
Reference Genes for Expression Analysis Using RT-qPCR in Cnaphalocrocis medinalis (Lepidoptera: Pyralidae)

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Rice Plant Preparing and Insect Rearing
2.2. Experimental Treatments
2.2.1. Developmental Stages
2.2.2. Larval Tissues
2.2.3. Larvae Feeding on Different Rice Varieties
2.2.4. Larvae Temperature Treatments
2.2.5. Adult Ages
2.2.6. Adult Nutritional Conditions
2.2.7. Adult Mating Statuses
2.2.8. Different Adult Take-Off Characteristics
2.3. Total RNA Isolation and cDNA Synthesis
2.4. Selection of Candidate Reference Genes and Primer Design
2.5. RT-qPCR
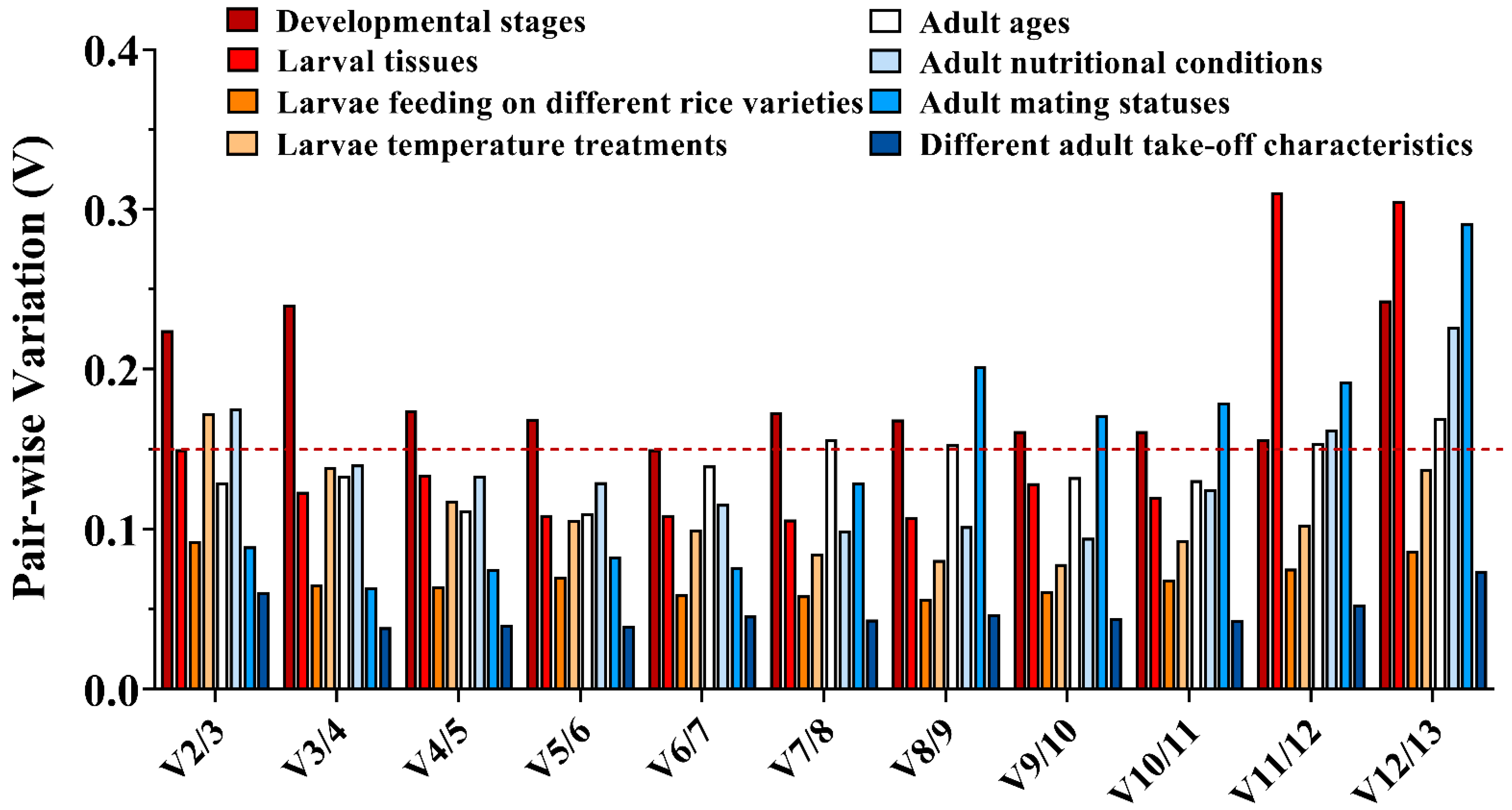
2.6. Expression Stability of Candidate Reference Genes under Different Treatments
2.7. Verification of Reference Gene
3. Results
3.1. Total RNA Quality and Amplification Efficiencies
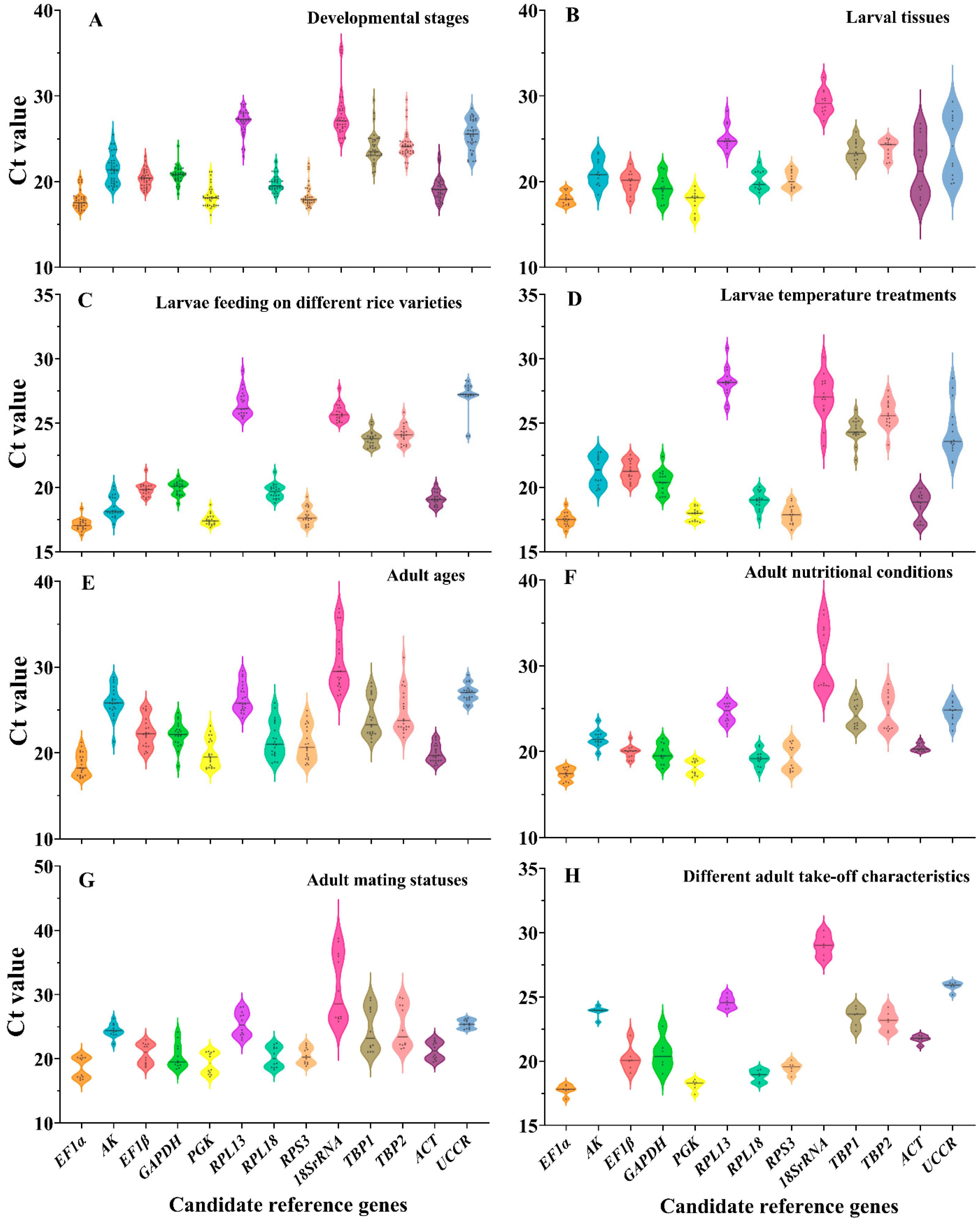
3.2. Expression Profiles of Candidate Reference Genes
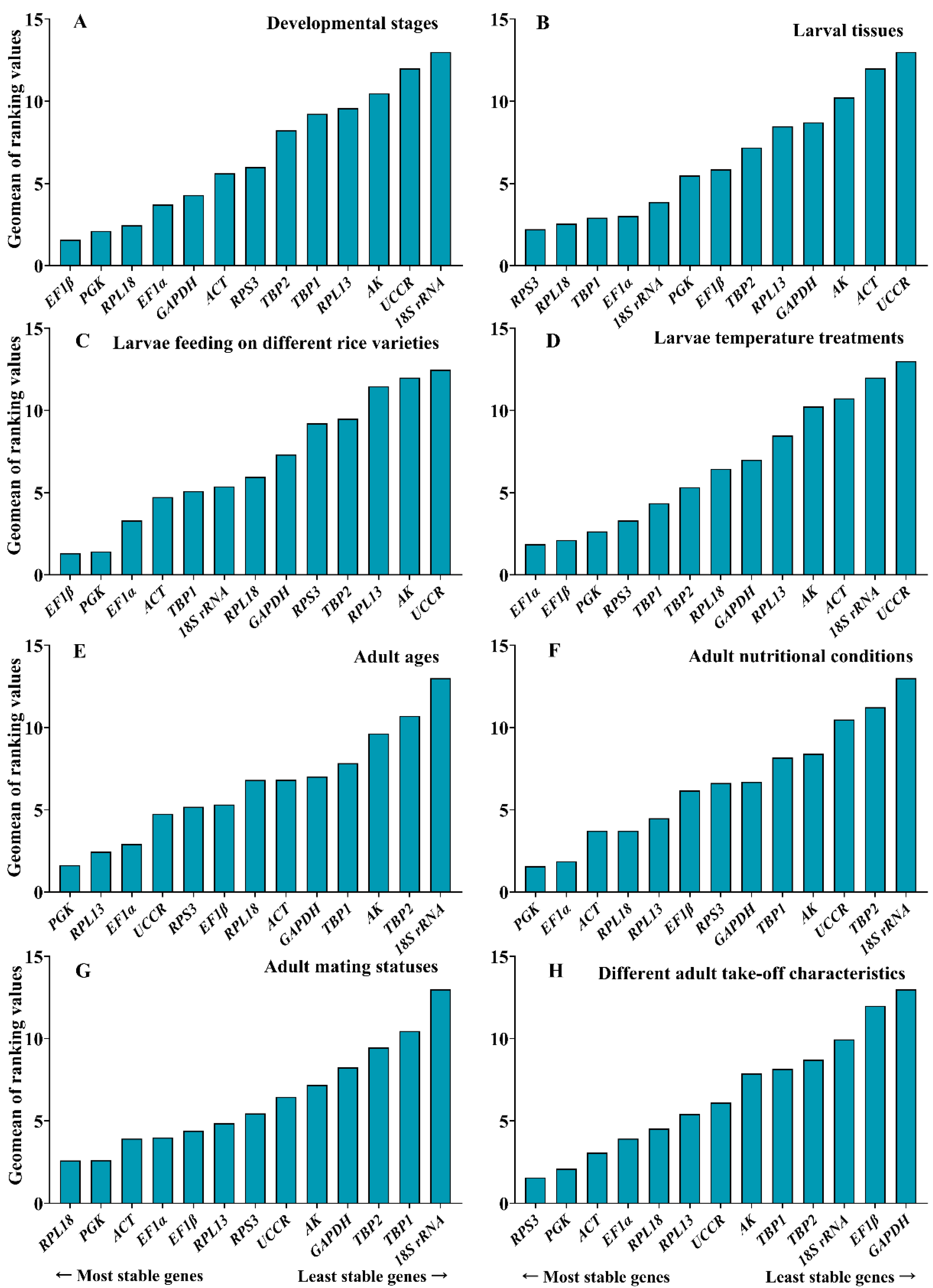
3.3. Stability of Candidate Reference Genes in C. medinalis under Different Experimental Conditions
3.3.1. Developmental Stages
3.3.2. Larval Tissues
3.3.3. Larvae Feeding on Different Rice Varieties
3.3.4. Larvae Temperature Treatments
3.3.5. Adult Ages
3.3.6. Adult Nutritional Conditions
3.3.7. Adult Mating Statuses
3.3.8. Different Adult Take-Off Characteristics
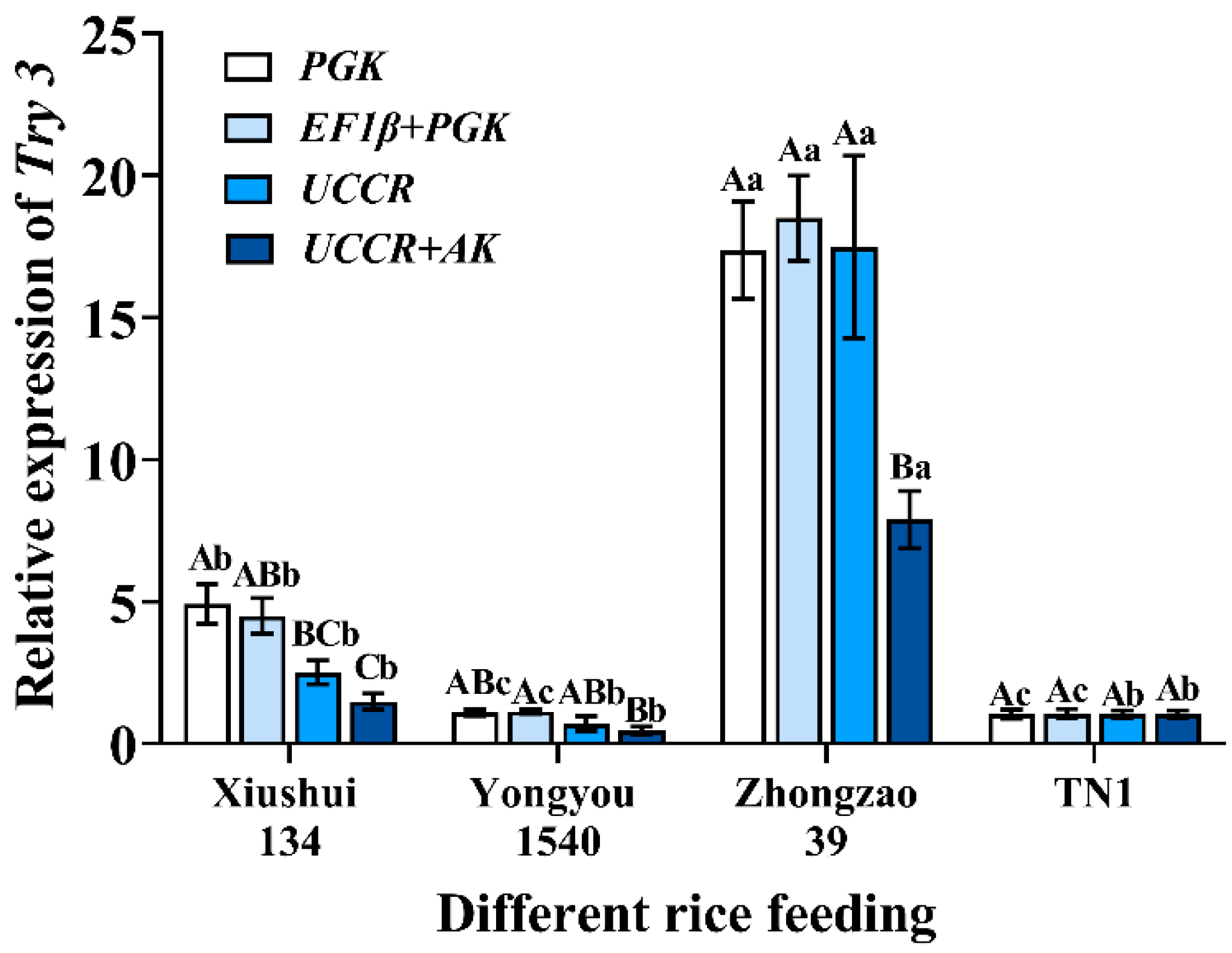
3.4. Validation of Reference Genes with Try3
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Zong, J.; Chen, J.; Li, L.; Li, J.; Li, D.; Wang, J.; Liu, J.; Liu, J. Reference gene selection for quantitative RT-PCR in Miscanthus sacchariflorus under abiotic stress conditions. Mol. Biol. Rep. 2022, 49, 907–915. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Li, Z.; Cao, J.; Zhang, S.; Zhang, H.; Wu, X.; Zhang, Q.; Liu, X. Selection and assessment of reference genes for quantitative PCR normalization in migratory locust Locusta migratoria (Orthoptera: Acrididae). PLoS ONE 2014, 9, e98164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, J.H.; Moon, K.; Kim, Y.; Kim, Y.H. Reference gene selection for qRT-PCR analysis of season- and tissue-specific gene expression profiles in the honey bee Apis mellifera. Sci. Rep. 2020, 10, 13935. [Google Scholar] [CrossRef] [PubMed]
- Chen, M.D.; Wang, B.; Li, Y.P.; Zeng, M.J.; Liu, J.T.; Ye, X.R.; Zhu, H.S.; Wen, Q.F. Reference gene selection for qRT-PCR analyses of luffa (Luffa cylindrica) plants under abiotic stress conditions. Sci. Rep. 2021, 11, 3161. [Google Scholar] [CrossRef]
- Wei, Z.H.; Liu, M.; Hu, C.; Yang, X.Q. Overexpression of glutathione s-transferase genes in field lambda-cyhalothrin-resistant population of Cydia pomonella: Reference gene selection and expression analysis. J. Agric. Food Chem. 2020, 68, 5825–5834. [Google Scholar] [CrossRef]
- Zhao, X.; Geng, Y.; Hu, T.; Zhao, Y.; Yang, S.; Hao, D. Evaluation of optimal reference genes for qRT-PCR analysis in Hyphantria cunea (Drury). Insects 2022, 13, 97. [Google Scholar] [CrossRef]
- Rahman, S.; Zhao, Z.; Umair, S.M.; Zhang, Y.; Jiang, H. Case study using recommended reference genes actin and 18S for reverse-transcription quantitative real-time PCR analysis in Myzus persicae. PLoS ONE 2021, 16, e0258201. [Google Scholar] [CrossRef]
- Sellamuthu, G.; Amin, S.; Bily, J.; Synek, J.; Modlinger, R.; Sen, M.K.; Chakraborty, A.; Roy, A. Reference gene selection for normalizing gene expression in Ips Sexdentatus (Coleoptera: Curculionidae: Scolytinae) under different experimental conditions. Front. Physiol. 2021, 12, 752768. [Google Scholar] [CrossRef]
- Shen, C.H.; Peng, L.J.; Zhang, Y.X.; Zeng, H.R.; Yu, H.F.; Jin, L.; Li, G.Q. Reference genes for expression analyses by qRT-PCR in Phthorimaea operculella (Lepidoptera: Gelechiidae). Insects 2022, 13, 140. [Google Scholar] [CrossRef]
- Deng, Y.C.; Zhao, H.X.; Yang, S.; Zhang, L.; Zhang, L.N.; Hou, C.S. Screening and validation of reference genes for RT-qPCR under different honey bee Viral Infections and dsRNA treatment. Front. Microbiol. 2020, 11, 1715. [Google Scholar] [CrossRef]
- Zhang, B.Z.; Liu, J.J.; Chen, X.L.; Yuan, G.H. Selection and evaluation of reference genes for gene expression using quantitative real-time PCR in Mythimna separata walker (Lepidoptera: Noctuidae). Entomol. Res. 2018, 48, 390–399. [Google Scholar] [CrossRef]
- Nakamura, A.M.; Chahad-Ehlers, S.; Lima, A.L.; Taniguti, C.H.; Sobrinho, I.J.; Torres, F.R.; de Brito, R.A. Reference genes for accessing differential expression among developmental stages and analysis of differential expression of OBP genes in Anastrepha obliqua. Sci. Rep. 2016, 6, 17480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shakeel, M.; Rodriguez, A.; Tahir, U.B.; Jin, F. Gene expression studies of reference genes for quantitative real-time PCR: An overview in insects. Biotechnol. Lett. 2018, 40, 227–236. [Google Scholar] [CrossRef] [PubMed]
- Zheng, X.S.; Ren, X.B.; Su, J.Y. Insecticide susceptibility of Cnaphalocrocis medinalis (Lepidoptera: Pyralidae) in China. J. Econ. Entomol. 2011, 104, 653–658. [Google Scholar] [CrossRef] [PubMed]
- Deng, Q.Q.; Ye, M.; Wu, X.B.; Song, J.; Wang, J.; Chen, L.N.; Zhu, Z.Y.; Xie, J. Damage of brown planthopper (BPH) Nilaparvata lugens and rice leaf folder (RLF) Cnaphalocrocis medinalis in parent plants lead to distinct resistance in ratoon rice. Plant Signal. Behav. 2022, 17, 2096790. [Google Scholar] [CrossRef]
- Lv, W.X.; Jiang, X.F.; Chen, X.J.; Cheng, Y.X.; Xia, J.X.; Zhang, L. Flight and reproduction variations of rice leaf roller, Cnaphalocrocis medinalis in response to different rearing temperatures. Insects 2021, 12, 1083. [Google Scholar] [CrossRef]
- Guo, J.W.; Cui, Y.D.; Lin, P.J.; Zhai, B.P.; Lu, Z.X.; Chapman, J.W.; Hu, G. Male nutritional status does not impact the reproductive potential of female Cnaphalocrocis medinalis moths under conditions of nutrient shortage. Insect Sci. 2021, 29, 467–477. [Google Scholar] [CrossRef]
- Guo, J.W.; Yang, F.; Li, P.; Liu, X.D.; Wu, Q.L.; Hu, G.; Zhai, B.P. Female bias in an immigratory population of Cnaphalocrocis medinalis moths based on field surveys and laboratory tests. Sci. Rep. 2019, 9, 18388. [Google Scholar] [CrossRef] [Green Version]
- Liang, G.C.; Ouyang, Y.C.; Dai, S.M. Detection and classification of rice infestation with rice leaf folder (Cnaphalocrocis medinalis) using hyperspectral imaging techniques. Remote Sens. 2021, 13, 4587. [Google Scholar] [CrossRef]
- Alka, K.; Rabindra, P. Impact of abiotic factors on incidence of rice leaf folder (Cnaphalocrocis medinalis Guenée) in agro-climatic condition of Ranchi, Jharkhand. J. Eco-Friendly Agric. 2022, 17, 100–104. [Google Scholar]
- Vennila, S.; Adlul, I.; Nisar, S.; Bhat, M.N.; Ankur, T.; Sanjay, S.; Preetinder, S.S.; Mathirajan, V.G.; Rao, M.S.; Prabhakar, M. Prediction of rice leaf folder Cnaphalocrosis medinalis (Guenée) for future periods under climate change scenario of 4.5 representative concentration pathway. Plant Prot. Sci. 2019, 27, 214. [Google Scholar] [CrossRef]
- Liu, S.; Rao, X.J.; Li, M.Y.; Feng, M.F.; He, M.Z.; Li, S.G. Glutathione s-transferase genes in the leaffolder, Cnaphalocrocis medinalis (Lepidoptera; Pyralidae): Indentication and expression profiles. Arch. Insect Biochem. Physiol. 2015, 90, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Han, G.J.; Liu, Q.; Li, C.M.; Xu, B.; Xu, J. Transcriptome sequencing reveals Cnaphalocrocis medinalis against baculovirus infection by oxidative stress. Mol. Immunol. 2021, 129, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Wen, N.; Chen, J.J.; Chen, G.; Du, L.X.; Chen, H.; Li, Y.H.; Peng, Y.F.; Yang, X.W.; Han, L.Z. The overexpression of insect endogenous microRNA in transgenic rice inhibits the pupation of Chilo suppressalis and Cnaphalocrocis medinalis. Pest Manag. Sci. 2021, 77, 3990–3999. [Google Scholar] [CrossRef]
- Shakeel, M.; Du, J.; Li, S.W.; Zhou, Y.J.; Sarwar, N.; Bukhari, S.A.H. Characterization, knockdown and parental effect of hexokinase gene of Cnaphalocrocis medinalis (Lepidoptera: Pyralidae) revealed by RNA interference. Genes 2020, 11, 1258. [Google Scholar] [CrossRef]
- Chen, J.; Li, C.; Yang, Z.F. Identification and expression of two novel cytochrome P450 genes, CYP6CV1 and CYP9A38, in Cnaphalocrocis medinalis (Lepidoptera: Pyralidae). J. Insect Sci. 2015, 15, 50. [Google Scholar] [CrossRef] [Green Version]
- Tian, P.; Qiu, L.; Zhou, A.; Chen, G.; He, H.; Ding, W.; Li, Y. Evaluation of appropriate reference genes for investigating gene expression in Chlorops oryzae (Diptera: Chloropidae). J. Econ. Entomol. 2019, 112, 2207–2214. [Google Scholar] [CrossRef]
- Tan, Y.; Zhou, X.R.; Pang, B.P. Reference gene selection and evaluation for expression analysis using qRT-PCR in Galeruca daurica (Joannis). Bull. Entomol. Res. 2016, 107, 359–368. [Google Scholar] [CrossRef]
- Liu, Z.Y.; Han, H.; Chen, W.; Wang, S.W.; Meng, F.M.; Cai, J.F.; Guo, Y.D. Evaluation of reference genes and age estimation of forensically useful Aldrichina grahami (Diptera: Calliphoridae) during intrapuparial period. J. Med. Entomol. 2021, 58, 47–55. [Google Scholar] [CrossRef]
- Chen, C.; Li, S.Y.; Zhu, H.; Fan, B.Q.; Wang, Y.; Hao, D.J. Identification and evaluation of reference genes for gene expression analysis in the weevil pest Pagiophloeus tsushimanus using RT-qPCR. J. Asia-Pacif. Entomol. 2020, 23, 336–344. [Google Scholar] [CrossRef]
- Li, X.N.; Gong, P.P.; Wang, B.T.; Wang, C.; Li, M.Y.; Zhang, Y.H.; Li, X.R.; Gao, H.F.; Ju, J.S.; Zhu, X. Selection and validation of experimental condition-specific reference genes for qRT-PCR in Metopolophium dirhodum (Walker) (Hemiptera: Aphididae). Sci. Rep. 2020, 10, 21951. [Google Scholar] [CrossRef] [PubMed]
- Lu, J.; Yang, C.X.; Zhang, Y.J.; Pan, H.P. Selection of reference genes for the normalization of RT-qPCR data in gene expression studies in insects: A systematic review. Front. Physiol. 2018, 9, 1560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.J.; Xu, H.X.; Wu, Z.H.; LU, Z.X. Effects of inhibitors on the protease profiles and degradation of activated Cry toxins in larval midgut juices of Cnaphalocrocis medinalis (Lepidoptera: Pyralidae). J. Integr. Agric. 2021, 20, 2195–2203. [Google Scholar] [CrossRef]
- Zhu, A.X.; Qiu, Q.; Liu, X.D. A method for rearing the rice leaf folder(Cnaphalocrocis medinalis) using wheat seedlings. J. Appl. Entomol. 2015, 52, 883–889. [Google Scholar]
- Tu, K.Y.; Tsai, S.F.; Guo, T.W.; Lin, H.H.; Yang, Z.W.; Liao, C.T.; Chuang, W.P. The role of plant abiotic factors on the interactions between Cnaphalocrocis medinalis (Lepidoptera: Crambidae) and its host plant. Environ. Entomol. 2018, 47, 857–866. [Google Scholar] [CrossRef]
- Huang, J.C.; Zhang, G.A.; Wang, B.P. Effects of age, ambient temperature and reproductive status on wing beat frequency of the rice leaf roller Cnaphalocrocis medinalis (Guenée) (Lepidoptera: Crambidae). Appl. Entomol. Zool. 2013, 48, 499–505. [Google Scholar] [CrossRef]
- Wang, F.Y.; Yang, F.; Lu, M.H.; Luo, S.Y.; Zhai, B.P.; Lim, K.S.; McInerney, C.E.; Hu, G. Determining the migration duration of rice leaf folder (Cnaphalocrocis medinalis (Guenée)) moths using a trajectory analytical approach. Sci. Rep. 2017, 7, 39853. [Google Scholar] [CrossRef]
- Green, M.R.; Sambrook, J. Removing DNA contamination from RNA samples by treatment with RNase-free DNase I. Cold Spring Harb. Protoc. 2019, 10, 709–710. [Google Scholar] [CrossRef]
- Zhao, X.X.; Xu, H.X.; He, K.; Shi, Z.M.; Chen, X.; Ye, X.H.; Mei, Y.; Yang, Y.J.; Li, M.Z.; Gao, L.B.; et al. A chromosome-level genome assembly of rice leaffolder, Cnaphalocrocis medinalis. Mol. Ecol. Resour. 2021, 21, 561–572. [Google Scholar] [CrossRef]
- Derveaux, S.; Vandesompele, J.; Hellemans, J. How to do successful gene expression analysis using real-time PCR. Nat. Methods 2010, 50, 227–230. [Google Scholar] [CrossRef]
- Wang, Z.; Meng, Q.Q.; Zhu, X.; Sun, S.W.; Liu, A.Q.; Gao, S.F.; Gou, Y.F. Identification and evaluation of reference genes for normalization of gene expression in developmental stages, sexes, and tissues of Diaphania caesalis (Lepidoptera, Pyralidae). J. Insect Sci. 2020, 20, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Liang, C.; Han, S.P.; Han, H.; Zhao, F.; He, Y.Z. Selection of reference genes for Harmonia axyridis (Coleoptera: Coccinellidae) feeding on different diets. J. Asia-Pacif. Entomol. 2019, 22, 1115–1122. [Google Scholar] [CrossRef]
- Qu, C.; Wang, R.; Che, W.N.; Zhu, X.; Li, F.Q.; Luo, C. Selection and evaluation of reference genes for expression analysis using quantitative real-time PCR in the Asian Ladybird Harmonia axyridis (Coleoptera: Coccinellidae). PLoS ONE 2018, 13, e0192521. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vandesompele, J.; Katleen, D.P.; Filip, P.; Bruce, P.; Nadine, V.R.; Anne, D.P.; Frank, S. Accurate normalization of real-time quantitative RT-PCR data by geometric averaging of multiple internal control genes. Genome Biol. 2002, 3, research0034.1–research0034.11. [Google Scholar] [CrossRef] [Green Version]
- Wang, X.; Xiong, M.; Wang, J.; Lei, C.; Zhu, F. Reference gene stability of a synanthropic fly, Chrysomya megacephala. Parasit Vectors 2015, 8, 565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.H.; Zheng, X.S.; Liang, Q.; Xu, H.X.; Yang, Y.J.; Tian, J.C.; He, X.C.; Lu, Z.X. Evaluation and validation of reference genes for SYBR Green qRT-PCR normalization in Sesamia inferens (Lepidoptera: Noctuidae). J. Asia-Pacif. Entomol. 2015, 18, 669–675. [Google Scholar] [CrossRef]
- Sellamuthu, G.; Bily, J.; Joga, M.R.; Synek, J.; Roy, A. Identifying optimal reference genes for gene expression studies in Eurasian spruce bark beetle, Ips typographus (Coleoptera: Curculionidae: Scolytinae). Sci. Rep. 2022, 12, 4671. [Google Scholar] [CrossRef]
- Toutges, M.J.; Hartzer, K.; Lord, J.; Oppert, B. Evaluation of reference genes for quantitative polymerase chain reaction across life cycle stages and tissue types of Tribolium castaneum. J. Agric. Food Chem. 2010, 58, 8948–8951. [Google Scholar] [CrossRef]
- Hansen, M.C.; Nielsen, A.K.; Molin, S.; Hammer, K.; Kilstrup, M. Changes in rRNA levels during stress invalidates results from mRNA blotting: Fluorescence in situ rRNA hybridization permits renormalization for estimation of cellular mRNA levels. J. Bacteriol. 2001, 183, 4747–4751. [Google Scholar] [CrossRef] [Green Version]
- Guo, J.W.; Li, P.; Zhang, J.; Liu, X.D.; Zhai, B.P.; Hu, G. Cnaphalocrocis medinalis moths decide to migrate when suffering nutrient shortage on the first day after emergence. Insects 2019, 10, 364. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Colell, A.; Green, D.R.; Ricci, J.E. Novel roles for GAPDH in cell death and carcinogenesis. Cell Death Differ. 2009, 16, 1573–1581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, K.S.; Li, F.; Liang, P.Z.; Chen, X.W.; Liu, Y.; Gao, X.W. Identification and validation of reference genes for the normalization of gene expression data in qRT-PCR analysis in Aphis gossypii (Hemiptera: Aphididae). J. Insect Sci. 2016, 16, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Yu, Y.C.; Zhang, J.; Huang, C.; Hou, X.J.; Sun, X.L.; Xiao, B. Reference genes selection for quantitative gene expression studies in tea green leafhoppers, Empoasca onukii Matsuda. PLoS ONE 2018, 13, e0205182. [Google Scholar] [CrossRef] [PubMed]
- Pinheiro, D.H.; Siegfried, B.D. Selection of reference genes for normalization of RT-qPCR data in gene expression studies in Anthonomus eugenii Cano (Coleoptera: Curculionidae). Sci. Rep. 2020, 10, 5070. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Labroussaa, F.; Dubrana, M.P.; Nathalie, A.B.; Béven, L.; Saillard, C. Involvement of a minimal actin-binding region of Spiroplasma citri phosphoglycerate kinase in spiroplasma transmission by its leafhopper vector. PLoS ONE 2011, 6, e17357. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehfuss, L.R.; Feng, Y.; Lissemore, J.L.; Sullivan, D.T. Structure and expression of the phosphoglycerate kinase (Pgk) gene of Drosophila melanogaster. Mol. Gen. Genet. 1992, 235, 213–220. [Google Scholar] [CrossRef]
- Dzaki, N.; Azzam, G. Assessment of Aedes albopictus reference genes for quantitative PCR at different stages of development. PLoS ONE 2018, 13, e0194664. [Google Scholar] [CrossRef] [Green Version]
- Lazarević, J.; Tomanić, M.J. Dietary and phylogenetic correlates of digestive trypsin activity in insect pests. Entomol. Exp. Appl. 2015, 157, 123–151. [Google Scholar] [CrossRef]
- Fu, W.; Xie, W.; Zhang, Z.; Wang, S.L.; Wu, Q.J.; Liu, Y.; Zhou, X.M.; Zhou, X.G.; Zhang, Y.J. Exploring valid reference genes for quantitative real-time PCR analysis in Plutella xylostella (Lepidoptera: Plutellidae). Int. J. Biol. Sci. 2013, 9, 792–802. [Google Scholar] [CrossRef] [Green Version]
- Quan, P.Q.; Li, M.Z.; Wang, G.R.; Gu, L.L.; Liu, X.D. Comparative transcriptome analysis of the rice leaf folder (Cnaphalocrocis medinalis) to heat acclimation. BMC Genom. 2020, 21, 450. [Google Scholar] [CrossRef]
- Yang, Y.J.; Xu, H.X.; Lu, Y.H.; Wang, C.Y.; Lu, Z.X. Midgut transcriptomal response of the rice leaffolder, Cnaphalocrocis medinalis (Guenée) to Cry1C toxin. PLoS ONE 2018, 13, e0191686. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, P.L.; Guo, Z.Q.; Liu, X.D. Cuticular protein genes involve heat acclimation of insect larvae under global warming. Insect Mol. Biol. 2022, 31, 519–532. [Google Scholar] [CrossRef] [PubMed]
- Schmittgen, T.D.; Livak, K.J. Analyzing real-time PCR data by the comparative C(T) method. Nat. Protoc. 2008, 3, 1101–1108. [Google Scholar] [CrossRef] [PubMed]
- Yu, H.Z.; Wen, D.F.; Wang, W.L.; Geng, L.; Zhang, Y.; Xu, J.P. Identification of genes putatively involved in chitin metabolism and insecticide detoxification in the rice leaf folder (Cnaphalocrocis medinalis) larvae through transcriptomic analysis. Int. J. Mol. Sci. 2015, 16, 21873–21896. [Google Scholar] [CrossRef] [PubMed]
- Shakeel, M.; Du, J.; Li, S.W.; Zhou, Y.J.; Sarwar, N.; Guo, X. Glucosamine-6-phosphate N-acetyltransferase gene silencing by parental RNA interference in rice leaf folder, Cnaphalocrocis medinalis (Lepidoptera: Pyralidae). Sci. Rep. 2022, 12, 2141. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene Name | Gene Symbol | Primer Sequence (5′ to 3′) | Product Size (bp) | Tm (°C) | Efficiency (%) | Regression Coefficient (R2) | Slope |
|---|---|---|---|---|---|---|---|
| Elongation factor 1 α | EF1α | F: CTGCTGTCGCTTTCGTCCC R: CTTGCCCTCAGCCTTACCCTC | 122 | 55 | 105 | 0.992 | −3.217 |
| Arginine kinase | AK | F: CGCAACCCTCGAGAAATTGGA R: ACACCCGACTGGATGCAA | 159 | 55 | 112 | 0.996 | −3.071 |
| Elongation factor 1 β | EF1β | F: CTTCTTACACTCCCGCCGAAC R: GCGTCCTCTTCCTCATCACC | 154 | 55 | 108 | 0.996 | −3.135 |
| Glyceraldehyde-3-Phosphate dehydrogenase | GAPDH | F: CTGCCACTCAAAAGACCGT R: AAGGCCATACCAGTCAGT | 154 | 53 | 104 | 0.992 | −3.233 |
| Phosphoglycerate kinase | PGK | F: CAGCCCTCATTGCAAAGTCC R: GCAGCTTGTTGATTCCATAACCA | 162 | 57 | 109 | 0.999 | −3.115 |
| Ribosomal protein L 13 | RPL13 | F: ATCAACAGCCGTCAGATCG R: TTTCCATTGTGTGTCGCCTC | 193 | 55 | 109 | 0.995 | −3.117 |
| Ribosomal protein L 18 | RPL18 | F: GGCGCACCGAAGTTAAATCTCA R: AGCCACGGTCATCTTAGGAAC | 263 | 54 | 110 | 0.996 | −3.111 |
| Ribosomal protein S 3 | RPS3 | F: AGGTTCAACATCCCCGAGCA R: CGGACACAACAACCTCGCAAC | 193 | 55 | 109 | 0.995 | −3.114 |
| 18S ribosomal RNA | 18S rRNA | F: TTTTATAATGCCGACGAAGCGAGA R: CCCGAAAGCCCTGAACCAC | 155 | 56 | 104 | 0.990 | −3.226 |
| TATA-box binding protein 1 | TBP1 | F: AATGCTGAATACAACCCGAAG R: TCCTAGCAGCTAATCTTGAGT | 142 | 55 | 108 | 0.982 | −3.141 |
| TATA-box binding protein 2 | TBP2 | F: ATAACCAATGCTGCAAACACC R: CGCTGTCTTTCATTTGTAGAACCA | 128 | 55 | 108 | 0.996 | −3.146 |
| β-actin | ACT | F: CACACAGTGCCCATCTACGA R: GCGGTGGTGGTGAATGAGTA | 125 | 55 | 102 | 0.998 | −3.276 |
| Ubiquinol-cytochrome c reductase | UCCR | F: ACAGTCGCCTTCAAAGCTGGT R: CCAATCTGTGCCAACTTGCGT | 165 | 55 | 119 | 0.999 | −2.937 |
| Experimental Conditions | Ranking | ΔCt | BestKeeper | NormFinder | geNorm | ||||
|---|---|---|---|---|---|---|---|---|---|
| Developmental stages | 1 | EF1β | 1.264 | GAPDH | 0.660 | RPL18 | 0.537 | EF1β PGK | 0.632 |
| 2 | PGK | 1.278 | EF1β | 0.798 | PGK | 0.570 | — | — | |
| 3 | RPL18 | 1.286 | RPL18 | 0.800 | EF1β | 0.587 | EF1α | 0.838 | |
| 4 | EF1α | 1.332 | EF1α | 0.840 | EF1α | 0.682 | RPL18 | 0.745 | |
| 5 | ACT | 1.375 | PGK | 0.875 | ACT | 0.733 | ACT | 0.922 | |
| 6 | RPS3 | 1.395 | RPS3 | 0.911 | RPS3 | 0.813 | RPS3 | 0.955 | |
| 7 | GAPDH | 1.403 | RPL13 | 0.997 | GAPDH | 0.858 | GAPDH | 0.996 | |
| 8 | TBP2 | 1.470 | ACT | 1.005 | TBP2 | 0.906 | TBP2 | 1.053 | |
| 9 | TBP1 | 1.594 | TBP2 | 1.023 | TBP1 | 1.127 | TBP1 | 1.114 | |
| 10 | AK | 1.780 | TBP1 | 1.302 | AK | 1.397 | RPL13 | 1.204 | |
| 11 | RPL13 | 1.813 | AK | 1.398 | RPL13 | 1.469 | AK | 1.289 | |
| 12 | UCCR | 2.304 | UCCR | 1.426 | UCCR | 2.065 | UCCR | 1.440 | |
| 13 | 18S rRNA | 2.449 | 18S rRNA | 2.027 | 18S rRNA | 2.339 | 18S rRNA | 1.595 | |
| Larval tissues | 1 | RPL18 | 1.270 | EF1α | 0.624 | RPS3 | 0.153 | 18S rRNA TBP1 | 0.371 |
| 2 | RPS3 | 1.283 | TBP2 | 0.894 | RPL18 | 0.153 | — | — | |
| 3 | TBP1 | 1.318 | RPS3 | 0.896 | EF1α | 0.219 | RPL18 | 0.446 | |
| 4 | EF1α | 1.367 | EF1β | 0.990 | TBP1 | 0.326 | RPS3 | 0.490 | |
| 5 | 18S rRNA | 1.414 | PGK | 1.006 | 18S rRNA | 0.585 | PGK | 0.586 | |
| 6 | PGK | 1.457 | TBP1 | 1.027 | PGK | 0.633 | EF1β | 0.640 | |
| 7 | EF1β | 1.508 | RPL18 | 1.029 | EF1β | 0.738 | EF1α | 0.697 | |
| 8 | GAPDH | 1.557 | RPL13 | 1.081 | GAPDH | 0.773 | RPL13 | 0.761 | |
| 9 | RPL13 | 1.626 | 18S rRNA | 1.113 | RPL13 | 0.933 | GAPDH | 0.832 | |
| 10 | AK | 1.802 | GAPDH | 1.125 | AK | 1.115 | AK | 0.941 | |
| 11 | TBP2 | 1.837 | AK | 1.261 | TBP2 | 1.121 | TBP2 | 1.035 | |
| 12 | ACT | 4.052 | ACT | 3.144 | ACT | 3.944 | ACT | 1.494 | |
| 13 | UCCR | 4.054 | UCCR | 3.450 | UCCR | 3.949 | UCCR | 1.888 | |
| Larvae feeding on different rice varieties | 1 | EF1β | 0.529 | PGK | 0.322 | EF1β | 0.177 | EF1β PGK | 0.247 |
| 2 | PGK | 0.563 | EF1α | 0.328 | PGK | 0.249 | — | — | |
| 3 | EF1α | 0.597 | EF1β | 0.366 | 18S rRNA | 0.293 | ACT | 0.284 | |
| 4 | TBP1 | 0.597 | ACT | 0.420 | TBP1 | 0.299 | EF1α | 0.296 | |
| 5 | 18S rRNA | 0.604 | GAPDH | 0.435 | EF1α | 0.341 | RPL18 | 0.324 | |
| 6 | ACT | 0.612 | RPL18 | 0.458 | RPL18 | 0.355 | TBP1 | 0.371 | |
| 7 | RPL18 | 0.619 | TBP1 | 0.466 | ACT | 0.373 | 18S rRNA | 0.399 | |
| 8 | GAPDH | 0.691 | 18S rRNA | 0.513 | GAPDH | 0.441 | RPS3 | 0.433 | |
| 9 | TBP2 | 0.755 | RPS3 | 0.532 | TBP2 | 0.553 | GAPDH | 0.466 | |
| 10 | RPS3 | 0.757 | TBP2 | 0.533 | RPS3 | 0.609 | TBP2 | 0.509 | |
| 11 | RPL13 | 0.868 | UCCR | 0.638 | RPL13 | 0.699 | RPL13 | 0.566 | |
| 12 | AK | 1.007 | AK | 0.735 | AK | 0.870 | AK | 0.635 | |
| 13 | UCCR | 1.209 | RPL13 | 0.807 | UCCR | 1.114 | UCCR | 0.724 | |
| Larvae temperature treatments | 1 | EF1β | 0.895 | EF1α | 0.387 | EF1β | 0.412 | EF1α PGK | 0.507 |
| 2 | RPS3 | 0.921 | PGK | 0.426 | TBP1 | 0.464 | — | — | |
| 3 | EF1α | 0.924 | RPL18 | 0.515 | TBP2 | 0.489 | RPS3 | 0.555 | |
| 4 | PGK | 0.933 | RPS3 | 0.565 | EF1α | 0.506 | EF1β | 0.597 | |
| 5 | TBP1 | 0.937 | EF1β | 0.648 | RPS3 | 0.520 | TBP2 | 0.659 | |
| 6 | TBP2 | 0.942 | TBP1 | 0.687 | PGK | 0.574 | TBP1 | 0.684 | |
| 7 | GAPDH | 0.998 | GAPDH | 0.696 | GAPDH | 0.632 | GAPDH | 0.713 | |
| 8 | RPL18 | 1.040 | RPL13 | 0.740 | RPL13 | 0.715 | RPL18 | 0.735 | |
| 9 | RPL13 | 1.056 | TBP2 | 0.835 | RPL18 | 0.743 | RPL13 | 0.770 | |
| 10 | AK | 1.146 | ACT | 0.863 | AK | 0.885 | AK | 0.805 | |
| 11 | ACT | 1.211 | AK | 1.023 | ACT | 0.896 | ACT | 0.869 | |
| 12 | 18S rRNA | 1.402 | 18S rRNA | 1.291 | 18S rRNA | 1.168 | 18S rRNA | 0.953 | |
| 13 | UCCR | 1.925 | UCCR | 1.602 | UCCR | 1.803 | UCCR | 1.102 | |
| Adult ages | 1 | PGK | 1.041 | UCCR | 0.807 | PGK | 0.202 | PGK RPL13 | 0.404 |
| 2 | RPL13 | 1.110 | ACT | 0.892 | EF1α | 0.281 | — | — | |
| 3 | EF1α | 1.113 | GAPDH | 1.022 | RPL13 | 0.302 | EF1α | 0.426 | |
| 4 | RPS3 | 1.161 | EF1α | 1.158 | RPS3 | 0.488 | EF1β | 0.505 | |
| 5 | EF1β | 1.197 | AK | 1.231 | EF1β | 0.538 | RPS3 | 0.556 | |
| 6 | RPL18 | 1.231 | RPL13 | 1.374 | RPL18 | 0.602 | RPL18 | 0.615 | |
| 7 | TBP1 | 1.392 | PGK | 1.396 | TBP1 | 0.902 | TBP1 | 0.739 | |
| 8 | UCCR | 1.546 | EF1β | 1.401 | UCCR | 1.136 | UCCR | 0.886 | |
| 9 | GAPDH | 1.668 | RPS3 | 1.709 | GAPDH | 1.334 | ACT | 1.020 | |
| 10 | TBP2 | 1.692 | RPL18 | 1.744 | TBP2 | 1.365 | GAPDH | 1.117 | |
| 11 | ACT | 1.712 | TBP1 | 1.960 | ACT | 1.380 | TBP2 | 1.203 | |
| 12 | AK | 2.134 | TBP2 | 2.075 | AK | 1.960 | AK | 1.335 | |
| 13 | 18S rRNA | 2.314 | 18S rRNA | 2.919 | 18S rRNA | 2.184 | 18S rRNA | 1.485 | |
| Adult nutritional conditions | 1 | PGK | 1.024 | ACT | 0.379 | PGK | 0.169 | EF1α PGK | 0.338 |
| 2 | EF1α | 1.061 | EF1β | 0.520 | EF1α | 0.310 | — | — | |
| 3 | RPL13 | 1.140 | EF1α | 0.594 | RPL13 | 0.380 | RPL18 | 0.481 | |
| 4 | RPL18 | 1.152 | RPL18 | 0.710 | RPL18 | 0.512 | ACT | 0.554 | |
| 5 | RPS3 | 1.205 | AK | 0.718 | RPS3 | 0.512 | RPL13 | 0.628 | |
| 6 | ACT | 1.214 | PGK | 0.808 | GAPDH | 0.570 | GAPDH | 0.709 | |
| 7 | GAPDH | 1.221 | UCCR | 0.896 | TBP1 | 0.627 | RPS3 | 0.768 | |
| 8 | TBP1 | 1.256 | GAPDH | 0.911 | ACT | 0.721 | TBP1 | 0.807 | |
| 9 | EF1β | 1.351 | RPL13 | 1.000 | EF1β | 0.952 | EF1β | 0.857 | |
| 10 | AK | 1.411 | TBP1 | 1.377 | AK | 1.013 | AK | 0.904 | |
| 11 | TBP2 | 1.609 | RPS3 | 1.430 | TBP2 | 1.232 | TBP2 | 1.004 | |
| 12 | UCCR | 2.236 | TBP2 | 1.957 | UCCR | 2.145 | UCCR | 1.173 | |
| 13 | 18S rRNA | 2.982 | 18S rRNA | 3.395 | 18S rRNA | 2.933 | 18S rRNA | 1.451 | |
| Adult mating statuses | 1 | RPL18 | 1.230 | UCCR | 0.560 | RPL18 | 0.088 | EF1α PGK | 0.210 |
| 2 | PGK | 1.248 | AK | 0.707 | RPL13 | 0.249 | — | — | |
| 3 | ACT | 1.271 | RPS3 | 1.105 | PGK | 0.268 | EF1β | 0.261 | |
| 4 | RPL13 | 1.286 | ACT | 1.260 | ACT | 0.412 | RPL18 | 0.277 | |
| 5 | EF1β | 1.295 | EF1β | 1.465 | EF1β | 0.478 | ACT | 0.331 | |
| 6 | EF1α | 1.316 | RPL18 | 1.488 | EF1α | 0.518 | RPS3 | 0.398 | |
| 7 | RPS3 | 1.341 | EF1α | 1.555 | RPS3 | 0.576 | RPL13 | 0.454 | |
| 8 | GAPDH | 1.558 | PGK | 1.573 | GAPDH | 0.668 | GAPDH | 0.608 | |
| 9 | TBP2 | 2.027 | GAPDH | 1.583 | TBP2 | 1.540 | TBP2 | 0.887 | |
| 10 | TBP1 | 2.102 | RPL13 | 1.717 | TBP1 | 1.660 | TBP1 | 1.065 | |
| 11 | AK | 2.234 | TBP2 | 3.040 | AK | 1.974 | AK | 1.224 | |
| 12 | UCCR | 2.669 | TBP1 | 3.116 | UCCR | 2.564 | UCCR | 1.433 | |
| 13 | 18S rRNA | 3.814 | 18S rRNA | 4.845 | 18S rRNA | 3.770 | 18S rRNA | 1.799 | |
| Different adult take-off characteristics | 1 | RPS3 | 0.374 | ACT | 0.208 | RPS3 | 0.085 | PGK RPS3 | 0.093 |
| 2 | PGK | 0.391 | UCCR | 0.240 | PGK | 0.186 | — | — | |
| 3 | RPL18 | 0.410 | EF1α | 0.258 | RPL13 | 0.203 | ACT | 0.156 | |
| 4 | EF1α | 0.414 | AK | 0.276 | RPL18 | 0.222 | EF1α | 0.165 | |
| 5 | ACT | 0.419 | PGK | 0.310 | EF1α | 0.237 | RPL18 | 0.188 | |
| 6 | RPL13 | 0.423 | RPS3 | 0.340 | ACT | 0.264 | RPL13 | 0.212 | |
| 7 | TBP1 | 0.463 | RPL18 | 0.364 | TBP1 | 0.278 | UCCR | 0.247 | |
| 8 | TBP2 | 0.478 | RPL13 | 0.392 | TBP2 | 0.312 | AK | 0.277 | |
| 9 | 18S rRNA | 0.524 | TBP2 | 0.581 | 18S rRNA | 0.381 | TBP1 | 0.316 | |
| 10 | UCCR | 0.535 | TBP1 | 0.592 | UCCR | 0.459 | TBP2 | 0.347 | |
| 11 | AK | 0.569 | 18S rRNA | 0.661 | AK | 0.483 | 18S rRNA | 0.375 | |
| 12 | EF1β | 0.672 | EF1β | 0.668 | EF1β | 0.566 | EF1β | 0.424 | |
| 13 | GAPDH | 1.007 | GAPDH | 0.973 | GAPDH | 0.957 | GAPDH | 0.514 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhao, X.; Guo, J.; Lu, Y.; Sun, T.; Tian, J.; Huang, J.; Xu, H.; Wang, Z.; Lu, Z. Reference Genes for Expression Analysis Using RT-qPCR in Cnaphalocrocis medinalis (Lepidoptera: Pyralidae). Insects 2022, 13, 1046. https://doi.org/10.3390/insects13111046
Zhao X, Guo J, Lu Y, Sun T, Tian J, Huang J, Xu H, Wang Z, Lu Z. Reference Genes for Expression Analysis Using RT-qPCR in Cnaphalocrocis medinalis (Lepidoptera: Pyralidae). Insects. 2022; 13(11):1046. https://doi.org/10.3390/insects13111046
Chicago/Turabian StyleZhao, Xiaoyu, Jiawen Guo, Yanhui Lu, Tianyi Sun, Junce Tian, Jianlei Huang, Hongxing Xu, Zhengliang Wang, and Zhongxian Lu. 2022. "Reference Genes for Expression Analysis Using RT-qPCR in Cnaphalocrocis medinalis (Lepidoptera: Pyralidae)" Insects 13, no. 11: 1046. https://doi.org/10.3390/insects13111046