
Identification and Expression Analysis of Odorant-Binding and Chemosensory Protein Genes in Virus Vector Nephotettix cincticeps

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects Rearing and Tissue Collection
2.2. cDNA Library Construction and Transcriptome Sequencing
2.3. De Novo Assembly, Unigene Annotation, and Functional Classification
2.4. Identification of OBP and CSP Genes
2.5. Sequence Alignment and Phylogenetic Analysis
2.6. Tissue Expression Profile Analysis
2.7. Statistical Analysis
3. Results
3.1. Overview of the Head and Rest of the Body Transcriptomes
3.2. Homology Searches and Functional Annotation
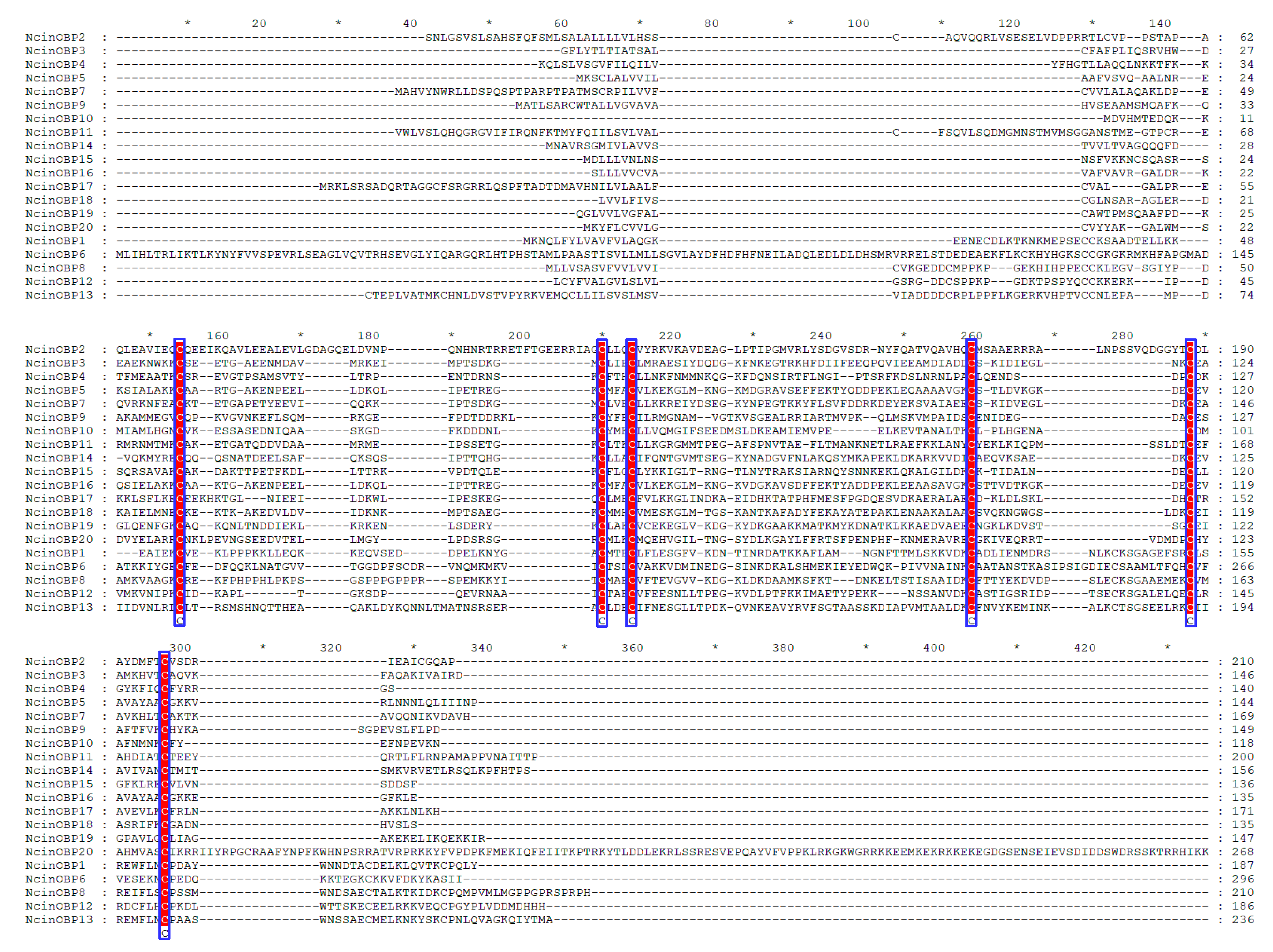
3.3. Identification of Candidate OBP and CSP Genes in N. cincticeps
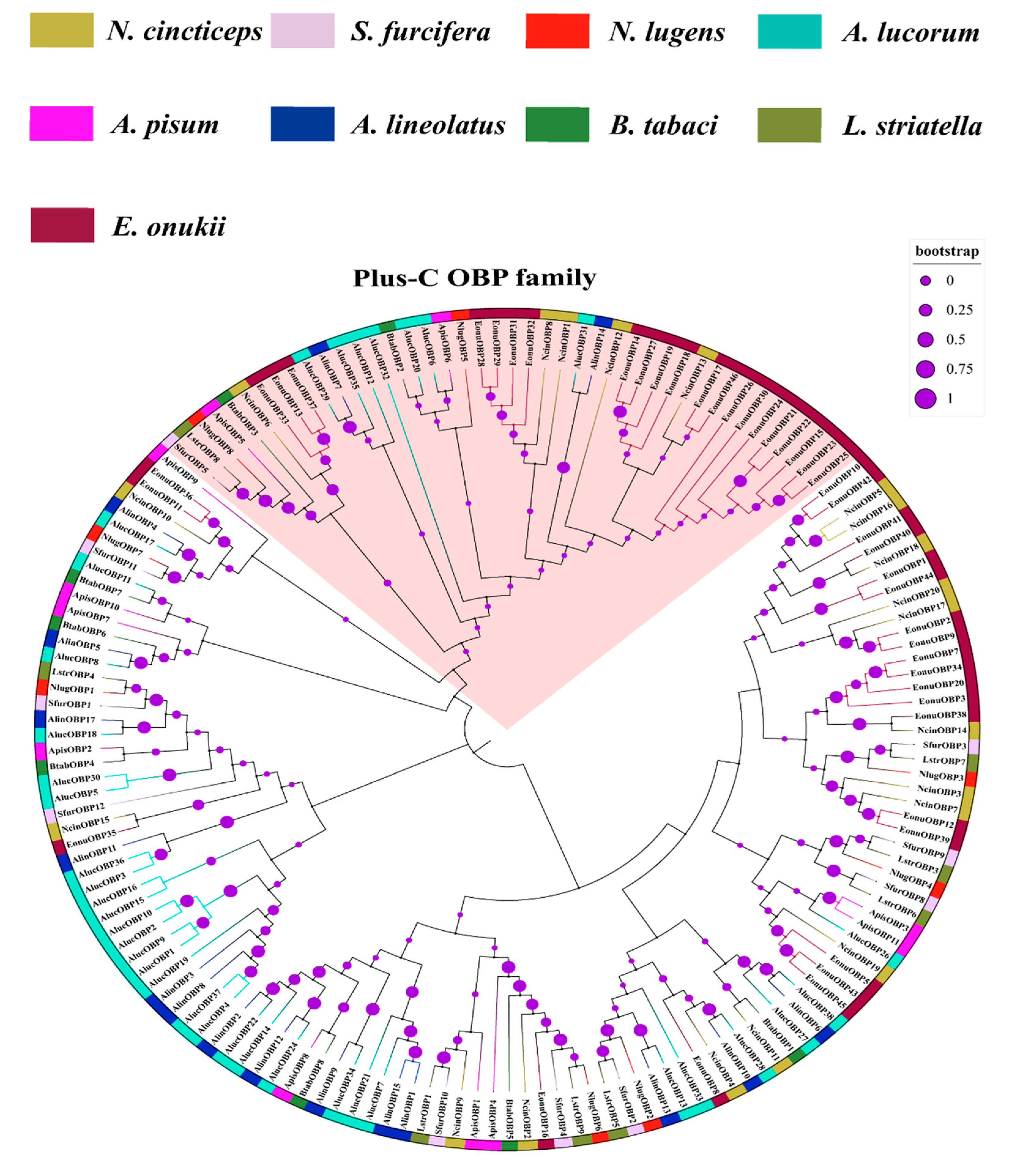
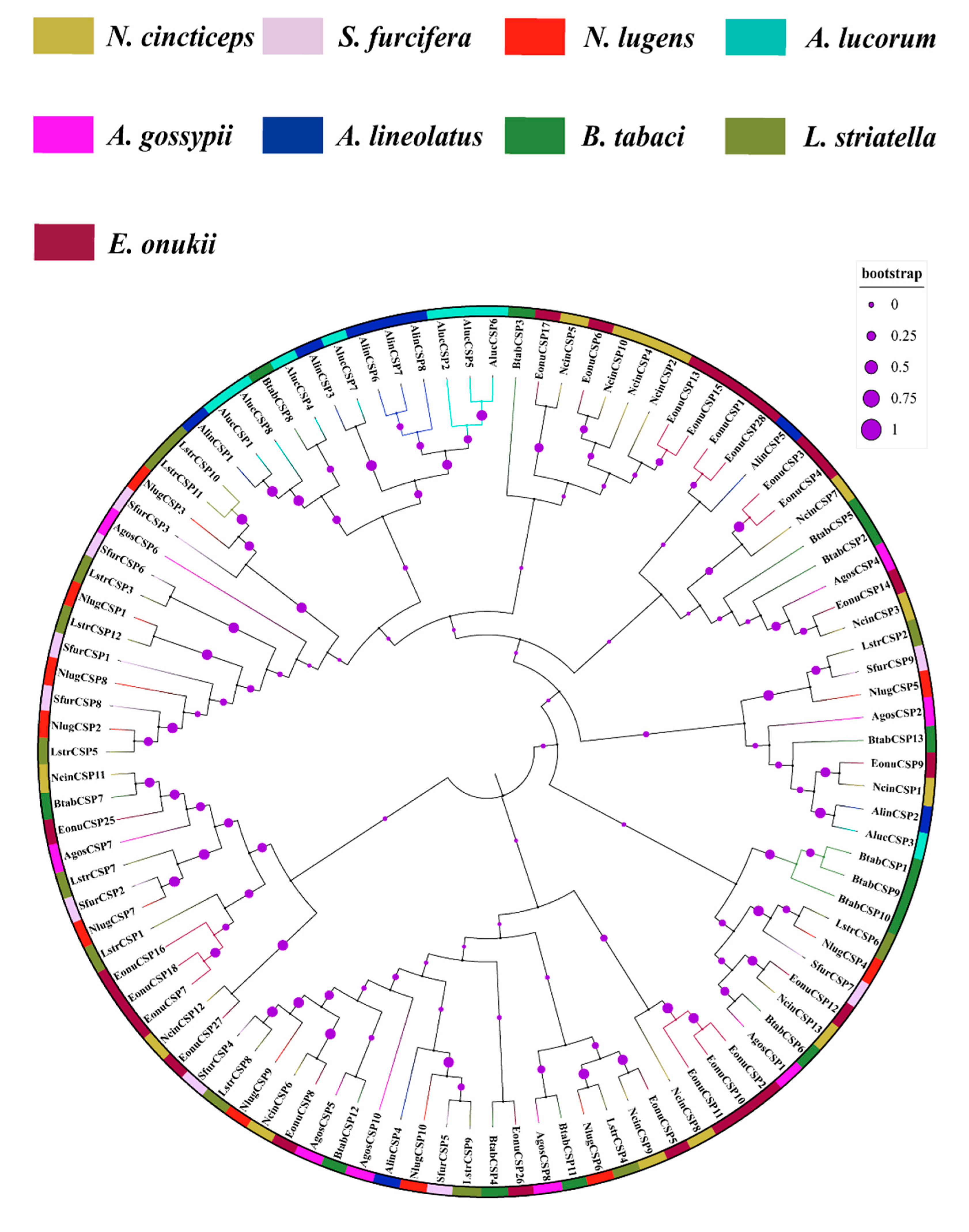
3.4. Phylogenetic Analysis of N. cincticeps OBP and CSP Genes
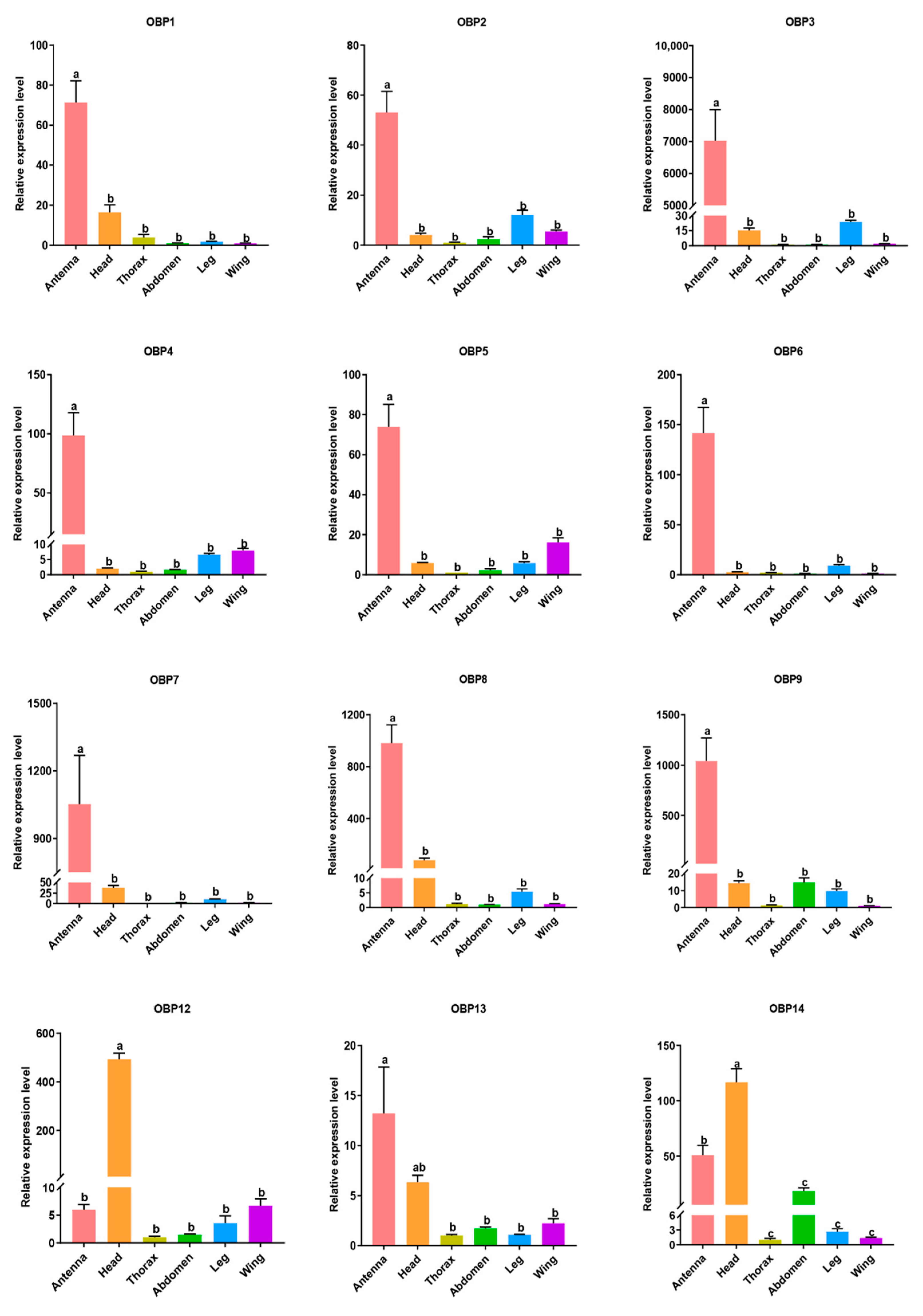
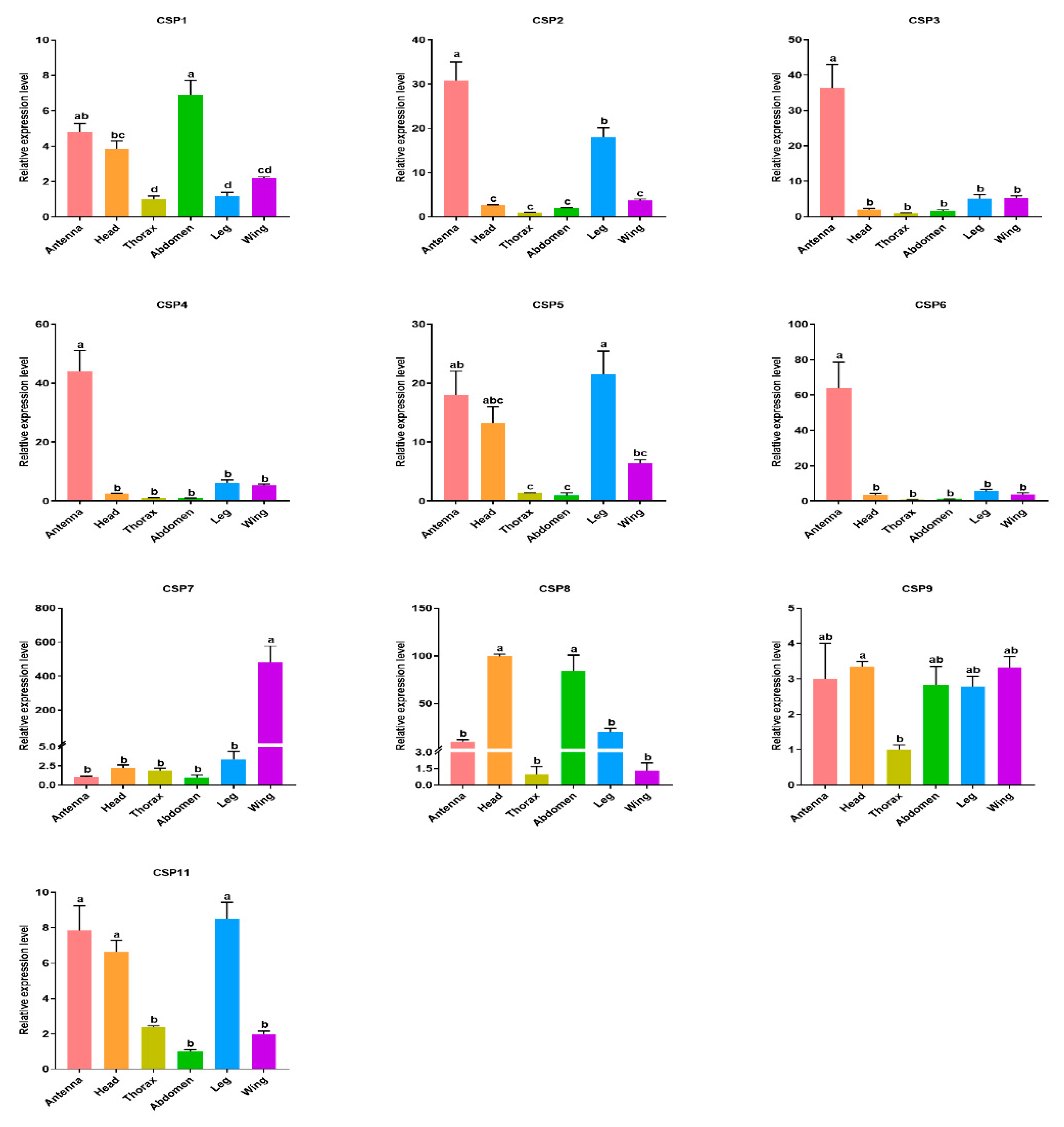
3.5. Tissue Expression Levels of Candidate OBP and CSP Genes
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gadenne, C.; Barrozo, R.B.; Anton, S. Plasticity in insect olfaction: To smell or not to Smell? Annu. Rev. Entomol. 2016, 61, 317–333. [Google Scholar] [CrossRef]
- Wang, Z.H.; Yang, F.; Sun, A.; Song, J.Y.; Shan, S.; Zhang, Y.J.; Wang, S. Expressional and functional comparisons of five clustered odorant binding proteins in the brown marmorated stink bug Halyomirpha halys. Int. J. Biol. Macromol. 2022, 209, 1352–1358. [Google Scholar] [CrossRef]
- Leal, W. Odorant reception in insects: Roles of receptors, binding proteins, and degrading enzymes. Annu. Rev. Entomol. 2013, 58, 373–391. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Iovinella, I.; Zhu, J.; Wang, G.R.; Dani, F.R. Beyond chemoreception: Diverse tasks of soluble olfactory proteins in insects. Biol. Rev. 2018, 93, 184–200. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pelosi, P.; Zhou, J.J.; Ban, L.P.; Calvello, M. Soluble proteins in insect chemical communication. Cell. Mol. Life Sci. 2006, 63, 1658–1676. [Google Scholar] [CrossRef] [PubMed]
- Pelosi, P.; Iovinella, I.; Felicioli, A.; Dani, F.R. Soluble proteins of chemical communication: An overview across arthropods. Front. Physiol. 2014, 5, 320. [Google Scholar] [CrossRef] [Green Version]
- He, P.; Li, Z.Q.; Zhang, Y.F.; Chen, L.; Wang, J.; Xu, L.; He, M. Identification of odorant-binding and chemosensory protein genes and the ligand affinity of two of the encoded proteins suggest a complex olfactory perception system in Periplaneta americana. Insect Mol. Biol. 2017, 26, 687–701. [Google Scholar] [CrossRef]
- Rihani, K.; Ferveur, J.F.; Briand, L. The 40-Year mystery of insect odorant-binding proteins. Biomolecules 2021, 11, 509. [Google Scholar] [CrossRef]
- Laughlin, J.D.; Ha, T.S.; Jones, D.N.M.; Smith, D.P. Activation of pheromone-sensitive neurons is mediated by conformational activation of pheromone-binding protein. Cell 2008, 133, 1255–1265. [Google Scholar] [CrossRef] [Green Version]
- Grosse-Wilde, E.; Svatos, A.; Krieger, J. A pheromone-binding protein mediates the bombykol-induced activation of a pheromone receptor in vitro. Chem. Senses 2006, 31, 547–555. [Google Scholar] [CrossRef]
- Gomez-Diaz, C.; Reina, J.H.; Cambillau, C.; Benton, R. Ligands for pheromone-sensing neurons are not conformationally activated odorant binding proteins. PLoS Biol. 2013, 11, e1001546. [Google Scholar] [CrossRef] [Green Version]
- Benton, R.; Vannice, K.S.; Vosshall, L.B. An essential role for a CD36-related receptor in pheromone detection in Drosophila. Nature 2007, 450, 289–293. [Google Scholar] [CrossRef] [PubMed]
- Suh, E.; Bohbot, J.; Zwiebel, L.J. Peripheral olfactory signaling in insects. Curr. Opin. Insect Sci. 2014, 6, 86–92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brito, N.F.; Moreira, M.F.; Melo, A.C. A look inside odorant-binding proteins in insect chemoreception. J. Insect Physiol. 2016, 95, 51–56. [Google Scholar] [CrossRef] [PubMed]
- Sanchez-Gracia, A.; Vieira, F.; Rozas, J. Molecular evolution of the major chemosensory gene families in insects. Heredity 2009, 103, 208–216. [Google Scholar] [CrossRef] [PubMed]
- Andersson, M.N.; Grosse-Wilde, E.; Keeling, C.I.; Bengtsson, J.M.; Yuen, M.M.; Li, M.; Hillbur, Y.; Bohlmann, J.; Hansson, B.S.; Schlyter, F. Antennal transcriptome analysis of the chemosensory gene families in the tree killing bark beetles, Ips typographus and Dendroctonus ponderosae (Coleoptera: Curculionidae: Scolytinae). BMC Genom. 2013, 14, 198. [Google Scholar] [CrossRef] [Green Version]
- Zhou, J.J. Odorant-binding proteins in insects. Vitam. Horm. 2010, 83, 241–272. [Google Scholar] [CrossRef]
- He, P.; Zhang, J.; Liu, N.Y.; Zhang, Y.N.; Yang, K.; Dong, S.L. Distinct expression profiles and different functions of odorant binding proteins in Nilaparvata lugens Stål. PLoS ONE 2011, 6, e28921. [Google Scholar] [CrossRef] [Green Version]
- Shanbhag, S.R.; Park, S.K.; Pikielny, C.W.; Steinbrecht, R.A. Gustatory organs of Drosophila melanogaster: Fine structure and expression of the putative odorant-binding protein PBPRP2. Cell Tissue Res. 2001, 304, 423–437. [Google Scholar] [CrossRef]
- Jeong, Y.T.; Shim, J.; Oh, S.R.; Yoon, H.I.; Kim, C.H.; Moon, S.J.; Montell, C. An odorant-binding protein required for suppression of sweet taste by bitter chemicals. Neuron 2013, 79, 725–737. [Google Scholar] [CrossRef]
- Zhang, J.J.; Mao, K.K.; Ren, Z.J.; Jin, R.H.; Zhang, Y.H.; Cai, T.W.; He, S.; Li, J.; Wan, H. Odorant bingding protein 3 is associated with nitenpyram and sulfoxaflor resistance in Nilaparvata lugens. Int. J. Biol. Macromol. 2022, 209, 1352–1358. [Google Scholar] [CrossRef] [PubMed]
- Harada, E.; Haba, D.; Aigaki, T.; Matsuo, T. Behavioral analyses of mutants for two odorant-binding protein genes, Obp57d and Obp57e, in Drosophila melanogaster. Genes Genet. Syst. 2008, 83, 257–264. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swarup, S.; Morozova, T.V.; Sridhar, S.; Nokes, M.; Anholt, R.R. Modulation of feeding behavior by odorant-binding proteins in Drosophila melanogaster. Chem. Senses 2014, 39, 125–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rihani, K.; Fraichard, S.; Chauvel, I.; Poirier, N.; Delompré, T.; Neiers, F.; Tanimura, T.; Ferveur, J.-F.; Briand, L. A conserved odorant binding protein is required for essential amino acid detection in Drosophila. Commun. Biol. 2019, 2, 425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lartigue, A.; Campanacci, V.; Roussel, A.; Larsson, A.M.; Jones, T.A.; Tegoni, M.; Cambillau, C. X-ray structure and ligand binding study of a moth chemosensory protein. J. Biol. Chem. 2002, 277, 32094–32098. [Google Scholar] [CrossRef] [Green Version]
- Tomaselli, S.; Crescenzi, O.; Sanfelice, D.; Ab, E.; Wechselberger, R.; Angeli, S.; Scaloni, A.; Boelens, R.; Tancredi, T.; Pelosi, A.P.; et al. Solution structure of a chemosensory protein from the desert locust Schistocerca gregaria. Biochemistry 2006, 45, 10606–10613. [Google Scholar] [CrossRef]
- Jacquin-Joly, E.; Vogt, R.G.; Francois, M.C.; Meillour, P.N.-L. Functional and expression pattern analysis of chemosensory proteins expressed in antennae and pheromonal gland of Mamestra brassicae. Chem. Senses 2001, 26, 833–844. [Google Scholar] [CrossRef]
- Iovinella, I.; Bozza, F.; Caputo, B.; Torre, A.D.; Pelosi, P. Ligand-binding study of Anopheles gambiae chemosensory proteins. Chem. Senses 2013, 38, 409–419. [Google Scholar] [CrossRef] [Green Version]
- Kitabayashi, A.N.; Arai, T.; Kubo, T.; Natori, S. Molecular cloning of cDNA for p10, a novel protein that increases in the regenerating legs of Periplaneta Americana (American cockroach). Insect Biochem. Mol. Biol. 1998, 28, 785–790. [Google Scholar] [CrossRef]
- Levy, F.; Bulet, P.; Ehret-Sabatier, F. Proteomic analysis of the systemic immune response of Drosophila. Mol. Cell. Proteom. 2004, 3, 156–166. [Google Scholar] [CrossRef]
- Maleszka, J.; Foret, S.; Saint, R.; Maleszka, R. RNAi-induced phenotypes suggest a novel role for a chemosensory protein CSP5 in the development of embryonic integument in the honeybee (Apis mellifera). Dev. Genes Evol. 2007, 217, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; He, X.; Lehane, S.; Lehane, M.; Hertz-Fowler, C.; Berriman, M.; Field, L.M.; Zhou, J.-J. Expression of chemosensory proteins in the tsetse fly Glossina morsitans morsitans is related to female host-seeking behavior. Insect Mol. Biol. 2012, 21, 41–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.L.; Guo, H.; Huang, L.Q.; Pelosi, P.; Wang, C.Z. Unique function of a chemosensory protein in the proboscis of two Helicoverpa species. J. Exp. Biol. 2014, 217, 1821–1826. [Google Scholar] [CrossRef] [Green Version]
- Liu, G.; Ma, H.; Xie, H.; Xuan, N.; Guo, X.; Fan, Z.; Rajashekar, B.; Arnaud, P.; Offmann, B.; Picimbon, J.-F. Biotype characterization, developmental profiling, insecticide response and binding property of Bemisia tabaci chemosensory proteins: Role of CSP in insect defense. PLoS ONE 2016, 11, e0154706. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pathak, M.D.; Khan, Z.R. Insect Pests of Rice; International Rice Research Institute: Manila, Philippines, 1994. [Google Scholar]
- Abo, M.E.; Sy, A.A. Rice virus diseases. J. Sustain. Agric. 1998, 11, 113–134. [Google Scholar] [CrossRef]
- Wei, T.Y.; Li, Y. Rice reoviruses in insect vectors. Annu. Rev. Phytopathol. 2016, 54, 99–120. [Google Scholar] [CrossRef] [PubMed]
- Chang, X.F.; Wang, F.; Fang, Q.; Chen, F.; Yao, H.W.; Gatehouse, A.M.R.; Ye, G. Virus-induced plant volatiles mediate the olfactory behavior of its insect vectors. Plant Cell Environ. 2021, 44, 2700–2715. [Google Scholar] [CrossRef]
- Chang, X.F.; Sun, L.L.; Ning, D.; Dang, C.; Yao, H.W.; Fang, Q.; Peng, Y.; Wang, F.; Ye, G. Cry1C rice doesn’t affect the ecological fitness of rice brown planthopper, Nilaparvata lugens either under RDV stress or not. Sci. Rep. 2020, 10, 16423. [Google Scholar] [CrossRef]
- Martin, M. Cutadapt removes adapter sequences from high-throughput sequencing reads. EMBnet J. 2011, 17, 10–12. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.D.; et al. Full-length transcriptome assembly from RNA-seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644. [Google Scholar] [CrossRef]
- Buchfink, B.; Xie, C.; Huson, D.H. Fast and sensitive protein alignment using diamond. Nat. Methods 2015, 12, 59. [Google Scholar] [CrossRef] [PubMed]
- Larkin, M.A.; Blackshields, G.; Brown, N.P.; Chenna, R.; McGettigan, P.A.; McWilliam, H.; Valentin, F.; Wallace, I.M.; Wilm, A.; Lopez, R.; et al. Clustal W and Clustal X version 2.0. Bioinformatics 2007, 23, 2947–2948. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, Y.; Li, H.; Wang, Q.; Liu, T.J.; Zhang, L.X.; Wu, M.; Xu, Y.; Zhang, Z.; Gu, S. Identification and expression analysis of chemosensory genes in the tea green leafhopper, Empoasca onukii Matsuda. J. Appl. Entomol. 2018, 142, 828–846. [Google Scholar] [CrossRef]
- Tamura, K.; Stecher, G.; Peterson, D.; Filipski, A.; Kumar, S. MEGA6: Molecular evolutionary genetics analysis version 6.0. Mol. Biol. Evol. 2013, 30, 2725–2729. [Google Scholar] [CrossRef] [Green Version]
- Xu, G.; Chang, X.F.; Gu, G.X.; Jia, W.X.; Guo, L.; Huang, J.; Ye, G.-Y. Molecular and pharmacological characterization of a β-adrenergic-like octopamine receptor from the green rice leafhopper Nephotettix cincticeps. Insect Biochem. Mol. Biol. 2020, 120, 103337. [Google Scholar] [CrossRef]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCt Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Xu, Y.L.; He, P.; Zhang, L.; Fang, S.Q.; Dong, S.L.; Zhang, Y.J.; Li, F. Large-scale identification of odorant-binding proteins and chemosensory proteins from expressed sequence tags in insects. BMC Genom. 2009, 10, 632. [Google Scholar] [CrossRef] [Green Version]
- Zhao, S.; Hong, W.; Wu, J.; Wang, Y.; Ji, S.; Zhu, S.; Wei, C.; Zhang, J.; Li, Y. A viral protein promotes host SAMS1 activity and ethylene production for the benefit of virus infection. Elife 2017, 6, e27529. [Google Scholar] [CrossRef]
- Zhou, J.; Vieira, F.G.; He, X.; Smadja, C.; Liu, R.; Rozas, J.; Field, L.M. Genome annotation and comparative analyses of the odorant-binding proteins and chemosensory proteins in the pea aphid Acyrthosiphon pisum. Insect Mol. Biol 2010, 19, 113–122. [Google Scholar] [CrossRef] [Green Version]
- Wang, R.; Li, F.; Zhang, W.; Zhang, X.M.; Qu, C.; Tetreau, G.; Sun, L.; Luo, C. Identification and expression profile analysis of odorant binding protein and chemosensory protein genes in Bemisia tabaci MED by head transcriptome. PLoS ONE 2017, 12, e0171739. [Google Scholar] [CrossRef]
- Zhou, S.S.; Sun, Z.; Ma, W.; Chen, W.; Wang, M.Q. De novo analysis of the Nilaparvata lugens (Stål) antenna transcriptome and expression patterns of olfactory genes. Comp. Biochem. Physiol. Part D Genom. Proteom. 2014, 9, 31–39. [Google Scholar] [CrossRef] [PubMed]
- Cui, H.H.; Gu, S.H.; Zhu, X.Q.; Wei, Y.; Liu, H.W.; Khalid, H.D.; Guo, Y.-Y.; Zhang, Y.-J. Odorant-binding and chemosensory proteins identified in the antennal transcriptome of Adelphocoris suturalis Jakovlev. Comp. Biochem. Physiol. Part D Genom. Proteom. 2017, 24, 139–145. [Google Scholar] [CrossRef] [PubMed]
- Yuan, H.B.; Ding, Y.X.; Gu, S.H.; Sun, L.; Zhu, X.Q.; Liu, H.W.; Dhiloo, K.H.; Zhang, Y.-J.; Guo, Y.-Y. Molecular characterization and expression profiling of odorant-binding proteins in Apolygus lucorum. PLoS ONE 2015, 10, e0140562. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, W.W.; Yuan, X.; Qian, P.; Cheng, J.A.; Zhang, C.X.; Gurr, G.; Zhu, Z. Identification and expression profiling of putative chemosensory protein genes in two rice planthoppers, Laodelphax striatellus (Fallen) and Sogatella furcifera (Horvath). J. Asia-Pac. Entomol. 2015, 18, 771–778. [Google Scholar] [CrossRef]
- Cai, X.M.; Xu, X.X.; Bian, L.; Luo, Z.X.; Xin, Z.J.; Chen, Z.M. Attractiveness of host volatiles combined with background visual cues to the tea leafhopper, Empoasca vitis. Entomol. Exp. Appl. 2015, 157, 291–299. [Google Scholar] [CrossRef]
- Guo, Y.Y. Crop diseases and insect pests in China. In Tea Diseases and Insect Pests; Institute of Plant Protection, Chinese Academy of Agricultural Sciences & China Society of Plant Protection, Eds.; China Agriculture Press: Beijing, China, 2014; Volume 3, pp. 97–101. [Google Scholar]
- Bian, L.; Li, Z.Q.; Ma, L.; Cai, X.M.; Luo, Z.X.; Chen, Z.M. Identification of the genes in tea leafhopper, Empoasca onukii (Hemiptera: Cicadellidae), that encode odorant-binding proteins and chemosensory proteins using transcriptome analyses of insect heads. Appl. Entomol. Zool. 2018, 53, 93–105. [Google Scholar] [CrossRef]
- Ozaki, K.; Utoguchi, A.; Yamada, A.; Yoshikawa, H. Identifcation and genomic structure of chemosensory proteins (CSP) and odorant binding proteins (OBP) genes expressed in foreleg tarsi of the swallowtail butterfy Papilio xuthus. Insect Biochem. Mol. Biol. 2008, 38, 969–976. [Google Scholar] [CrossRef]
- Li, Z.; Zhang, S.; Luo, J.; Zhu, J.; Cui, J.; Dong, S. Expression analysis and binding assays in the chemosensory protein gene family indicate multiple roles in Helicoverpa armigera. J. Chem. Ecol. 2015, 41, 473–485. [Google Scholar] [CrossRef]
- Wang, Q.; Xin, Z.J.; Li, J.C.; Hu, L.F.; Lou, Y.G.; Lu, J. (E)-β-caryophyllene functions as a host location signal for the rice white-backed planthopper Sogatella furcifera. Physiol. Mol. Plant Pathol. 2015, 91, 106–112. [Google Scholar] [CrossRef]
- Mitaka, H.; Matsuo, T.; Miura, N.; Ishikawa, Y. Identification of odorant-binding protein genes from antennal expressed sequence tags of the onion fly, Delia antiqua. Mol. Biol. Rep. 2011, 38, 1787–1792. [Google Scholar] [CrossRef]
- Sun, L.; Wang, Q.; Wang, Q.; Dong, K.; Xiao, Y.; Zhang, Y.J. Identification and characterization of odorant binding proteins in the forelegs of Adelphocoris lineolatus (Goeze). Front. Physiol. 2017, 8, 735. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sun, L.; Wei, Y.; Zhang, D.; Ma, X.; Xiao, Y.; Zhang, Y.; Yang, X.-M.; Xiao, Q.; Guo, Y.-Y.; Zhang, Y.-J. The mouthparts enriched odorant binding protein 11 of the alfalfa plant bug Adelphocoris lineolatus displays a preferential binding behavior to host plant secondary metabolites. Front. Physiol. 2016, 7, 201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xue, W.X.; Fan, J.; Zhang, Y.; Xu, Q.X.; Han, Z.L.; Sun, J.R.; Chen, J.L. Identification and expression analysis of candidate odorant-binding protein and chemosensory protein genes by antennal transcriptome of Sitobion avenae. PLoS ONE 2016, 11, e0161839. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.; Chen, J.L.; Lin, J.H.; Lin, J.T.; Wu, Z.Z. Odorant-binding proteins and chemosensory proteins potentially involved in host plant recognition in the Asian citrus psyllid, Diaphorina citri. Pest Manag. Sci. 2020, 76, 2609–2618. [Google Scholar] [CrossRef]
- Pickett, J.A.; Wadhams, L.J.; Woodcock, C.M. The chemical ecology of aphids. Annu. Rev. Entomol. 1992, 37, 67–90. [Google Scholar] [CrossRef]
- Pickett, J.A.; Glinwood, R.T. Chemical ecology. In Aphids as Crop Pests; Van Emden, H.F., Harrington, R., Eds.; CAB International Press: Oxford, UK, 2007; pp. 235–260. [Google Scholar]
- Chang, H.T.; Liu, Y.; Yang, T.; Pelosi, P.; Dong, S.L.; Wang, G.R. Pheromone binding proteins enhance the sensitivity of olfactory receptors to sex pheromones in Chilo suppressalis. Sci. Rep. 2015, 5, 13093. [Google Scholar] [CrossRef] [Green Version]
- Zhang, R.; Wang, B.; Grossi, G.; Falabella, P.; Liu, Y.; Yan, S.; Lu, J.; Xi, J.; Wang, G. Molecular basis of alarm pheromone detection in aphids. Curr. Biol. 2017, 27, 55–61. [Google Scholar] [CrossRef] [Green Version]
- Lin, X.; Jiang, Y.; Zhang, L.; Cai, Y. Effects of insecticides chlorpyrifos, emamectin benzoate and fipronil on Spodopter litura might be mediated by OBPs and CSPs. Bull. Entomol. Res. 2018, 108, 658–666. [Google Scholar] [CrossRef]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | Gene Name | ORF | SP | Best Blastx Match | |||||
|---|---|---|---|---|---|---|---|---|---|
| (aa) | (aa) | Species | Gene Name | Acc. No | Score | E-Value | Identity (%) | ||
| evm.model.scaffold644.6 | OBP1 | 187 | 17 | Empoasca onukii | OBP6 | AWC68015.1 | 97.6 | 8 × 10−22 | 28.26 |
| TRINITY_DN14345_c0_g1 | OBP2 | 210 | 33 | Empoasca onukii | OBP16 | AWC67989.1 | 258 | 1 × 10−84 | 64.43 |
| TRINITY_DN20712_c0_g1 | OBP3 | 146 | 16 | Empoasca onukii | OBP12 | AWC67992.1 | 145 | 1 × 10−41 | 50.41 |
| TRINITY_DN17302_c1_g2 | OBP4 | 140 | No | Empoasca onukii | OBP8 | AWC67990.1 | 125 | 6 × 10−34 | 42.96 |
| TRINITY_DN20407_c2_g4 | OBP5 | 144 | 19 | Empoasca onukii | OBP42 | AWC67997.1 | 165 | 2 × 10−49 | 61.54 |
| evm.model.scaffold232.48 | OBP6 | 296 | No | Empoasca onukii | OBP13 | AWC68014.1 | 286 | 3 × 10−93 | 60.73 |
| evm.model.scaffold64.23 | OBP7 | 169 | 44 | Empoasca onukii | OBP12 | AWC67992.1 | 176 | 2 × 10−53 | 58.57 |
| TRINITY_DN19892_c0_g6 | OBP8 | 210 | 19 | Empoasca onukii | OBP22 | AWC68011.1 | 165 | 8 × 10−48 | 46.46 |
| evm.model.scaffold64.19 | OBP9 | 149 | 24 | Subpsaltria yangi | OBP2 | AXY87861.1 | 125 | 7 × 10−34 | 42.22 |
| evm.model.scaffold94.54 | OBP10 | 118 | 26 | Empoasca onukii | OBP11 | AWC67987.1 | 124 | 1 × 10−33 | 49.55 |
| TRINITY_DN16837_c1_g7 | OBP11 | 200 | No | Ctenocephalides felis | GOBP 19d-like | XP_026482103.1 | 62.0 | 2 × 10−8 | 30.91 |
| TRINITY_DN16837_c1_g7 | OBP12 | 236 | No | Empoasca onukii | OBP22 | AWC68011.1 | 165 | 8 × 10−48 | 46.46 |
| TRINITY_DN16837_c1_g7 | OBP13 | 143 | 21 | Empoasca onukii | OBP46 | AWC68013.1 | 121 | 7 × 10−31 | 36.02 |
| evm.model.scaffold64.25 | OBP14 | 156 | 23 | Empoasca onukii | OBP38 | AWC68002.1 | 180 | 4 × 10−55 | 64.12 |
| evm.model.scaffold371.21 | OBP15 | 135 | No | Empoasca onukii | OBP49 | AWC68018.1 | 99.0 | 2 × 10−13 | 38.28 |
| TRINITY_DN21214_c0_g2 | OBP16 | 135 | 17 | Empoasca onukii | OBP2 | AWC67994.1 | 188 | 3 × 10−58 | 72.95 |
| TRINITY_DN20748_c1_g13 | OBP17 | 171 | No | Empoasca onukii | OBP42 | AWC67997.1 | 159 | 2 × 10−47 | 58.96 |
| TRINITY_DN2151_c0_g1 | OBP18 | 135 | 16 | Empoasca onukii | OBP42 | AWC67997.1 | 112 | 1 × 10−28 | 45.97 |
| TRINITY_DN17208_c0_g2 | OBP19 | 147 | 20 | Empoasca onukii | OBP5 | AWC67993.1 | 84.3 | 1 × 10−17 | 35.00 |
| TRINITY_DN20849_c1_g2 | OBP20 | 267 | 17 | Subpsaltria yangi | OBP1 | AXY87860.1 | 59.3 | 5 × 10−7 | 27.97 |
| Gene ID | Gene Name | ORF | SP | Best Blastx Match | |||||
|---|---|---|---|---|---|---|---|---|---|
| (aa) | (aa) | Species | Gene Name | Acc. No | Score | E-Value | Identity (%) | ||
| TRINITY_DN19371_c0_g5 | CSP1 | 123 | 18 | Empoasca onukii | CSP9 | AWC68028.1 | 202 | 1 × 10−64 | 77.69 |
| TRINITY_DN20809_c0_g2 | CSP2 | 128 | 17 | Empoasca onukii | CSP13 | AWC68032.1 | 143 | 2 × 10−41 | 57.89 |
| TRINITY_DN19425_c1_g3 | CSP3 | 137 | 24 | Empoasca onukii | CSP14 | AWC68033.1 | 166 | 4 × 10−50 | 60.47 |
| TRINITY_DN18965_c0_g5 | CSP4 | 130 | 19 | Nilaparvata lugens | putative CSP8 | ACJ64054.1 | 152 | 6 × 10−45 | 52.31 |
| TRINITY_DN20331_c0_g1 | CSP5 | 132 | 21 | Empoasca onukii | CSP17 | AWC68036.1 | 166 | 3 × 10−50 | 72.22 |
| TRINITY_DN18304_c1_g6 | CSP6 | 130 | 15 | Empoasca onukii | CSP8 | AWC68026.1 | 194 | 8 × 10−61 | 67.97 |
| evm.model.scaffold1081.12 | CSP7 | 119 | 19 | Empoasca onukii | CSP4 | AWC68023.1 | 147 | 3 × 10−43 | 56.90 |
| TRINITY_DN21169_c0_g11 | CSP8 | 124 | 18 | Empoasca onukii | CSP2 | AWC68021.1 | 124 | 7 × 10−34 | 47.93 |
| TRINITY_DN16691_c0_g1 | CSP9 | 126 | 15 | Empoasca onukii | CSP5 | AWC68022.1 | 167 | 7 × 10−71 | 67.77 |
| TRINITY_DN15719_c0_g3 | CSP10 | 267 | 17 | Nezara viridula | CSP6 | AWC68037.1 | 158 | 4 × 10−45 | 59.13 |
| TRINITY_DN13577_c0_g1 | CSP11 | 109 | 20 | Lygus hesperus | putative CSP1 | APB88037.1 | 161 | 6 × 10−9 | 68.18 |
| TRINITY_DN20737_c0_g2 | CSP12 | 123 | 18 | Empoasca onukii | CSP27 | AWC68029.1 | 178 | 4 × 10−55 | 83.84 |
| evm.model.scaffold312.53 | CSP13 | 127 | 17 | Empoasca onukii | CSP13 | AWC68032.1 | 162 | 1 × 10−48 | 64.41 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chang, X.; Bi, Y.; Chi, H.; Fang, Q.; Lu, Z.; Wang, F.; Ye, G. Identification and Expression Analysis of Odorant-Binding and Chemosensory Protein Genes in Virus Vector Nephotettix cincticeps. Insects 2022, 13, 1024. https://doi.org/10.3390/insects13111024
Chang X, Bi Y, Chi H, Fang Q, Lu Z, Wang F, Ye G. Identification and Expression Analysis of Odorant-Binding and Chemosensory Protein Genes in Virus Vector Nephotettix cincticeps. Insects. 2022; 13(11):1024. https://doi.org/10.3390/insects13111024
Chicago/Turabian StyleChang, Xuefei, Yaluan Bi, Haipeng Chi, Qi Fang, Zhaozhi Lu, Fang Wang, and Gongyin Ye. 2022. "Identification and Expression Analysis of Odorant-Binding and Chemosensory Protein Genes in Virus Vector Nephotettix cincticeps" Insects 13, no. 11: 1024. https://doi.org/10.3390/insects13111024