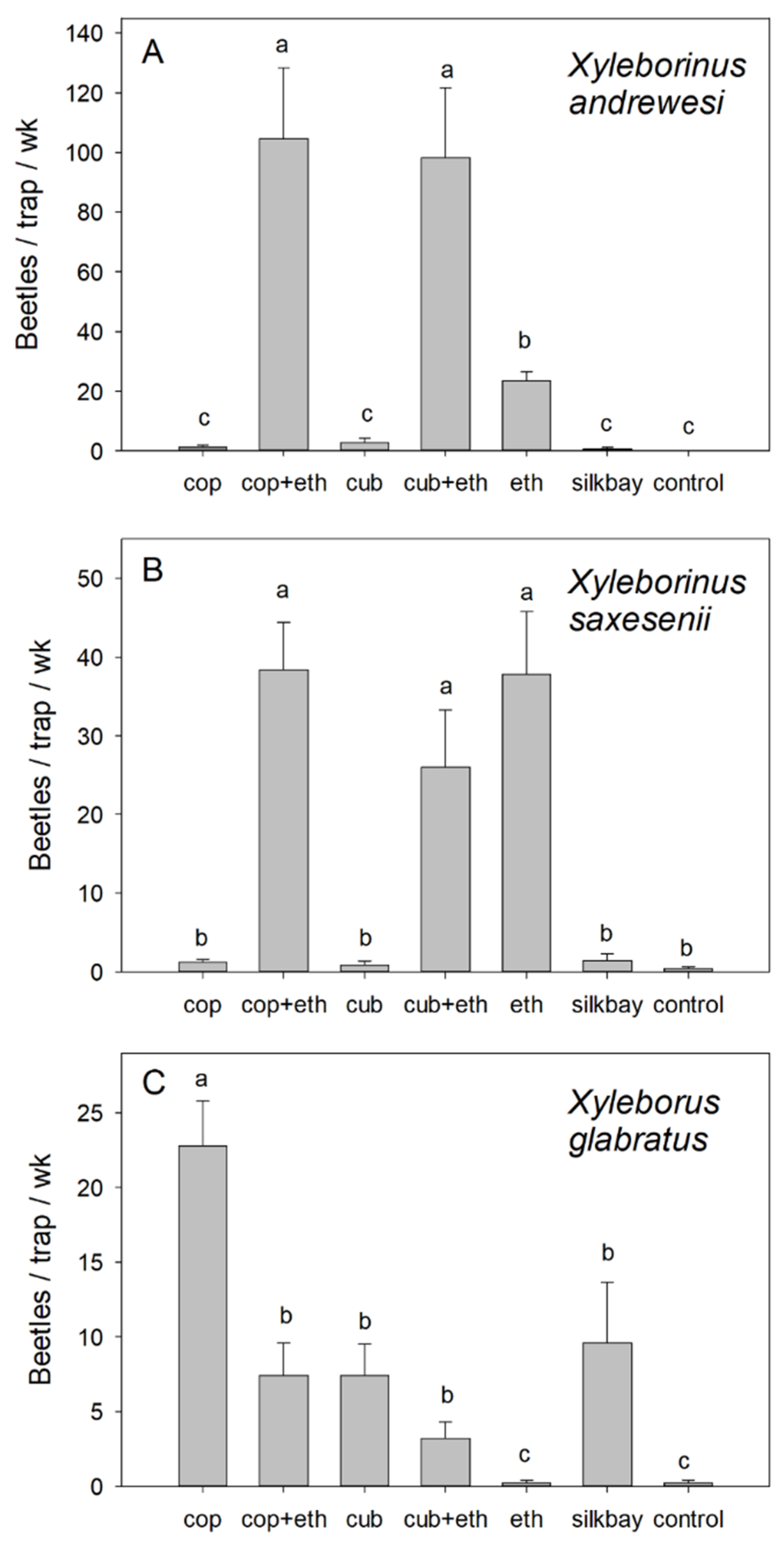
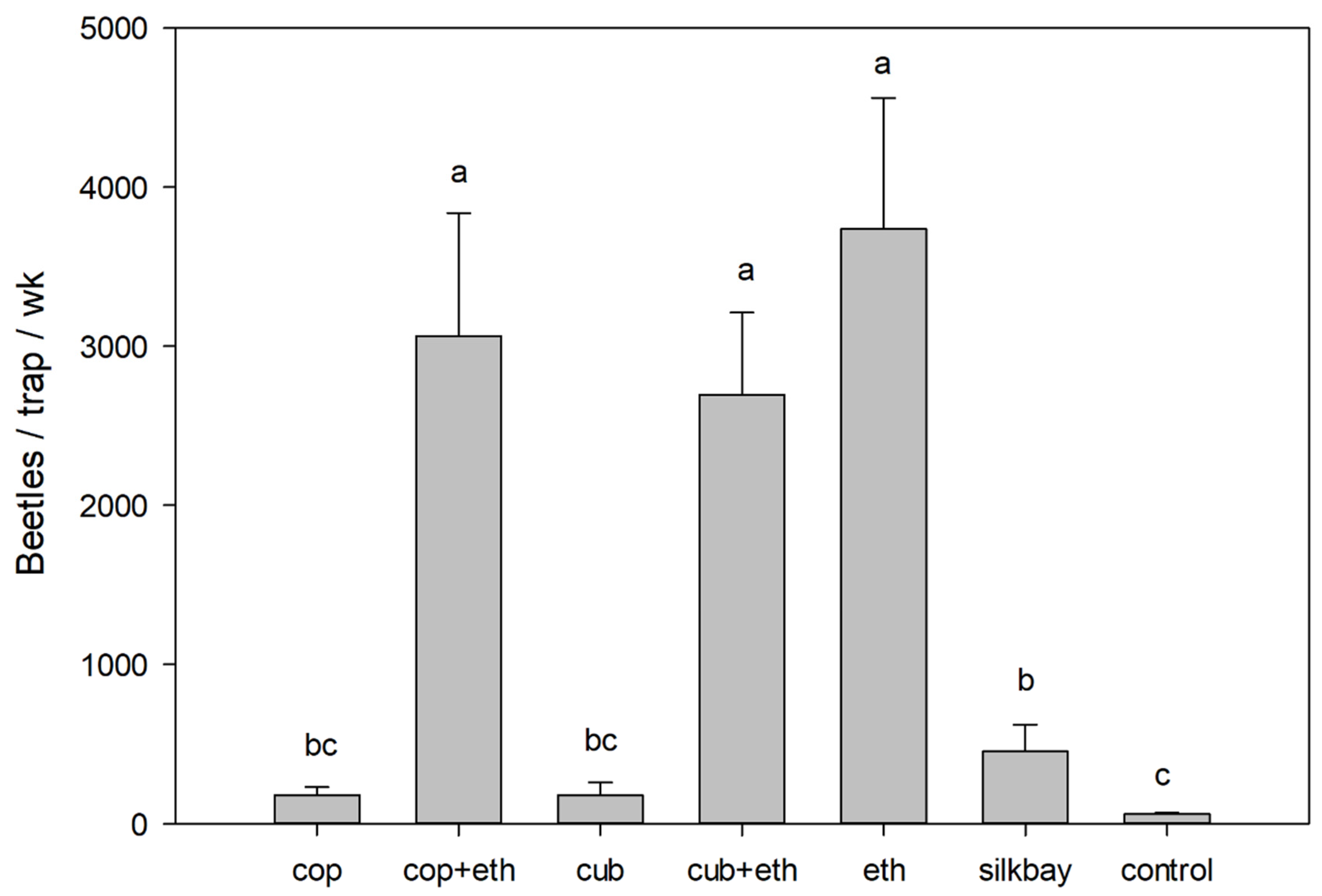
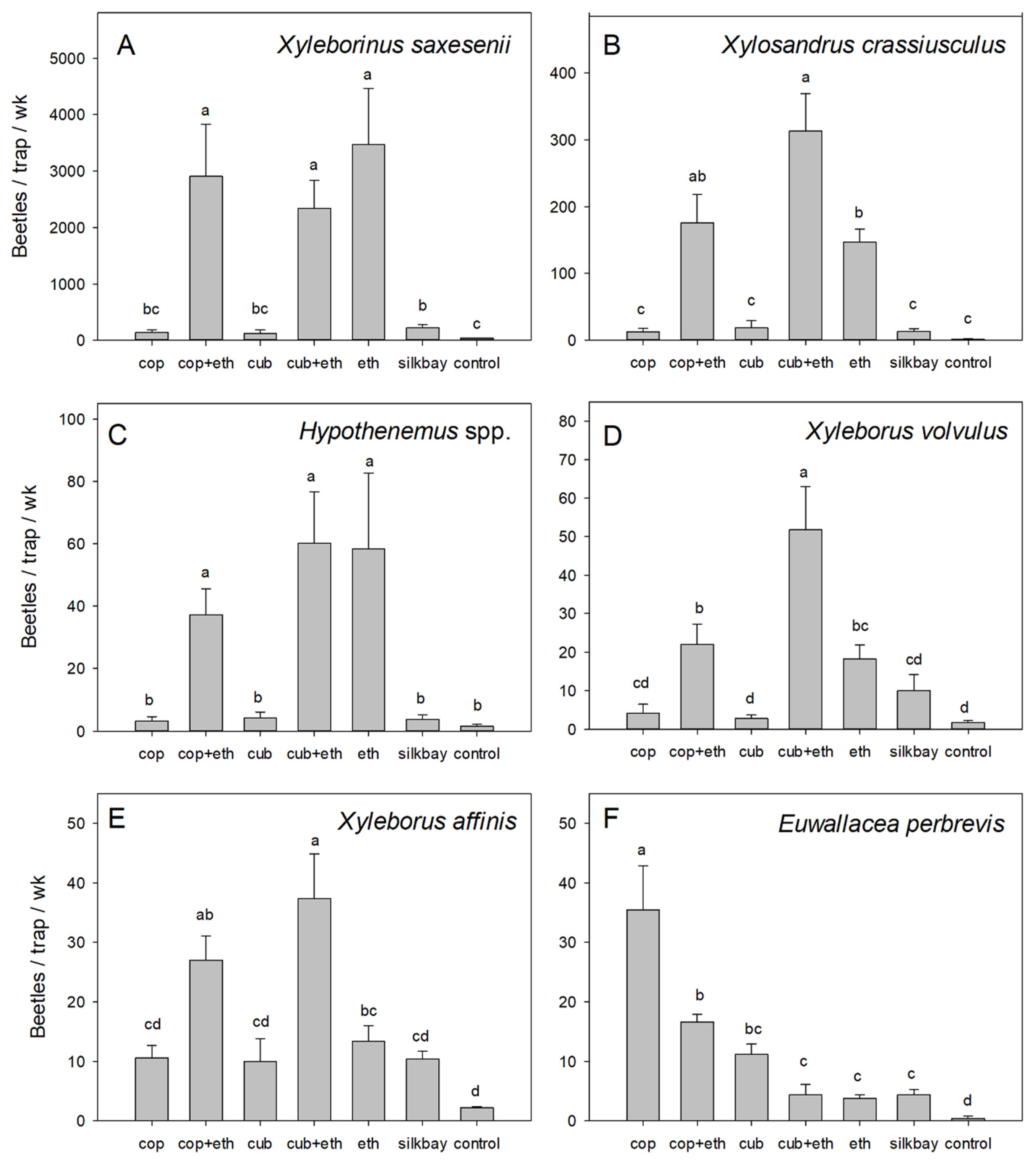
3. Results
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fraedrich, S. California laurel is susceptible to laurel wilt caused by Raffaelea lauricola. Plant Dis. 2008, 92, 1469. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrington, T.; Fraedrich, S.; Aghayeva, D. Raffaelea lauricola, a new ambrosia beetle symbiont and pathogen on the Lauracea. Mycotaxon 2008, 104, 399–404. [Google Scholar]
- de Beer, Z.; Proctor, M.; Wingfield, M.; Marincowitz, S.; Duong, T. Generic boundaries in the Ophiostomatales reconsidered and revised. Stud. Mycol. 2022, 101, 57–120. [Google Scholar] [CrossRef] [PubMed]
- Hughes, M.A.; Riggins, J.J.; Koch, F.H.; Cognato, A.I.; Anderson, C.; Formby, J.P.; Dreaden, T.J.; Ploetz, R.C.; Smith, J.A. No rest for the laurels: Symbiotic invaders cause unprecedented damage to southern USA forests. Biol. Invasions 2017, 19, 2143–2157. [Google Scholar] [CrossRef]
- Fraedrich, S.; Harrington, T.; Rabaglia, R.; Ulyshen, M.; Mayfield, A., III; Hanula, J.; Eickwort, J.; Miller, D. A fungal symbiont of the redbay ambrosia beetle causes a lethal wilt in redbay and other Lauraceae in the southeastern United States. Plant Dis. 2008, 92, 215–224. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ploetz, R.C.; Kendra, P.E.; Choudhury, R.A.; Rollins, J.A.; Campbell, A.; Garrett, K.; Hughes, M.; Dreaden, T. Laurel wilt in natural and agricultural ecosystems: Understanding the drivers and scales of complex pathosystems. Forests 2017, 8, 48. [Google Scholar] [CrossRef]
- Castillo-Argaez, R.; Vazquez, A.; Konkol, J.L.; Vargas, A.I.; Ploetz, R.C.; Etxeberria, E.; Schaffer, B. Sap flow, xylem anatomy and photosynthetic variables of three Persea species in response to laurel wilt. Tree Physiol. 2020, 41, 1004–1018. [Google Scholar] [CrossRef]
- Inch, S.; Ploetz, R.; Blanchette, R.; Held, B. Histological and ultrastructural changes in avocado (Persea americana) induced by Raffaelea lauricola. Phytopathology 2011, 101, S78. [Google Scholar] [CrossRef] [Green Version]
- Hanula, J.L.; Mayfield, A.E., III; Fraedrich, S.W.; Rabaglia, R.J. Biology and host associations of redbay ambrosia beetle (Coleoptera: Curculionidae: Scolytinae), exotic vector of laurel wilt killing redbay trees in the southeastern United States. J. Econ. Entomol. 2008, 101, 1276–1286. [Google Scholar] [CrossRef]
- Mayfield, A.E., III; Hanula, J.L. Effect of tree species and end seal on attractiveness and utility of cut bolts to the redbay ambrosia beetle and granulate ambrosia beetle (Coleoptera: Curculionidae: Scolytinae). J. Econ. Entomol. 2012, 105, 461–470. [Google Scholar] [CrossRef] [Green Version]
- Rodgers, L.; Derksen, A.; Pernas, T. Expansion and impact of laurel wilt in the Florida Everglades. Fla. Entomol. 2014, 97, 1247–1250. [Google Scholar] [CrossRef]
- Hughes, M.; Shin, K.; Eickwort, J.; Smith, J. First report of laurel wilt disease caused by Raffaelea lauricola on silk bay in Florida. Plant Dis. 2012, 96, 910. [Google Scholar] [CrossRef] [PubMed]
- Mayfield, A.; Peña, J.; Crane, J.; Smith, J.; Branch, C.; Ottoson, E.; Hughes, M. Ability of the redbay ambrosia beetle (Coleoptera: Curculionidae: Scolytinae) to bore into young avocado (Lauraceae) plants and transmit the laurel wilt pathogen (Raffaelea sp.). Fla. Entomol. 2008, 91, 485–487. [Google Scholar] [CrossRef]
- Mayfield, A.E., III; MacKenzie, M.; Cannon, P.G.; Oak, S.W.; Horn, S.; Hwang, J.; Kendra, P.E. Suitability of California bay laurel and other species as hosts for the non-native redbay ambrosia beetle and granulate ambrosia beetle. Agric. For. Entomol. 2013, 15, 227–235. [Google Scholar] [CrossRef]
- Fraedrich, S.; Harrington, T.; Bates, C.; Johnson, J.; Reid, L.; Best, G.S.; Leininger, T.; Hawkins, T. Susceptibility to laurel wilt and disease incidence in two rare plant species, pondberry and pondspice. Plant Dis. 2011, 95, 1056–1062. [Google Scholar] [CrossRef] [Green Version]
- Hughes, M.; Smith, J.; Mayfield, A., III; Minno, M.; Shin, K. First report of laurel wilt disease caused by Raffaelea lauricola on pondspice in Florida. Plant Dis. 2011, 95, 1588. [Google Scholar] [CrossRef]
- Hughes, M.; Black, A.; Smith, J.A. First report of laurel wilt caused by Raffaelea lauricola on bay laurel (Laurus nobilis) in the United States. Plant Dis. 2014, 98, 1159. [Google Scholar] [CrossRef]
- Ploetz, R.C.; Konkol, J. First report of Gulf Licaria, Licaria trianda, as a suscept of laurel wilt. Plant Dis. 2013, 97, 1248. [Google Scholar] [CrossRef]
- Fraedrich, S.; Harrington, T.; Best, G. Xyleborus glabratus attacks and systemic colonization by Raffaelea lauricola associated with dieback of Cinnamomum camphora in the southeastern United States. For. Pathol. 2015, 45, 60–70. [Google Scholar] [CrossRef]
- Smith, J.; Mount, L.; Mayfield, A., III; Bates, C.; Lamborn, W.; Fraedrich, S. First report of laurel wilt disease caused by Raffaelea lauricola on camphor in Florida and Georgia. Plant Dis. 2009, 93, 198. [Google Scholar] [CrossRef]
- Beaver, R.; Liu, L. An annotated synopsis of Taiwanese bark and ambrosia beetles, with new synonymy, new combinations and new records (Coleoptera: Curculionidae: Scolytinae). Zootaxa 2010, 2602, 1–47. [Google Scholar] [CrossRef]
- Haack, R.A. Exotic bark-and wood-boring Coleoptera in the United States: Recent establishments and interceptions. Can. J. For. Res. 2006, 36, 269–288. [Google Scholar] [CrossRef]
- Rabaglia, R.J.; Dole, S.A.; Cognato, A.I. Review of American Xyleborina (Coleoptera: Curculionidae: Scolytinae) occurring north of Mexico, with an illustrated key. Ann. Entomol. Soc. Am. 2006, 99, 1034–1056. [Google Scholar] [CrossRef]
- Bates, C.; Reid, L.; Trickel, R.; Eickwort, J.; Riggins, J.; Stone, D. Distribution of Counties with Laurel Wilt Disease by Year of Initial Detection; USDA Forest Service, Forest Health Protection, Region-8: 2016. Available online: https://www.fdacs.gov/content/download/95555/file/laurel-wilt-distribution-map.pdf (accessed on 19 September 2022).
- Gazis, R.; DeWitt, K.; Johnson, L.; Chamberlin, L.; Kennedy, A.H.; Hansen, M.A.; Bush, E. First Report of Laurel Wilt Disease Caused by Raffaelea lauricola on Sassafras in Virginia. Plant Dis. 2022, PDIS-11-21-2616-PDN. [Google Scholar] [CrossRef] [PubMed]
- Koch, F.H.; Smith, W.D. Spatio-temporal analysis of Xyleborus glabratus (Coleoptera: Circulionidae: Scolytinae) invasion in eastern US forests. Environ. Entomol. 2008, 37, 442–452. [Google Scholar] [CrossRef]
- Loyd, A.; Chase, K.; Nielson, A.; Hoover, N.; Dreaden, T.; Mayfield, A.; Crocker, E.; Fraedrich, S. First report of laurel wilt caused by Raffaelea lauricola on Sassafras albidum in Tennessee and Kentucky. Plant Dis. 2020, 104, 567. [Google Scholar] [CrossRef]
- Mayfield, A.E.; Villari, C.; Hamilton, J.L.; Slye, J.; Langston, W.; Oten, K.; Fraedrich, S. First report of laurel wilt disease caused by Raffaelea lauricola on sassafras in North Carolina. Plant Dis. 2019, 103, 155–156. [Google Scholar] [CrossRef]
- Ploetz, R.C.; Peña, J.E.; Smith, J.A.; Dreaden, T.J.; Crane, J.H.; Schubert, T.; Dixon, W. Laurel wilt, caused by Raffaelea lauricola, is confirmed in Miami-Dade county, center of Florida’s commercial avocado production. Plant Dis. 2011, 95, 1589. [Google Scholar] [CrossRef]
- Goldberg, N.; Heine, J. A comparison of arborescent vegetation pre-(1983) and post-(2008) outbreak of the invasive species the Asian ambrosia beetle Xyleborus glabratus in a Florida maritime hammock. Plant Ecol. Divers. 2009, 2, 77–83. [Google Scholar] [CrossRef]
- Shields, J.; Jose, S.; Freeman, J.; Bunyan, M.; Celis, G.; Hagan, D.; Morgan, M.; Pieterson, E.C.; Zak, J. Short-term impacts of laurel wilt on redbay (Persea borbonia [L.] spreng.) in a mixed evergreen-deciduous forest in northern Florida. J. For. 2011, 109, 82–88. [Google Scholar] [CrossRef]
- Smith, C.K.; Landreaux, E.; Steinmann, H.; McGrath, D.; Hayes, C.; Hayes, R. Redbay survival eleven years after infection with an exotic disease on St. Catherines Island, Georgia, USA. Environ. Nat. Resour. Res. 2015, 6, 27. [Google Scholar] [CrossRef]
- Spiegel, K.S.; Leege, L.M. Impacts of laurel wilt disease on redbay (Persea borbonia (L.) Spreng.) population structure and forest communities in the coastal plain of Georgia, USA. Biol. Invasions 2013, 15, 2467–2487. [Google Scholar] [CrossRef]
- United States Department of Agriculture; National Agricultural Stastitics Servive. Noncitrus Fruits and Nuts 2020 Summary. 2020. Available online: https://downloads.usda.library.cornell.edu/usda-esmis/files/zs25x846c/sf269213r/6t054c23t/ncit0521.pdf (accessed on 19 September 2022).
- Ploetz, R.; Pérez-Martínez, J.; Smith, J.; Hughes, M.; Dreaden, T.; Inch, S.; Fu, Y. Responses of avocado to laurel wilt, caused by Raffaelea lauricola. Plant Pathol. 2012, 61, 801–808. [Google Scholar] [CrossRef]
- Ploetz, R.C.; Pérez-Martínez, J.M.; Evans, E.A.; Inch, S.A. Toward fungicidal management of laurel wilt of avocado. Plant Dis. 2011, 95, 977–982. [Google Scholar] [CrossRef] [Green Version]
- Evans, E.A.; Crane, J.; Hodges, A.; Osborne, J.L. Potential economic impact of laurel wilt disease on the Florida avocado industry. HortTechnology 2010, 20, 234–238. [Google Scholar] [CrossRef] [Green Version]
- Hughes, M.; Inch, S.; Ploetz, R.; Er, H.; van Bruggen, A.; Smith, J. Responses of swamp bay, Persea palustris, and avocado, Persea americana, to various concentrations of the laurel wilt pathogen, Raffaelea lauricola. For. Pathol. 2015, 45, 111–119. [Google Scholar] [CrossRef]
- Mosquera, M.; Evans, E.A.; Ploetz, R. Assessing the profitability of avocado production in south Florida in the presence of laurel wilt. Theor. Econ. Lett. 2015, 5, 343. [Google Scholar] [CrossRef] [Green Version]
- Carrillo, D.; Duncan, R.E.; Peña, J.E. Ambrosia beetles (Coleoptera: Curculionidae: Scolytinae) that breed in avocado wood in Florida. Fla. Entomol. 2012, 95, 573–579. [Google Scholar] [CrossRef]
- Kendra, P.E.; Montgomery, W.S.; Narvaez, T.I.; Carrillo, D. Comparison of trap designs for detection of Euwallacea nr. fornicatus and other Scolytinae (Coleoptera: Curculionidae) that vector fungal pathogens of avocado trees in Florida. J. Econ. Entomol. 2020, 113, 980–987. [Google Scholar] [CrossRef]
- Kendra, P.E.; Owens, D.; Montgomery, W.S.; Narvaez, T.I.; Bauchan, G.R.; Schnell, E.Q.; Tabanca, N.; Carrillo, D. α-Copaene is an attractant, synergistic with quercivorol, for improved detection of Euwallacea nr. fornicatus (Coleoptera: Curculionidae: Scolytinae). PLoS ONE 2017, 12, e0179416. [Google Scholar] [CrossRef]
- Menocal, O.; Kendra, P.E.; Montgomery, W.S.; Crane, J.H.; Carrillo, D. Vertical distribution and daily flight periodicity of ambrosia beetles (Coleoptera: Curculionidae) in Florida avocado orchards affected by laurel wilt. J. Econ. Entomol. 2018, 111, 1190–1196. [Google Scholar] [CrossRef] [PubMed]
- Carrillo, D.; Duncan, R.; Ploetz, J.; Campbell, A.; Ploetz, R.; Peña, J. Lateral transfer of a phytopathogenic symbiont among native and exotic ambrosia beetles. Plant Pathol. 2014, 63, 54–62. [Google Scholar] [CrossRef]
- Harrington, T.; Aghayeva, D.; Fraedrich, S. New combinations in Raffaelea, Ambrosiella, and Hyalorhinocladiella, and four new species from the redbay ambrosia beetle, Xyleborus glabratus. Mycotaxon 2010, 111, 337–361. [Google Scholar] [CrossRef] [Green Version]
- Harrington, T.; Fraedrich, S. Quantification of propagules of the laurel wilt fungus and other mycangial fungi from the redbay ambrosia beetle, Xyleborus glabratus. Phytopathology 2010, 100, 1118–1123. [Google Scholar] [CrossRef] [Green Version]
- Ploetz, R.C.; Konkol, J.L.; Narvaez, T.; Duncan, R.E.; Saucedo, R.J.; Campbell, A.; Mantilla, J.; Carrillo, D.; Kendra, P.E. Presence and prevalence of Raffaelea lauricola, cause of laurel wilt, in different species of ambrosia beetle in Florida, USA. J. Econ. Entomol. 2017, 110, 347–354. [Google Scholar]
- Baker, J.M.; Norris, D.M. A complex of fungi mutualistically involved in the nutrition of the ambrosia beetle Xyleborus ferrugineus. J. Invertebr. Pathol. 1968, 11, 246–250. [Google Scholar] [CrossRef]
- Gibson, C.M.; Hunter, M.S. Extraordinarily widespread and fantastically complex: Comparative biology of endosymbiotic bacterial and fungal mutualists of insects. Ecol. Lett. 2010, 13, 223–234. [Google Scholar] [CrossRef]
- Kolařík, M.; Hulcr, J. Mycobiota associated with the ambrosia beetle Scolytodes unipunctatus (Coleoptera: Curculionidae, Scolytinae). Mycol. Res. 2009, 113, 44–60. [Google Scholar] [CrossRef]
- Wood, S.L. The bark and ambrosia beetles of North and Central America (Coleoptera: Scolytidae), a taxonomic monograph. Brigh. Young Univ. 1982, 1, 1359. Available online: https://biostor.org/reference/239409 (accessed on 19 September 2022).
- Saucedo, J.; Ploetz, R.; Konkol, J.; Ángel, M.; Mantilla, J.; Menocal, O.; Carrillo, D. Nutritional symbionts of a putative vector, Xyleborus bispinatus, of the laurel wilt pathogen of avocado, Raffaelea lauricola. Symbiosis 2018, 75, 29–38. [Google Scholar] [CrossRef]
- Ploetz, R.C.; Konkol, J.; Pérez-Martínez, J.; Fernandez, R. Management of laurel wilt of avocado, caused by Raffaelea lauricola. Eur. J. Plant Pathol. 2017, 149, 133–143. [Google Scholar] [CrossRef]
- Dunn, J.P.; Potter, D.A. Synergistic effects of oak volatiles with ethanol in the capture of saprophagous wood borers. J. Entomol. Sci. 1991, 26, 425–429. [Google Scholar] [CrossRef]
- Ranger, C.M.; Schultz, P.B.; Frank, S.D.; Chong, J.H.; Reding, M.E. Non-native ambrosia beetles as opportunistic exploiters of living but weakened trees. PLoS ONE 2015, 10, e0131496. [Google Scholar] [CrossRef] [PubMed]
- Hanula, J.L.; Sullivan, B. Manuka oil and phoebe oil are attractive baits for Xyleborus glabratus (Coleoptera: Scolytinae), the vector of laurel wilt. Environ. Entomol. 2008, 37, 1403–1409. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kendra, P.E.; Montgomery, W.S.; Niogret, J.; Schnell, E.Q.; Deyrup, M.A.; Epsky, N.D. Evaluation of seven essential oils identifies cubeb oil as most effective attractant for detection of Xyleborus glabratus. J. Pest Sci. 2014, 87, 681–689. [Google Scholar] [CrossRef]
- Kendra, P.E.; Montgomery, W.S.; Niogret, J.; Peña, J.E.; Capinera, J.L.; Brar, G.; Epsky, N.D.; Heath, R.R. Attraction of the redbay ambrosia beetle, Xyleborus glabratus, to avocado, lychee, and essential oil lures. J. Chem. Ecol. 2011, 37, 932–942. [Google Scholar] [CrossRef]
- Niogret, J.; Epsky, N.D.; Schnell, R.J.; Boza, E.J.; Kendra, P.E.; Heath, R.R. Terpenoid variations within and among half-sibling avocado trees, Persea americana Mill. (Lauraceae). PLoS ONE 2013, 8, e73601. [Google Scholar] [CrossRef] [Green Version]
- Niogret, J.; Kendra, P.E.; Epsky, N.D.; Heath, R.R. Comparative analysis of terpenoid emissions from Florida host trees of the redbay ambrosia beetle, Xyleborus glabratus (Coleoptera: Curculionidae: Scolytinae). Fla. Entomol. 2011, 94, 1010–1017. [Google Scholar] [CrossRef]
- Kendra, P.E.; Montgomery, W.S.; Niogret, J.; Pruett, G.E.; Mayfield, A.E., III; MacKenzie, M.; Deyrup, M.A.; Bauchan, G.R.; Ploetz, R.C.; Epsky, N.D. North American Lauraceae: Terpenoid emissions, relative attraction and boring preferences of redbay ambrosia beetle, Xyleborus glabratus (Coleoptera: Curculionidae: Scolytinae). PLoS ONE 2014, 9, e102086. [Google Scholar] [CrossRef] [Green Version]
- Kendra, P.E.; Montgomery, W.S.; Deyrup, M.A.; Wakarchuk, D. Improved lure for redbay ambrosia beetle developed by enrichment of α-copaene content. J. Pest Sci. 2016, 89, 427–438. [Google Scholar] [CrossRef]
- Kendra, P.E.; Montgomery, W.S.; Schnell, E.Q.; Deyrup, M.A.; Epsky, N.D. Efficacy of α-copaene, cubeb, and eucalyptol lures for detection of redbay ambrosia beetle (Coleoptera: Curculionidae: Scolytinae). J. Econ. Entomol. 2016, 109, 2428–2435. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorea-Hernández, F.G. La familia Lauraceae en el sur de México: Diversidad, distribución y estado de conservación. Bot. Sci. 2002, 59–70. Available online: https://www.redalyc.org/articulo.oa?id=57707104 (accessed on 19 September 2022). [CrossRef] [Green Version]
- United States Department of Agriculture; Foreign Agricultural Service. Avocado Annual Report. 2020. Available online: https://apps.fas.usda.gov/newgainapi/api/Report/DownloadReportByFileName?fileName=Avocado%20Annual_Mexico%20City_Mexico_12-01-2020 (accessed on 19 September 2022).
- Kendra, P.E.; Montgomery, W.S.; Niogret, J.; Deyrup, M.A.; Guillén, L.; Epsky, N.D. Xyleborus glabratus, X. affinis, and X. ferrugineus (Coleoptera: Curculionidae: Scolytinae): Electroantennogram responses to host-based attractants and temporal patterns in host-seeking flight. Environ. Entomol. 2012, 41, 1597–1605. [Google Scholar] [CrossRef]
- Brar, G.S.; Capinera, J.L.; McLean, S.; Kendra, P.E.; Ploetz, R.C.; Peña, J.E. Effect of trap size, trap height and age of lure on sampling Xyleborus glabratus (Coleoptera: Curculionidae: Scolytinae), and its flight periodicity and seasonality. Fla. Entomol. 2012, 95, 1003–1011. [Google Scholar] [CrossRef]
- Atkinson, T.H.; Carrillo, D.; Duncan, R.E.; Peña, J.E. Occurrence of Xyleborus bispinatus (Coleoptera: Curculionidae: Scolytinae) Eichhoff in southern Florida. Zootaxa 2013, 3669, 96–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, S.M.; Gomez, D.F.; Beaver, R.A.; Hulcr, J.; Cognato, A.I. Reassessment of the species in the Euwallacea fornicatus (Coleoptera: Curculionidae: Scolytinae) complex after the rediscovery of the “lost” type specimen. Insects 2019, 10, 261. [Google Scholar] [CrossRef] [Green Version]
- Campbell, S.A.; Borden, J.H. Additive and synergistic integration of multimodal cues of both hosts and non-hosts during host selection by woodboring insects. Oikos 2009, 118, 553–563. [Google Scholar] [CrossRef]
- Ranger, C.M.; Reding, M.E.; Addesso, K.; Ginzel, M.; Rassati, D. Semiochemical-mediated host selection by Xylosandrus spp. ambrosia beetles (Coleoptera: Curculionidae) attacking horticultural tree crops: A review of basic and applied science. Can. Entomol. 2021, 153, 103–120. [Google Scholar] [CrossRef]
- Ranger, C.M.; Reding, M.E.; Persad, A.B.; Herms, D.A. Ability of stress-related volatiles to attract and induce attacks by Xylosandrus germanus and other ambrosia beetles. Agric. For. Entomol. 2010, 12, 177–185. [Google Scholar] [CrossRef]
- Ranger, C.M.; Tobin, P.C.; Reding, M.E. Ubiquitous volatile compound facilitates efficient host location by a non-native ambrosia beetle. Biol. Invasions 2015, 17, 675–686. [Google Scholar] [CrossRef]
- Eisner, T.; Grant, R.P. Toxicity, odor aversion, and “olfactory aposematism”. Science 1981, 213, 476. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strong, D.R.; Lawton, J.H.; Southwood, S.R. Insects on plants. In Community Patterns and Mechanisms; Blackwell Scientific Publicatons: Tallahasee, FL, USA, 1984. [Google Scholar]
- Kendra, P.E.; Niogret, J.; Montgomery, W.S.; Sanchez, J.S.; Deyrup, M.A.; Pruett, G.E.; Ploetz, R.C.; Epsky, N.D.; Heath, R.R. Temporal analysis of sesquiterpene emissions from manuka and phoebe oil lures and efficacy for attraction of Xyleborus glabratus (Coleoptera: Curculionidae: Scolytinae). J. Econ. Entomol. 2012, 105, 659–669. [Google Scholar] [CrossRef] [PubMed]
- Kendra, P.E.; Sanchez, J.S.; Montgomery, W.S.; Okins, K.E.; Niogret, J.; Peña, J.E.; Epsky, N.D.; Heath, R.R. Diversity of Scolytinae (Coleoptera: Curculionidae) attracted to avocado, lychee, and essential oil lures. Fla. Entomol. 2011, 94, 123–130. [Google Scholar] [CrossRef]
- Zhang, Q.H.; Schlyter, F. Olfactory recognition and behavioural avoidance of angiosperm nonhost volatiles by conifer-inhabiting bark beetles. Agric. For. Entomol. 2004, 6, 1–20. [Google Scholar] [CrossRef]
- Wardhaugh, C.W. The spatial and temporal distributions of arthropods in forest canopies: Uniting disparate patterns with hypotheses for specialisation. Biol. Rev. 2014, 89, 1021–1041. [Google Scholar] [CrossRef] [PubMed]
- Okins, K.E.; Thomas, M.C. New North American record for Xyleborinus andrewesi (Coleoptera: Curculionidae: Scolytinae). Fla. Entomol. 2010, 93, 133–134. [Google Scholar] [CrossRef]
- Carrillo, D.; Cruz, L.F.; Kendra, P.E.; Narvaez, T.I.; Montgomery, W.S.; Monterroso, A.; De Grave, C.; Cooperband, M.F. Distribution, pest status and fungal associates of Euwallacea nr. fornicatus in Florida avocado groves. Insects 2016, 7, 55. [Google Scholar] [CrossRef]
- Johnson, A.J.; Kendra, P.E.; Skelton, J.; Hulcr, J. Species diversity, phenology, and temporal flight patterns of Hypothenemus pygmy borers (Coleoptera: Curculionidae: Scolytinae) in South Florida. Environ. Entomol. 2016, 45, 627–632. [Google Scholar] [CrossRef] [Green Version]
- Kendra, P.E.; Niogret, J.; Montgomery, W.S.; Deyrup, M.A.; Epsky, N.D. Cubeb oil lures: Terpenoid emissions, trapping efficacy, and longevity for attraction of redbay ambrosia beetle (Coleoptera: Curculionidae: Scolytinae). J. Econ. Entomol. 2015, 108, 350–361. [Google Scholar] [CrossRef] [Green Version]
- Smith, R.H. Xylem Monoterpenes of Pines: Distribution, Variation, Genetics, Function; US Department of Agriculture, Forest Service, Pacific Southwest Research Station: Berkeley, CA, USA, 2000; Volume 177.
- Miller, D.R.; Rabaglia, R.J. Ethanol and (−)-α-pinene: Attractant kairomones for bark and ambrosia beetles in the southeastern US. J. Chem. Ecol. 2009, 35, 435–448. [Google Scholar] [CrossRef]
- Flint, M.; Liu, D.; Lee, J.; Beiriger, R.; Penrose, R.; Bright, D.; Seybold, S. Responses of red haired pine bark beetle (RPBB), Hylurgus ligniperda, and associated subcortical Coleoptera to host volatiles in southern California. In Proceedings of the Poster at the 2007 Entomological Society of America Annual Meeting, San Diego, CA, USA, 9–12 December 2007. [Google Scholar]
- Reding, M.E.; Schultz, P.B.; Ranger, C.M.; Oliver, J.B. Optimizing ethanol-baited traps for monitoring damaging ambrosia beetles (Coleoptera: Curculionidae, Scolytinae) in ornamental nurseries. J. Econ. Entomol. 2011, 104, 2017–2024. [Google Scholar] [CrossRef] [PubMed]
- Oliver, J.B.; Mannion, C.M. Ambrosia beetle (Coleoptera: Scolytidae) species attacking chestnut and captured in ethanol-baited traps in middle Tennessee. Environ. Entomol. 2001, 30, 909–918. [Google Scholar] [CrossRef]
- Rivera, M.J.; Martini, X.; Conover, D.; Mafra-Neto, A.; Carrillo, D.; Stelinski, L.L. Evaluation of semiochemical based push-pull strategy for population suppression of ambrosia beetle vectors of laurel wilt disease in avocado. Sci. Rep. 2020, 10, 2670. [Google Scholar] [CrossRef] [PubMed]
- Viloria, Z.; Villanueva, R.T.; Bessin, R.; O’Neal, P.; Ranger, C.M.; Dunwell, W. Scolytinae in nursery and fruit crops of Western Kentucky and seasonal population patterns of four invasive ambrosia beetles. J. Entomol. Sci. 2021, 56, 374–386. [Google Scholar] [CrossRef]
- Browne, F.G. The biology of Malayan Scolytidae and Platypodidae. Malay. For. Rec. 1961, 22, xi + 255. [Google Scholar]
- Cobb, R.C.; Filipe, J.A.; Meentemeyer, R.K.; Gilligan, C.A.; Rizzo, D.M. Ecosystem transformation by emerging infectious disease: Loss of large tanoak from California forests. J. Ecol. 2012, 100, 712–722. [Google Scholar] [CrossRef]
- Cruz, L.F.; Menocal, O.; Kendra, P.E.; Carrillo, D. Phoretic and internal transport of Raffaelea lauricola by different species of ambrosia beetle associated with avocado trees. Symbiosis 2021, 84, 151–161. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Species | Swampbay | Avocado |
|---|---|---|
| Subfamily Scolytinae | ||
| Tribe Xyleborini | ||
| Ambrosiodmus devexulus (Wood) | 21 | 51 |
| Ambrosiodmus lecontei Hopkins * | 77 | 54 |
| Euwallacea perbrevis (Schedl) | 0 | 402 |
| Premnobius cavipennis Eichhoff | 1 | 40 |
| Theoborus ricini (Eggers) | 0 | 19 |
| Xyleborinus andrewesi (Blandford) * | 1139 | 2 |
| Xyleborinus gracilis (Eichhoff) * | 16 | 42 |
| Xyleborinus saxesenii (Ratzeburg) * | 474 | 46,537 |
| Xyleborus affinis Eichhoff * | 23 | 653 |
| Xyleborus bispinatus Eichhoff * | 86 | 64 |
| Xyleborus ferrugineus (Fabricius) * | 2 | 2 |
| Xyleborus glabratus Eichhoff * | 254 | 5 |
| Xyleborus volvulus (Fabricius) * | 10 | 566 |
| Xylosandrus compactus (Eichhoff) | 36 | 11 |
| Xylosandrus crassiusculus (Motschulsky) * | 21 | 3425 |
| Tribe Cryphalini | ||
| Cryptocarenus heveae (Hagedorn) | 1 | 0 |
| Hypothenemus spp. | 91 | 863 |
| Tribe Corthylini | ||
| Corthylus papulans Eichhoff | 44 | 4 |
| Monarthrum mali (Fitch) | 1 | 0 |
| Subfamily Platypodinae | ||
| Euplatypus parallelus (Fabricius) | 17 | 14 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cloonan, K.R.; Montgomery, W.S.; Narvaez, T.I.; Carrillo, D.; Kendra, P.E. Community of Bark and Ambrosia Beetles (Coleoptera: Curculionidae: Scolytinae and Platypodinae) in Agricultural and Forest Ecosystems with Laurel Wilt. Insects 2022, 13, 971. https://doi.org/10.3390/insects13110971
Cloonan KR, Montgomery WS, Narvaez TI, Carrillo D, Kendra PE. Community of Bark and Ambrosia Beetles (Coleoptera: Curculionidae: Scolytinae and Platypodinae) in Agricultural and Forest Ecosystems with Laurel Wilt. Insects. 2022; 13(11):971. https://doi.org/10.3390/insects13110971
Chicago/Turabian StyleCloonan, Kevin R., Wayne S. Montgomery, Teresa I. Narvaez, Daniel Carrillo, and Paul E. Kendra. 2022. "Community of Bark and Ambrosia Beetles (Coleoptera: Curculionidae: Scolytinae and Platypodinae) in Agricultural and Forest Ecosystems with Laurel Wilt" Insects 13, no. 11: 971. https://doi.org/10.3390/insects13110971