
Mitochondrial DNA Suggests the Introduction of Honeybees of African Ancestry to East-Central Europe



Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
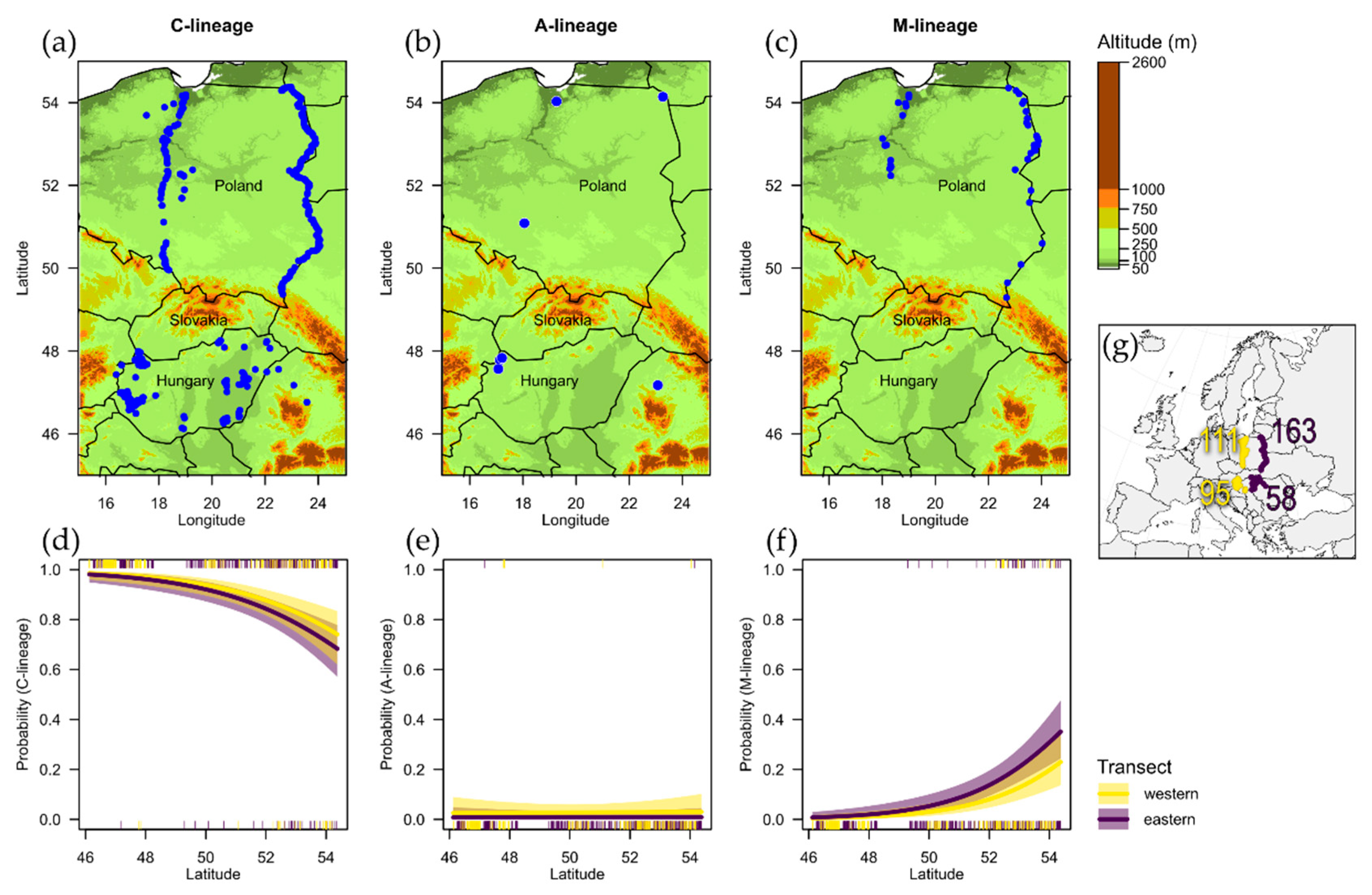
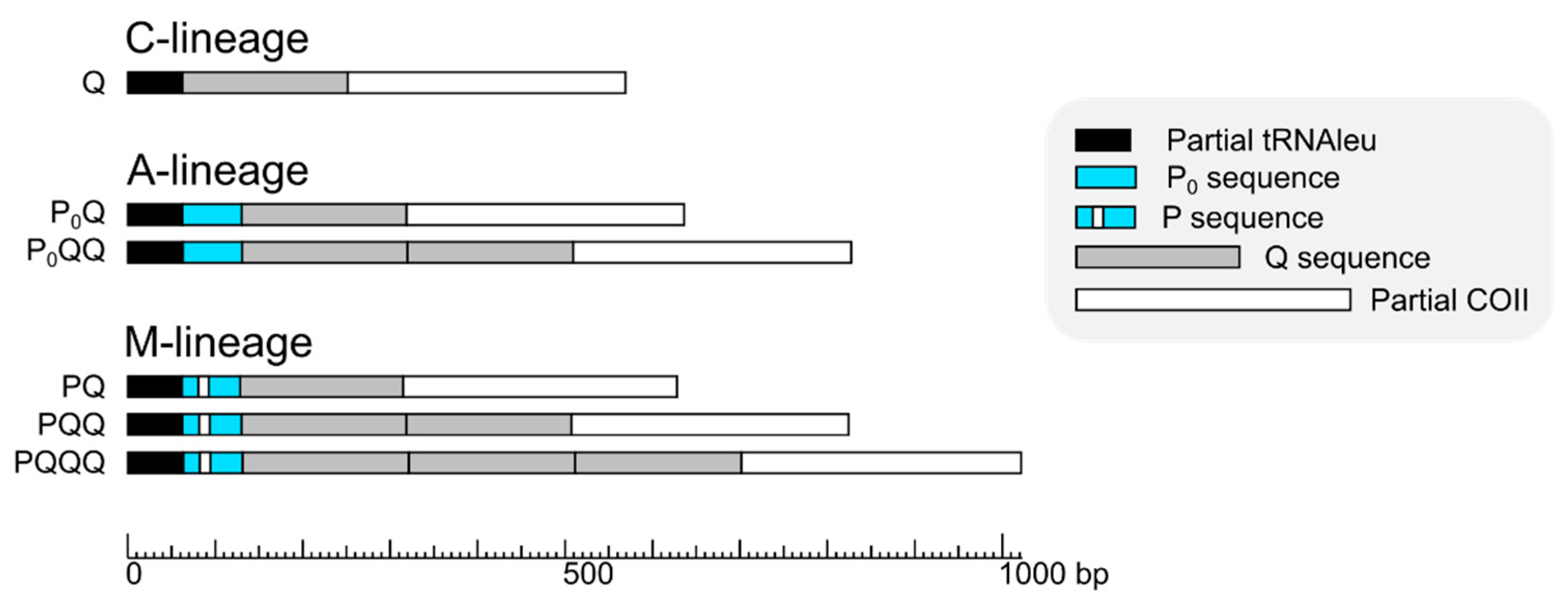
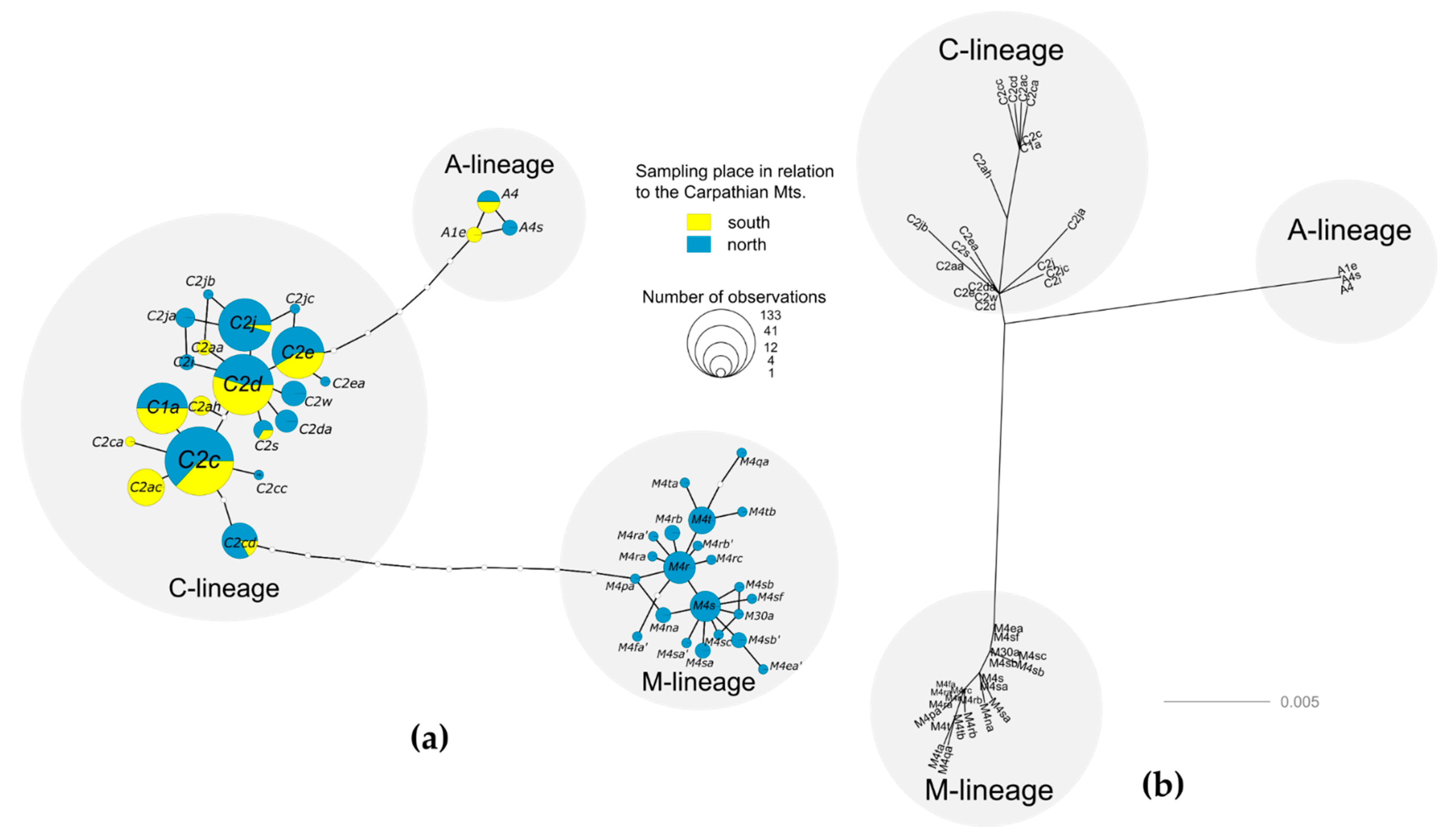
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- De la Rúa, P.; Jaffé, R.; Dall’Olio, R.; Muñoz, I.; Serrano, J. Biodiversity, conservation and current threats to European honeybees. Apidologie 2009, 40, 263–284. [Google Scholar] [CrossRef] [Green Version]
- vanEngelsdorp, D.; Meixner, M.D. A historical review of managed honey bee populations in Europe and the United States and the factors that may affect them. J. Invertebr. Pathol. 2010, 103. [Google Scholar] [CrossRef]
- Zayed, A. Bee genetics and conservation. Apidologie 2009, 40, 237–262. [Google Scholar] [CrossRef] [Green Version]
- Taberlet, P.; Coissac, E.; Pansu, J.; Pompanon, F. Conservation genetics of cattle, sheep, and goats. Comptes Rendus Biol. 2011, 334, 247–254. [Google Scholar] [CrossRef]
- Groeneveld, L.F.; Lenstra, J.A.; Eding, H.; Toro, M.A.; Scherf, B.; Pilling, D.; Negrini, R.; Finlay, E.K.; Jianlin, H.; Groeneveld, E.; et al. Genetic diversity in farm animals—A review. Anim. Genet. 2010, 41, 6–31. [Google Scholar] [CrossRef] [Green Version]
- De la Rúa, P.; Jaffé, R.; Muñoz, I.; Serrano, J.; Moritz, R.F.A.A.; Kraus, F.B. Conserving genetic diversity in the honeybee: Comments on Harpur et al. (2012). Mol. Ecol. 2013, 22, 3208–3210. [Google Scholar] [CrossRef]
- Harpur, B.A.; Minaei, S.; Kent, C.F.; Zayed, A. Management increases genetic diversity of honey bees via admixture. Mol. Ecol. 2012, 21, 4414–4421. [Google Scholar] [CrossRef]
- Jensen, A.B.; Palmer, K.A.; Boomsma, J.J.; Pedersen, B.V. Varying degrees of Apis mellifera ligustica introgression in protected populations of the black honeybee, Apis mellifera mellifera, in northwest Europe. Mol. Ecol. 2005, 14, 93–106. [Google Scholar] [CrossRef] [PubMed]
- Pinto, M.A.; Henriques, D.; Chávez-Galarza, J.; Kryger, P.; Garnery, L.; van der Zee, R.; Dahle, B.; Soland-Reckeweg, G.; de la Rúa, P.; Dall’ Olio, R.; et al. Genetic integrity of the Dark European honey bee (Apis mellifera mellifera) from protected populations: A genome-wide assessment using SNPs and mtDNA sequence data. J. Apic. Res. 2014, 53, 269–278. [Google Scholar] [CrossRef] [Green Version]
- Strange, J.P.; Garnery, L.; Sheppard, W.S. Persistence of the Landes ecotype of Apis mellifera mellifera in southwest France: Confirmation of a locally adaptive annual brood cycle trait. Apidologie 2007, 38, 259–267. [Google Scholar] [CrossRef] [Green Version]
- Oleksa, A.; Chybicki, I.; Tofilski, A.; Burczyk, J. Nuclear and mitochondrial patterns of introgression into native dark bees (Apis mellifera mellifera) in Poland. J. Apic. Res. 2011, 50, 116–129. [Google Scholar] [CrossRef] [Green Version]
- Parejo, M.; Wragg, D.; Gauthier, L.; Vignal, A.; Neumann, P.; Neuditschko, M. Using Whole-Genome Sequence Information to Foster Conservation Efforts for the European Dark Honey Bee, Apis mellifera mellifera. Front. Ecol. Evol. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Arias, M.C.; Sheppard, W.S. Molecular phylogenetics of honey bee subspecies (Apis mellifera L.) inferred from mitochondrial DNA sequence. Mol. Phylogenet. Evol. 1996, 5, 557–566. [Google Scholar] [CrossRef] [PubMed]
- Garnery, L.; Cornuet, J.M.; Solignac, M. Evolutionary history of the honey bee Apis mellifera inferred from mitochondrial DNA analysis. Mol. Ecol. 1992, 1, 145–154. [Google Scholar] [CrossRef]
- Franck, P.; Garnery, L.; Loiseau, A.; Oldroyd, B.P.; Hepburn, H.R.; Solignac, M.; Cornuet, J.M. Genetic diversity of the honeybee in Africa: Microsatellite and mitochondrial data. Heredity (Edinb) 2001, 86, 420–430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wallberg, A.; Han, F.; Wellhagen, G.; Dahle, B.; Kawata, M.; Haddad, N.; Simões, Z.L.P.; Allsopp, M.H.; Kandemir, I.; De la Rúa, P.; et al. A worldwide survey of genome sequence variation provides insight into the evolutionary history of the honeybee Apis mellifera. Nat. Genet. 2014, 46, 1081–1088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garnery, L.; Franck, P.; Baudry, E.; Vautrin, D.; Cornuet, J.-M.; Solignac, M. Genetic diversity of the west European honey bee (Apis mellifera mellifera and A. m. iberica) II. Microsatellite loci. Genet. Sel. Evol. 1998, 30, 49–74. [Google Scholar] [CrossRef]
- Tihelka, E.; Cai, C.; Pisani, D.; Donoghue, P.C.J. Mitochondrial genomes illuminate the evolutionary history of the Western honey bee (Apis mellifera). Sci. Rep. 2020, 10, 1–10. [Google Scholar] [CrossRef]
- Kandemir, İ.; Özkan, A.; Fuchs, S. Reevaluation of honeybee (Apis mellifera) microtaxonomy: A geometric morphometric approach. Apidologie 2011, 42, 618–627. [Google Scholar] [CrossRef] [Green Version]
- Francoy, T.M.; Wittman, D.; Drauschke, M.; Muller, S.; Bezerra-Laure, M.A.F.; De Jong, D.; Goncalves, L.S. Identification of Africanized honey bees through wing morphometrics: Two fast and efficient procedures. Apidologie 2008, 39, 488–494. [Google Scholar] [CrossRef] [Green Version]
- Tofilski, A. Using geometric morphometrics and standard morphometry to discriminate three honeybee subspecies. Apidologie 2008, 39, 558–563. [Google Scholar] [CrossRef] [Green Version]
- Ruttner, F. Biogeography and Taxonomy of Honeybees; Springer: Heidelberg/Berlin, Germany; New York, NY, USA, 1988; ISBN 3-540-17781-7. [Google Scholar]
- Meixner, M.D.; Pinto, M.A.; Bouga, M.; Kryger, P.; Ivanova, E.; Fuchs, S. Standard methods for characterising subspecies and ecotypes of Apis mellifera. J. Apic. Res. 2013, 52, 1–28. [Google Scholar] [CrossRef]
- Sheppard, W.S.; Meixner, M.D. Apis mellifera pomonella, a new honey bee subspecies from Central Asia. Apidologie 2003, 34, 367–375. [Google Scholar] [CrossRef] [Green Version]
- Meixner, M.D.; Leta, M.A.; Koeniger, N.; Fuchs, S. The honey bees of Ethiopia represent a new subspecies of Apis mellifera—Apis mellifera simensis n. ssp. Apidologie 2011, 42, 425–437. [Google Scholar] [CrossRef]
- Hepburn, H.R.; Radloff, S.E. Honeybees of Africa; Springer Science & Business Media: Berlin/Heidelberg, Germany, 2013. [Google Scholar]
- Parker, R.; Melathopoulos, A.P.; White, R.; Pernal, S.F.; Guarna, M.M.; Foster, L.J. Ecological Adaptation of Diverse Honey Bee (Apis mellifera) Populations. PLoS ONE 2010, 5, e11096. [Google Scholar] [CrossRef]
- Büchler, R.; Costa, C.; Hatjina, F.; Andonov, S.; Meixner, M.D.; Le Conte, Y.; Uzunov, A.; Berg, S.; Bienkowska, M.; Bouga, M.; et al. The influence of genetic origin and its interaction with environmental effects on the survival of Apis mellifera L. colonies in Europe. J. Apic. Res. 2014, 53, 205–214. [Google Scholar] [CrossRef] [Green Version]
- Pinto, M.A.; Sheppard, W.S.; Johnston, J.S.; Rubink, W.L.; Coulson, R.N.; Schiff, N.M.; Kandemir, I.; Patton, J.C. Honey Bees (Hymenoptera: Apidae) of African Origin Exist in Non-Africanized Areas of the Southern United States: Evidence from Mitochondrial DNA. Ann. Entomol. Soc. Am. 2007, 100, 289–295. [Google Scholar] [CrossRef] [Green Version]
- Smith, D.R.; Palopoli, M.F.; Taylor, B.R.; Garnery, L.; Cornuet, J.-M.; Solignac, M.; Brown, W.M. Geographical Overlap of Two Mitochondrial Genomes in Spanish Honeybees (Apis mellifera iberica). J. Hered. 1991, 82, 96–100. [Google Scholar] [CrossRef]
- Miguel, I.; Iriondo, M.; Garnery, L.; Sheppard, W.S.; Estonba, A. Gene flow within the M evolutionary lineage of Apis mellifera: Role of the Pyrenees, isolation by distance and post-glacial re-colonization routes in the western Europe. Apidologie 2007, 38, 141–155. [Google Scholar] [CrossRef]
- Cánovas, F.; De la Rúa, P.; Serrano, J.; Galián, J.; Rúa, P.D. La Geographical patterns of mitochondrial DNA variation in Apis mellifera iberiensis (Hymenoptera: Apidae). J. Zool. Syst. Evol. Res. 2007, 46, 24–30. [Google Scholar] [CrossRef]
- Chávez-Galarza, J.; Garnery, L.; Henriques, D.; Neves, C.J.; Loucif-Ayad, W.; Jonhston, J.S.; Pinto, M.A. Mitochondrial DNA variation of Apis mellifera iberiensis: Further insights from a large-scale study using sequence data of the tRNAleu-cox2 intergenic region. Apidologie 2017, 48, 533–544. [Google Scholar] [CrossRef] [Green Version]
- Garnery, L.; Mosshine, E.H.; Oldroyd, B.P.; Cornuet, J.M. Mitochondrial DNA variation in Moroccan and Spanish honey bee populations. Mol. Ecol. 1995, 4, 465–472. [Google Scholar] [CrossRef]
- Pinto, M.A.; Henriques, D.; Neto, M.; Guedes, H.; Muñoz, I.; Azevedo, J.C.; de la Rúa, P. Maternal diversity patterns of Ibero-Atlantic populations reveal further complexity of Iberian honeybees. Apidologie 2013, 44, 430–439. [Google Scholar] [CrossRef] [Green Version]
- Benstead, E. Genetic composition and phenology of mating drone congregations in the honey bee Apis melllfera. Rev. d’Ecologie (La Terre la Vie) 2009, 64, 343–350. [Google Scholar]
- Franck, P.; Garnery, L.; Celebrano, G.; Solignac, M.; Cornuet, J.-M. Hybrid origins of honeybees from Italy (Apis mellifera ligustica) and Sicily (A. m. sicula). Mol. Ecol. 2000, 9, 907–921. [Google Scholar] [CrossRef] [PubMed]
- Zammit-Mangion, M.; Meixner, M.; Mifsud, D.; Sammut, S.; Camilleri, L. Thorough morphological and genetic evidence confirm the existence of the endemic honey bee of the Maltese Islands Apis mellifera ruttneri: Recommendations for conservation. J. Apic. Res. 2017, 56, 514–522. [Google Scholar] [CrossRef]
- De La Rúa, P.; Galián, J.; Serrano, J.; Moritz, R.F.A. Molecular characterization and population structure of the honeybees from the balearic islands (Spain). Apidologie 2001, 32, 417–427. [Google Scholar] [CrossRef] [Green Version]
- De la Rúa, P.; Serrano, J.; Galián, J. Mitochondrial DNA variability in the Canary Islands honeybees (Apis mellifera L.). Mol. Ecol. 1998, 7, 1543–1547. [Google Scholar] [CrossRef] [PubMed]
- de la Rúa, P.; Galián, J.; Pedersen, B.V.; Serrano, J. Molecular characterization and population structure of Apis mellifera from Madeira and the Azores. Apidologie 2006, 37, 699–708. [Google Scholar] [CrossRef] [Green Version]
- Lodesani, M.; Costa, C. Bee breeding and genetics in Europe. Bee World 2003, 84, 69–85. [Google Scholar] [CrossRef]
- Meixner, M.D.; Costa, C.; Kryger, P.; Hatjina, F.; Bouga, M.; Ivanova, E.; Büchler, R. Conserving diversity and vitality for honey bee breeding. J. Apic. Res. 2010, 49, 85–92. [Google Scholar] [CrossRef]
- Bornus, I.; Demianowicz, A.; Gromisz, M. Morfometryczne badania krajowej pszczoly miodnej. Pszczel. Zesz. Nauk. 1966, 10, 1–46. [Google Scholar]
- Meixner, M.D.; Worobik, M.; Wilde, J.; Fuchs, S.; Koeniger, N. Apis mellifera mellifera in eastern Europe—morphometric variation and determination of its range limits. Apidologie 2007, 38, 191–197. [Google Scholar] [CrossRef]
- Péntek-Zakar, E.; Oleksa, A.; Borowik, T.; Kusza, S. Population structure of honey bees in the Carpathian Basin (Hungary) confirms introgression from surrounding subspecies. Ecol. Evol. 2015, 5, 5456–5467. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Oleksa, A.; Tofilski, A. Wing geometric morphometrics and microsatellite analysis provide similar discrimination of honey bee subspecies. Apidologie 2015, 46, 49–60. [Google Scholar] [CrossRef] [Green Version]
- Pew, J.; Wang, J.; Muir, P.; Frasier, T. Related: An R package for Analyzing Pairwise Relatedness Data Based on Codominant Molecular Markers. R Package Version 0.8/r2. 2014. Available online: https://R-Forge.R-project.org/projects/related/ (accessed on 20 March 2021).
- Cornuet, J.M.; Garnery, L.; Solignac, M. Putative origin and function of the intergenic region between COI and COII of Apis mellifera L. mitochondrial DNA. Genetics 1991, 128, 393–403. [Google Scholar] [CrossRef]
- Cornuet, J.M.; Garnery, L. Mitochondrial DNA variability in honeybees and its phylogeographic implications. Apidologie 1991, 22, 627–642. [Google Scholar] [CrossRef] [Green Version]
- Garnery, L.; Solignac, M.; Celebrano, G.; Cornuet, J.M. A simple test using restricted PCR-amplifled mitochondrial DNA to study the genetic structure of Apis mellifera L. Experientia 1993, 49, 1016–1021. [Google Scholar] [CrossRef]
- Rortais, A.; Arnold, G.; Alburaki, M.; Legout, H.; Garnery, L. Review of the DraI COI-COII test for the conservation of the black honeybee (Apis mellifera mellifera). Conserv. Genet. Resour. 2011, 3, 383–391. [Google Scholar] [CrossRef]
- Hall, T.T.A. BioEdit: A user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symp. Ser. 1999, 41, 95–98. [Google Scholar]
- Katoh, K.; Rozewicki, J.; Yamada, K.D. MAFFT online service: Multiple sequence alignment, interactive sequence choice and visualization. Brief. Bioinform. 2017. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Simmons, M.P.; Ochoterena, H. Gaps as Characters in Sequence-Based Phylogenetic Analyses. Syst. Biol. 2000, 49, 369–381. [Google Scholar] [CrossRef] [Green Version]
- Aktas, C. Package “haplotypes”: Haplotype Inference and Statistical Analysis of Genetic Variation; R Package ver. 1.0. R Foundation for Statistical Computing: Vienna, Austria, 2015. Available online: https://cran.r-project.org/web/packages/haplotypes/ (accessed on 20 March 2021).
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular Evolutionary Genetics Analysis across Computing Platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef] [PubMed]
- Hasegawa, M.; Kishino, H.; Yano, T. aki Dating of the human-ape splitting by a molecular clock of mitochondrial DNA. J. Mol. Evol. 1985, 22, 160–174. [Google Scholar] [CrossRef] [PubMed]
- Minh, B.Q.; Schmidt, H.A.; Chernomor, O.; Schrempf, D.; Woodhams, M.D.; von Haeseler, A.; Lanfear, R. IQ-TREE 2: New Models and Efficient Methods for Phylogenetic Inference in the Genomic Era. Mol. Biol. Evol. 2020, 37, 1530–1534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorai-Raj, S. binom: Binomial Confidence Intervals for Several Parameterizations. R Package Version 1.1-1. 2014. Available online: https://CRAN.R-project.org/package=binom (accessed on 20 March 2021).
- Muñoz, I.; Dall’olio, R.; Lodesani, M.; De la Rúa, P. Estimating introgression in Apis mellifera siciliana populations: Are the conservation islands really effective? Insect Conserv. Divers. 2014. [Google Scholar] [CrossRef]
- Bertrand, B.; Alburaki, M.; Legout, H.; Moulin, S.; Mougel, F.; Garnery, L. MtDNA COI-COII marker and drone congregation area: An efficient method to establish and monitor honeybee (Apis mellifera L.) conservation centres. Mol. Ecol. Resour. 2015, 15, 673–683. [Google Scholar] [CrossRef]
- Božič, J.; Kordiš, D.; Križaj, I.; Leonardi, A.; Močnik, R.; Nakrst, M.; Podgoršek, P.; Prešern, J.; Sušnik Bajec, S.; Zorc, M.; et al. Novel aspects in characterisation of Carniolan honey bee (Apis mellifera carnica, Pollmann 1879). Acta Agric. Slov. 2016, 5 (Suppl. 5), 18–27. [Google Scholar]
- Eimanifar, A.; Kimball, R.T.; Braun, E.L.; Ellis, J.D. Mitochondrial genome diversity and population structure of two western honey bee subspecies in the Republic of South Africa. Sci. Rep. 2018, 8, 1333. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Szalanski, A.L.; Magnus, R.M. Mitochondrial DNA characterization of Africanized honey bee (Apis mellifera L.) populations from the USA. J. Apic. Res. 2010, 49, 177–185. [Google Scholar] [CrossRef] [Green Version]
- Collet, T.; Ferreira, K.M.; Arias, M.C.; Soares, A.E.E.; Del Lama, M.A. Genetic structure of Africanized honeybee populations (Apis mellifera L.) from Brazil and Uruguay viewed through mitochondrial DNA COI–COII patterns. Hered (Edinb) 2006, 97, 329–335. [Google Scholar] [CrossRef] [Green Version]
- Han, F.; Wallberg, A.; Webster, M.T. From where did the Western honeybee (Apis mellifera) originate? Ecol. Evol. 2012, 2, 1949–1957. [Google Scholar] [CrossRef] [PubMed]
- Moritz, R.F.A.; Mautz, D. Development of Varroa jacobsoni in colonies of Apis mellifera capensis and Apis mellifera carnica. Apidologie 1990, 21, 53–58. [Google Scholar] [CrossRef] [Green Version]
- Köppler, K.; Vorwohl, G.; Koeniger, N. Comparison of pollen spectra collected by four different subspecies of the honey bee Apis mellifera. Apidologie 2007, 38, 341–353. [Google Scholar] [CrossRef] [Green Version]
- Woyke, J. Experiences with Apis mellifera adansonii in Brazil and in Poland. Apiacta 1973, 8, 115–116. [Google Scholar]
- Woyke, J.; Jasiński, Z.; Smagowska, B. Badania nad międzyrasowymi mieszańcami pszczoły miodnej. II. Porównanie organów rozrodczych oraz efektów sztucznego i naturalnego unasieniania pszczół różnych ras i ich mieszańców. Pszczel. Zesz. Nauk. 1974, 18, 53–75. [Google Scholar]
- De La Rúa, P.; Jiménez, Y.; Galián, J.; Serrano, J. Evaluation of the biodiversity of honey bee (Apis mellifera) populations from eastern Spain. J. Apic. Res. 2004, 43, 162–166. [Google Scholar] [CrossRef]
- Magnus, R.M.; Tripodi, A.D.; Szalanski, A.L. Mitochondrial DNA diversity of honey bees, Apis mellifera L. (Hymenoptera: Apidae) from queen breeders in the United States. J. Apic. Sci. 2011, 55, 37–47. [Google Scholar]
- Susnik, S.; Kozmus, P.; Poklukar, J.; Meglic, V. Molecular characterisation of indigenous Apis mellifera carnica in Slovenia. Apidologie 2004, 35, 623–636. [Google Scholar] [CrossRef] [Green Version]
- Kralj, J. Selection of Honey Bees with Rapid Development as a Component of Varroa Mite Resistance; University of Guelph: Guelph, ON, Canada, 1998. [Google Scholar]
- Österlund, E. Exploring Monticola—Efforts to find an acceptable Varroa-resistant honey bee. Am. Bee J. 1991, 131, 49–56. [Google Scholar]
- Okuyama, H.; Hill, J.; Martin, S.J.; Takahashi, J. The complete mitochondrial genome of a Buckfast bee, Apis mellifera (Insecta: Hymenoptera: Apidae) in Northern Ireland. Mitochondrial DNA Part B 2018, 3, 338–339. [Google Scholar] [CrossRef] [Green Version]
- Strange, J.P.; Garnery, L.; Sheppard, W.S. Morphological and molecular characterization of the Landes honey bee (Apis mellifera L.) ecotype for genetic conservation. J. Insect Conserv. 2008, 12, 527–537. [Google Scholar] [CrossRef]
- Šťastný, M.; Gasper, J.; Bauer, M. Genetic structure of Apis mellifera carnica in Slovakia based on microsatellite DNA polymorphism. Biologia (Bratisl) 2017, 72, 1341–1346. [Google Scholar] [CrossRef]
- Learner, J. Imports and Exports of Honey Bees. Bee Cr. 2017, 99, 7–10. [Google Scholar]
- Bieńkowska, M.; Wilde, J.; Panasiuk, B.; Gerula, D. Bee breeding activity in Poland. In Proceedings of the SICAMM 2018 Conference, Mustiala, Finland, 13–15 July 2018; Organized by the Finnish Beekeepers’ Association at HAMK, Häme University of Applied Sciences. p. 6. [Google Scholar]
- Soland-Reckeweg, G.; Heckel, G.; Neumann, P.; Fluri, P.; Excoffier, L. Gene flow in admixed populations and implications for the conservation of the Western honeybee, Apis mellifera. J. Insect Conserv. 2008, 13, 317–328. [Google Scholar] [CrossRef] [Green Version]
- Techer, M.A.; Clémencet, J.; Simiand, C.; Turpin, P.; Garnery, L.; Reynaud, B.; Delatte, H. Genetic diversity and differentiation among insular honey bee populations in the southwest Indian Ocean likely reflect old geographical isolation and modern introductions. PLoS ONE 2017, 12, e0189234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- De la Rúa, P.; Serrano, J.; Galián, J. Biodiversity of Apis mellifera populations from Tenerife (Canary Islands) and hybridisation with East European races. Biodivers. Conserv. 2002, 11, 59–67. [Google Scholar] [CrossRef]
- Winston, M.L.; Otis, G.W.; Taylor, O.R. Absconding Behaviour of the Africanized Honeybee in South America. J. Apic. Res. 1979, 18, 85–94. [Google Scholar] [CrossRef]
- Martin, S.; Wossler, T.; Kryger, P. Usurpation of African Apis mellifera scutellata colonies by parasitic Apis mellifera capensis workers. Apidologie 2002, 33, 215–232. [Google Scholar] [CrossRef] [Green Version]
- Muli, E.; Patch, H.; Frazier, M.; Frazier, J.; Torto, B.; Baumgarten, T.; Kilonzo, J.; Kimani, J.N.; Mumoki, F.; Masiga, D.; et al. Evaluation of the Distribution and Impacts of Parasites, Pathogens, and Pesticides on Honey Bee (Apis mellifera) Populations in East Africa. PLoS ONE 2014, 9, e94459. [Google Scholar] [CrossRef]
- McDowell, R. The Africanized Honey Bee in the United States: What will Happen to the US Beekeeping Industry? Agricultural Economic Report; United States Dept. of Agriculture (USA): Washington, DC, USA, 1984.
- Taylor, O.R. African Bees: Potential Impact in the United States. Bull. Entomol. Soc. Am. 1985, 31, 15–24. [Google Scholar] [CrossRef]
- Winston, M.L. Killer Bees. The Africanized Honey Bee in the Americas; Harvard University Press: Cambridge, MA, USA, 1992; ISBN 067450352X. [Google Scholar]
- Byatt, M.A.; Chapman, N.C.; Latty, T.; Oldroyd, B.P. The genetic consequences of the anthropogenic movement of social bees. Insectes Soc. 2016, 63, 15–24. [Google Scholar] [CrossRef]
- Livanis, G.; Moss, C.B. The effect of Africanized honey bees on honey production in the United States: An informational approach. Ecol. Econ. 2010, 69, 895–904. [Google Scholar] [CrossRef]
- Villa, J.D. Defensive Behaviour of Africanized and European Honeybees at two Elevations in Colombia. J. Apic. Res. 1988, 27, 141–145. [Google Scholar] [CrossRef]
- Ferreira, R.S.; Almeida, R.A.M.B.; Barraviera, S.R.C.S.; Barraviera, B. Historical Perspective and Human Consequences of Africanized Bee Stings in the Americas. J. Toxicol. Environ. Health Part B 2012, 15, 97–108. [Google Scholar] [CrossRef] [PubMed]
- Moritz, R.F.A.; Härtel, S.; Neumann, P. Global invasions of the western honeybee (Apis mellifera) and the consequences for biodiversity. Écoscience 2005, 12, 289–301. [Google Scholar] [CrossRef]
- Smith, D.R. African bees in the Americas: Insights from biogeography and genetics. Trends Ecol. Evol. 1991, 6, 17–21. [Google Scholar] [CrossRef] [Green Version]
- Pinto, M.A. Africanization in the United States: Replacement of Feral European Honeybees (Apis mellifera L.) by an African Hybrid Swarm. Genetics 2005, 170, 1653–1665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sylvester, H.A.; Rinderer, T.E.; Shimanuki, H. Certification options for dealing with Africanized bees. Am. Bee J. 1992, 132, 182–184. [Google Scholar]
- Page, R.E.; Erickson, E.H. Identification and Certification of Africanized Honey Bees. Ann. Entomol. Soc. Am. 1985, 78, 149–158. [Google Scholar] [CrossRef]
- Abrahamovich, A.H.; Atela, O.; De la Rúa, P.; Galián, J. Assessment of the mitochondrial origin of honey bees from Argentina. J. Apic. Res. 2007, 46, 191–194. [Google Scholar] [CrossRef]
- Winston, M.L.; Mitchell, S.R.; Punnett, E.N. Feasibility of Package Honey Bee (Hymenoptera: Apidae) Production in Southwestern British Columbia, Canada. J. Econ. Entomol. 1985, 78, 1037–1041. [Google Scholar] [CrossRef]
- Armitage, P. Regulatory Framework for the Importation of Honey Bees in Canada. BeesCene 2018, 34, 42–47. [Google Scholar]
- Harrison, P.A.; Berry, P.M.; Butt, N.; New, M. Modelling climate change impacts on species’ distributions at the European scale: Implications for conservation policy. Environ. Sci. Policy 2006, 9, 116–128. [Google Scholar] [CrossRef]
- Moretto, G.; Gonçalves, L.S.; De Jong, D.; Bichuette, M.Z. The effects of climate and bee race on Varroa jacobsoni Oud infestations in Brazil. Apidologie 1991, 22, 197–203. [Google Scholar] [CrossRef] [Green Version]
- Arechavaleta-Velasco, M.E.; Guzmán-Novoa, E.; Guzman-Novoa, E. Relative effect of four characteristics that restrain the population growth of the mite Varroa destructor in honey bee (Apis mellifera) colonies. Apidologie 2001, 32, 157–174. [Google Scholar] [CrossRef] [Green Version]
- Guzman-Novoa, E.; Emsen, B.; Unger, P.; Espinosa-Montaño, L.G.; Petukhova, T. Genotypic variability and relationships between mite infestation levels, mite damage, grooming intensity, and removal of Varroa destructor mites in selected strains of worker honey bees (Apis mellifera L.). J. Invertebr. Pathol. 2012, 110, 314–320. [Google Scholar] [CrossRef]
- Rosenkranz, P. Honey bee (Apis mellifera L.) tolerance to Varroa jacobsoni Oud. in South America. Apidologie 1999, 30, 159–172. [Google Scholar] [CrossRef]
- Carreck, N.L. Breeding honey bees for varroa tolerance. In Varroa-Still a Problem in the 21th Century; Carreck, N.L., Ed.; International Bee Research Association: Cardiff, UK, 2011; pp. 60–66. [Google Scholar]
- Corrêa-Marques, M.H.; De Jong, D.; Rosenkranz, P.; Gonçalves, L.S. Varroa-tolerant Italian honey bees introduced from Brazil were not more efficient in defending themselves against the mite Varroa destructor than Carniolan bees in Germany. Genet. Mol. Res. 2002, 1, 153–158. [Google Scholar] [PubMed]
- Locke, B. Natural Varroa mite-surviving Apis mellifera honeybee populations. Apidologie 2016, 47, 467–482. [Google Scholar] [CrossRef] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| Lineage | Sequence Type | Estimated Frequency (95% Confidence Intervals) |
|---|---|---|
| C | Q | 0.881 (0.846–0.910) |
| all C | 0.881 (0.846–0.910) | |
| A | P0Q | 0.005 (0.001–0.017) |
| P0QQ | 0.012 (0.004–0.027) | |
| all A | 0.016 (0.007–0.033) | |
| M | PQ | 0.002 (0.000–0.013) |
| PQQ | 0.084 (0.060–0.115) | |
| PQQQ | 0.016 (0.007–0.033) | |
| all M | 0.103 (0.076–0.136) |
| C-Lineage | A-Lineage | M-Lineage | |
|---|---|---|---|
| Latitude | −0.383 *** | −0.044 | 0.535 *** |
| (0.079) | (0.136) | (0.108) | |
| Transect # | 0.340 | 0.962 | −0.615 + |
| (0.319) | (0.852) | (0.356) | |
| Constant | 21.578 *** | −2.441 | −29.638 *** |
| (4.155) | (6.947) | (5.681) | |
| Model fit χ2 (2) | 35.48 *** | 1.68 | 48.17 *** |
| Pseudo-R2 (McFadden) | 0.114 | 0.024 | 0.170 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Oleksa, A.; Kusza, S.; Tofilski, A. Mitochondrial DNA Suggests the Introduction of Honeybees of African Ancestry to East-Central Europe. Insects 2021, 12, 410. https://doi.org/10.3390/insects12050410
Oleksa A, Kusza S, Tofilski A. Mitochondrial DNA Suggests the Introduction of Honeybees of African Ancestry to East-Central Europe. Insects. 2021; 12(5):410. https://doi.org/10.3390/insects12050410
Chicago/Turabian StyleOleksa, Andrzej, Szilvia Kusza, and Adam Tofilski. 2021. "Mitochondrial DNA Suggests the Introduction of Honeybees of African Ancestry to East-Central Europe" Insects 12, no. 5: 410. https://doi.org/10.3390/insects12050410