
Temporal and Spatial Blood Feeding Patterns of Urban Mosquitoes in the San Juan Metropolitan Area, Puerto Rico

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
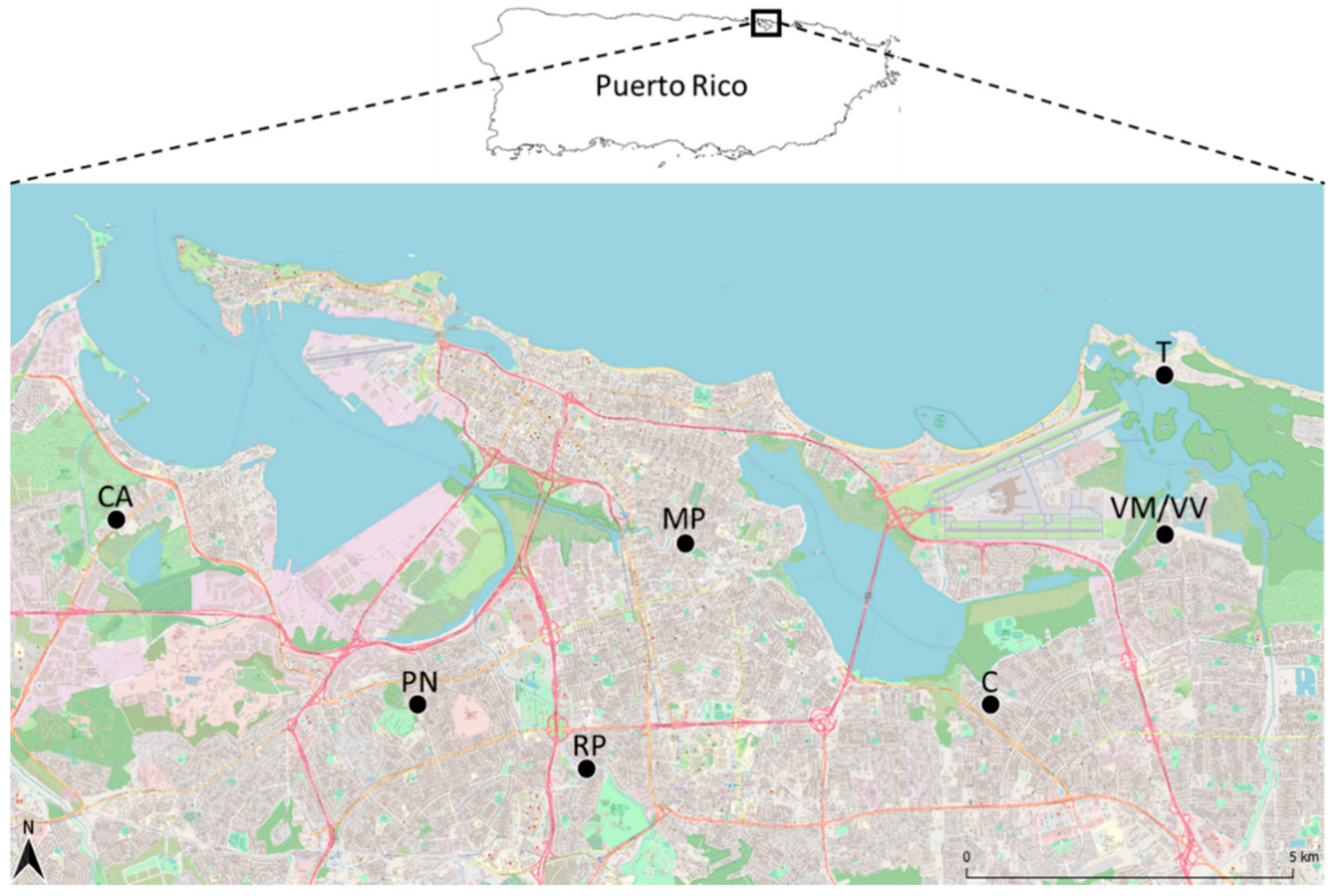
2.1. Study Site
2.2. Human and Landscape Variables
2.3. Mosquito Adult Sampling and Sample Preservation
2.4. DNA Extraction
2.5. Molecular Verification of Mosquito Species
2.6. DNA Metabarcoding
2.7. Vertebrate Species Verification of Inconclusive Samples
2.8. Bioinformatics and Statistical Analysis
3. Results
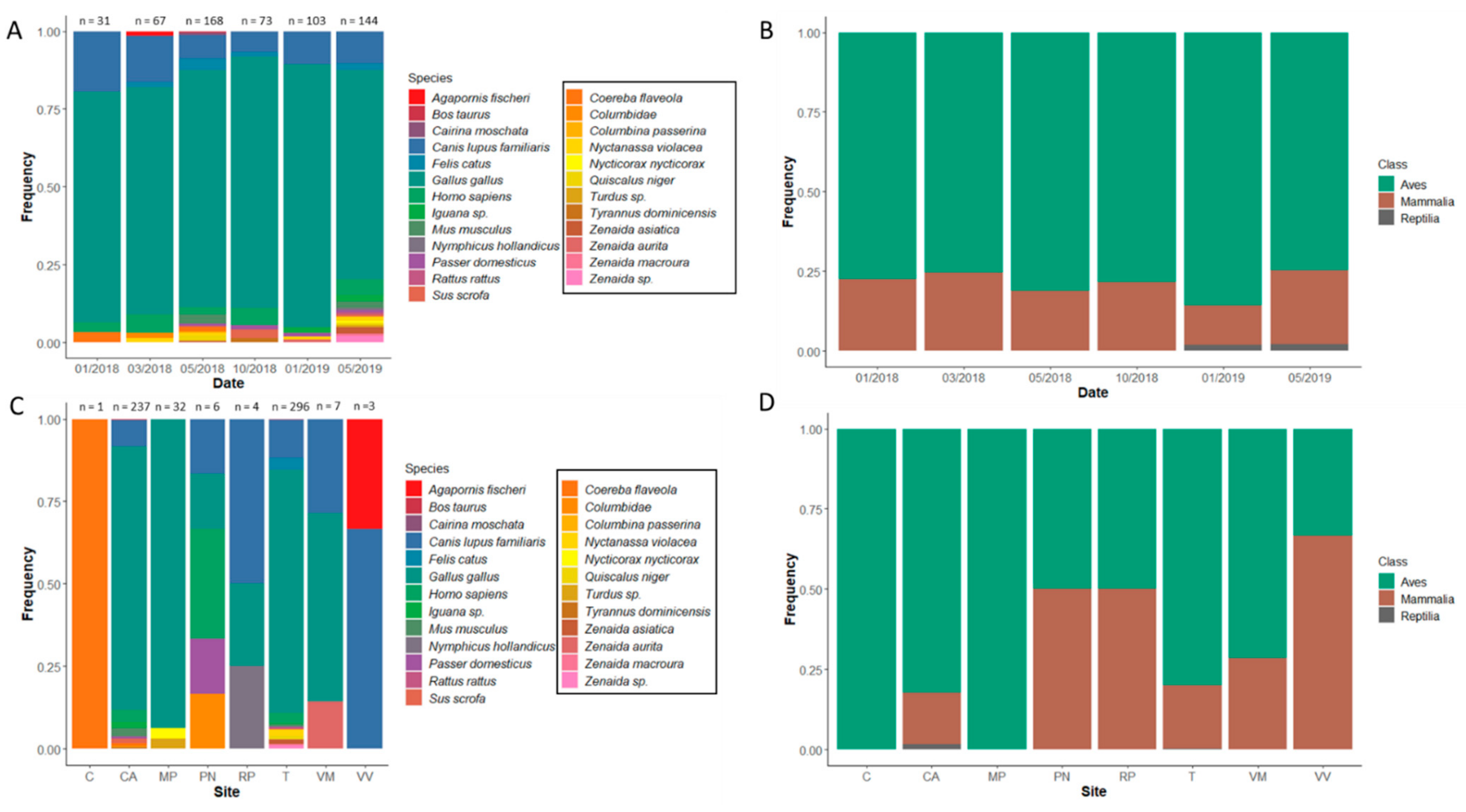
3.1. Blood-Engorged Mosquito Collection and Identification
3.2. Molecular Blood Meal Identification
4. Discussion
4.1. Temporal and Spatial Host Patterns
4.2. Spatial Host Patterns
4.3. General Blood Meal Findings
4.4. Implications for Zoonotic Pathogen Transmission
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Angold, P.G.; Sadler, J.P.; Hill, M.O.; Pullin, A.; Rushton, S.; Austin, K.; Small, E.; Wood, B.; Wadsworth, R.; Sanderson, R.; et al. Biodiversity in urban habitat patches. Sci. Total Environ. 2006, 360, 196–204. [Google Scholar] [CrossRef] [PubMed]
- Jones, E.L.; Leather, S.R. Invertebrates in urban areas: A review. EJE 2013, 109, 463–478. [Google Scholar] [CrossRef] [Green Version]
- McIntyre, N.E. Ecology of Urban Arthropods: A Review and a Call to Action. Ann. Entomol. Soc. Am. 2000, 93, 825–835. [Google Scholar] [CrossRef]
- LaDeau, S.L.; Allan, B.F.; Leisnham, P.T.; Levy, M.Z. The ecological foundations of transmission potential and vector-borne disease in urban landscapes. Funct. Ecol. 2015, 29, 889–901. [Google Scholar] [CrossRef] [Green Version]
- Cox, J.; Grillet, M.E.; Ramos, O.M.; Amador, M.; Barrera, R. Habitat segregation of dengue vectors along an urban environmental gradient. Am. J. Trop. Med. Hyg. 2007, 76, 820–826. [Google Scholar] [CrossRef]
- Johnston, E.; Weinstein, P.; Slaney, D.; Flies, A.S.; Fricker, S.; Williams, C. Mosquito communities with trap height and urban-rural gradient in Adelaide, South Australia: Implications for disease vector surveillance. J. Vector Ecol. 2014, 39, 48–55. [Google Scholar] [CrossRef]
- Valdez, M.R.W.d. Mosquito Species Distribution Across Urban, Suburban, and Semi-Rural Residences in San Antonio, Texas. J. Vector Ecol. 2017, 42, 184–188. [Google Scholar] [CrossRef] [Green Version]
- de Jesús Crespo, R.; Lázaro, P.M.; Yee, S.H. Linking wetland ecosystem services to vector-borne disease: Dengue fever in the San Juan Bay estuary, Puerto Rico. Wetlands 2019, 39, 1281–1293. [Google Scholar] [CrossRef]
- Goodman, H.; Egizi, A.; Fonseca, D.M.; Leisnham, P.T.; LaDeau, S.L. Primary blood-hosts of mosquitoes are influenced by social and ecological conditions in a complex urban landscape. Parasites Vectors 2018, 11, 218. [Google Scholar] [CrossRef]
- Valentini, A.; Taberlet, P.; Miaud, C.; Civade, R.; Herder, J.; Thomsen, P.F.; Bellemain, E.; Besnard, A.; Coissac, E.; Boyer, F.; et al. Next-generation monitoring of aquatic biodiversity using environmental DNA metabarcoding. Mol. Ecol. 2016, 25, 929–942. [Google Scholar] [CrossRef] [Green Version]
- Piaggio, A.J.; Engeman, R.M.; Hopken, M.W.; Humphrey, J.S.; Keacher, K.L.; Bruce, W.E.; Avery, M.L. Detecting an elusive invasive species: A diagnostic PCR to detect Burmese python in Florida waters and an assessment of persistence of environmental DNA. Mol. Ecol. Resour. 2014, 14, 374–380. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ficetola, G.F.; Miaud, C.; Pompanon, F.; Taberlet, P. Species detection using environmental DNA from water samples. Biol. Lett. 2008, 4, 423–425. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Calvignac-Spencer, S.; Leendertz, F.H.; Gilbert, M.T.P.; Schubert, G. An invertebrate stomach’s view on vertebrate ecology. BioEssays 2013, 35, 1004–1013. [Google Scholar] [CrossRef] [PubMed]
- Calvignac-Spencer, S.; Merkel, K.; Kutzner, N.; Kühl, H.; Boesch, C.; Kappeler, P.M.; Metzger, S.; Schubert, G.; Leendertz, F.H. Carrion fly-derived DNA as a tool for comprehensive and cost-effective assessment of mammalian biodiversity. Mol. Ecol. 2013, 22, 915–924. [Google Scholar] [CrossRef] [PubMed]
- Schnell, I.B.; Thomsen, P.F.; Wilkinson, N.; Rasmussen, M.; Jensen, L.R.D.; Willerslev, E.; Bertelsen, M.F.; Gilbert, M.T.P. Screening mammal biodiversity using DNA from leeches. Curr. Biol. 2012, 22, R262–R263. [Google Scholar] [CrossRef] [Green Version]
- Bohmann, K.; Evans, A.; Gilbert, M.T.P.; Carvalho, G.R.; Creer, S.; Knapp, M.; Yu, D.W.; de Bruyn, M. Environmental DNA for wildlife biology and biodiversity monitoring. Trends Ecol. Evol. 2014, 29, 358–367. [Google Scholar] [CrossRef]
- Taberlet, P.; Coissac, E.; Pompanon, F.; Brochmann, C.; Willerslev, E. Towards next-generation biodiversity assessment using DNA metabarcoding. Mol. Ecol. 2012, 21, 2045–2050. [Google Scholar] [CrossRef]
- Galan, M.; Pons, J.B.; Tournayre, O.; Pierre, É.; Leuchtmann, M.; Pontier, D.; Charbonnel, N. Metabarcoding for the parallel identification of several hundred predators and their prey: Application to bat species diet analysis. Mol. Ecol. Resour. 2018, 18, 474–489. [Google Scholar] [CrossRef]
- Beng, K.C.; Tomlinson, K.W.; Shen, X.H.; Surget-Groba, Y.; Hughes, A.C.; Corlett, R.T.; Slik, J.W.F. The utility of DNA metabarcoding for studying the response of arthropod diversity and composition to land-use change in the tropics. Sci. Rep. 2016, 6, 24965. [Google Scholar] [CrossRef]
- Logue, K.; Keven, J.B.; Cannon, M.V.; Reimer, L.; Siba, P.; Walker, E.D.; Zimmerman, P.A.; Serre, D. Unbiased characterization of Anopheles mosquito blood meals by targeted high-throughput sequencing. PLoS Negl. Trop. Dis. 2016, 10, e0004512. [Google Scholar] [CrossRef]
- Kent, R.J. Molecular methods for arthropod bloodmeal identification and applications to ecological and vector-borne disease studies. Mol. Ecol. Resour. 2009, 9, 4–18. [Google Scholar] [CrossRef] [PubMed]
- Garrett-Jones, C. Prognosis for interruption of malaria transmission through assessment of the mosquito’s vectorial capacity. Nature 1964, 204, 1173–1175. [Google Scholar] [CrossRef] [PubMed]
- Lyimo, I.N.; Ferguson, H.M. Ecological and evolutionary determinants of host species choice in mosquito vectors. Trends Parasitol. 2009, 25, 189–196. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.; Amador, M.; Barrera, R. Seasonal and habitat effects on dengue and West Nile virus vectors in San Juan, Puerto Rico. J. Am. Mosq. Control. Assoc. 2009, 25, 38–46. [Google Scholar] [CrossRef] [PubMed]
- Medeiros-Sousa, A.R.; Fernandes, A.; Ceretti-Junior, W.; Wilke, A.B.B.; Marrelli, M.T. Mosquitoes in urban green spaces: Using an island biogeographic approach to identify drivers of species richness and composition. Sci. Rep. 2017, 7, 17826. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Little, E.; Biehler, D.; Leisnham, P.T.; Jordan, R.; Wilson, S.; LaDeau, S.L. Socio-ecological mechanisms supporting high densities of Aedes albopictus (Diptera: Culicidae) in Baltimore, MD. J. Med. Entomol. 2017, 54, 1183–1192. [Google Scholar] [CrossRef] [Green Version]
- Zittra, C.; Vitecek, S.; Obwaller, A.G.; Rossiter, H.; Eigner, B.; Zechmeister, T.; Waringer, J.; Fuehrer, H.P. Landscape structure affects distribution of potential disease vectors (Diptera: Culicidae). Parasites Vectors 2017, 10, 205. [Google Scholar] [CrossRef] [Green Version]
- Hopken, M.W.; Ryan, B.M.; Huyvaert, K.P.; Piaggio, A.J. Picky eaters are rare: DNA-based blood meal analysis of Culicoides (Diptera: Ceratopogonidae) species from the United States. Parasites Vectors 2017, 10, 169. [Google Scholar] [CrossRef]
- Bellis, G.; Dyce, A.; Gopurenko, D.; Mitchell, A. Revision of the immaculatus group of Culicoides Latreille (Diptera: Ceratopogonidae) from the Australasian region with description of two new species. Zootaxa 2013, 3680, 15–37. [Google Scholar] [CrossRef] [Green Version]
- Dallas, J.F.; Cruickshank, R.; Linton, Y.M.; Nolan, D.; Patakakis, M.; Braverman, Y.; Capela, R.; Capela, M.; Pena, I.; Meiswinkel, R. Phylogenetic status and matrilineal structure of the biting midge, Culicoides imicola, in Portugal, Rhodes and Israel. Med. Vet. Entomol. 2003, 17, 379–387. [Google Scholar] [CrossRef]
- Altschul, S.F.; Gish, W.; Miller, W.; Myers, E.W.; Lipman, D.J. Basic local alignment search tool. J. Mol. Biol. 1990, 215, 403–410. [Google Scholar] [CrossRef]
- Hebert, P.D.N.; Cywinska, A.; Ball, S.L.; deWaard, J.R. Biological identifications through DNA barcodes. Proc. R. Soc. Lond. Ser. 2003, 270, 313–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Riaz, T.; Shehzad, W.; Viari, A.; Pompanon, F.; Taberlet, P.; Coissac, E. ecoPrimers: Inference of new DNA barcode markers from whole genome sequence analysis. Nucleic Acids Res. 2011, 39, e145. [Google Scholar] [CrossRef] [PubMed]
- Townzen, J.S.; Brower, A.V.Z.; Judd, D.D. Identification of mosquito bloodmeals using mitochondrial cytochrome oxidase subunit I and cytochrome b gene sequences. Med. Vet. Entomol. 2008, 22, 386–393. [Google Scholar] [CrossRef] [PubMed]
- Pettersson, E.; Bensch, S.; Ander, M.; Chirico, J.; Sigvald, R.; Ignell, R. Molecular identification of bloodmeals and species composition in Culicoides biting midges. Med. Vet. Entomol. 2013, 27, 104–112. [Google Scholar] [CrossRef]
- Bolger, A.M.; Lohse, M.; Usadel, B. Trimmomatic: A flexible trimmer for Illumina sequence data. Bioinformatics 2014, 30, 2114–2120. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. Search and clustering orders of magnitude faster than BLAST. Bioinformatics 2010, 26, 2460–2461. [Google Scholar] [CrossRef] [Green Version]
- Rognes, T.; Flouri, T.; Nichols, B.; Quince, C.; Mahé, F. VSEARCH: A versatile open source tool for metagenomics. PeerJ 2016, 4, e2584. [Google Scholar] [CrossRef]
- Cristescu, M.E. From barcoding single individuals to metabarcoding biological communities: Towards an integrative approach to the study of global biodiversity. Trends Ecol. Evol. 2014, 29, 566–571. [Google Scholar] [CrossRef]
- Kilpatrick, A.M.; Kramer, L.D.; Jones, M.J.; Marra, P.P.; Daszak, P. West Nile virus epidemics in North America are driven by shifts in mosquito feeding behavior. PLoS Biol. 2006, 4, e82. [Google Scholar] [CrossRef]
- Thiemann, T.C.; Wheeler, S.S.; Barker, C.M.; Reisen, W.K. Mosquito host selection varies seasonally with host availability and mosquito density. PLoS Negl. Trop. Dis. 2011, 5, e1452. [Google Scholar] [CrossRef]
- Mackay, A.J.; Kramer, W.L.; Meece, J.K.; Brumfield, R.T.; Foil, L.D. Host feeding patterns of Culex mosquitoes (Diptera: Culicidae) in East Baton Rouge Parish, Louisiana. J. Med. Entomol. 2014, 47, 238–248. [Google Scholar] [CrossRef]
- Harrigan, R.J.; Thomassen, H.A.; Buermann, W.; Cummings, R.F.; Kahn, M.E.; Smith, T.B. Economic conditions predict prevalence of West Nile virus. PLoS ONE 2010, 5, e15437. [Google Scholar] [CrossRef] [PubMed]
- Dowling, Z.; Ladeau, S.L.; Armbruster, P.; Biehler, D.; Leisnham, P.T. Socioeconomic status affects mosquito (Diptera: Culicidae) larval habitat type availability and infestation level. J. Med. Entomol. 2013, 50, 764–772. [Google Scholar] [CrossRef] [PubMed]
- Branoff, B. Urban mangrove biology and ecology: Emergent patterns and management implications. In Threats to Mangrove Forests: Hazards, Vulnerability, and Management; Makowski, C., Finkl, C.W., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 521–537. [Google Scholar]
- Falcón, W.; Tremblay, R.L. From the cage to the wild: Introductions of Psittaciformes to Puerto Rico. PeerJ 2018, 6, e5669. [Google Scholar] [CrossRef] [Green Version]
- Faraji, A.; Egizi, A.; Fonseca, D.M.; Unlu, I.; Crepeau, T.; Healy, S.P.; Gaugler, R. Comparative host feeding patterns of the Asian tiger mosquito, Aedes albopictus, in urban and suburban northeastern USA and implications for disease transmission. PLoS Negl. Trop. Dis. 2014, 8, e3037. [Google Scholar] [CrossRef] [Green Version]
- Witmer, G.; Constantin, B.; Boyd, F. Feral and introduced carnivores: Issues and challenges. In Proceedings of the 11th Wildlife Damage Management Conference, Traverse, MI, USA, 16–19 May 2005. [Google Scholar]
- Savage, H.M.; Aggarwal, D.; Apperson, C.S.; Katholi, C.R.; Gordon, E.; Hassan, H.K.; Anderson, M.; Charnetzky, D.; McMillen, L.; Unnasch, E.A.; et al. Host choice and West Nile virus infection rates in blood-fed mosquitoes, including members of the Culex pipiens complex, from Memphis and Shelby County, Tennessee, 2002–2003. Vector-Borne Zoonotic Dis. 2007, 7, 365–386. [Google Scholar] [CrossRef] [Green Version]
- Barrera, R.; Amador, M.; Young, G.; Komar, N. Mosquito (Diptera: Culicidae) bloodmeal sources during a period of West Nile virus transmission in Puerto Rico. J. Med. Entomol. 2011, 48, 701–704. [Google Scholar] [CrossRef] [Green Version]
- Fitzpatrick, D.M.; Hattaway, L.M.; Hsueh, A.N.; Ramos-Niño, M.E.; Cheetham, S.M. PCR-based bloodmeal analysis of Aedes aegypti and Culex quinquefasciatus (Diptera: Culicidae) in St. George Parish, Grenada. J. Med. Entomol. 2019, 56, 1170–1175. [Google Scholar] [CrossRef] [Green Version]
- López-Torres, A.L.; Claudio-Hernández, H.J.; Rodríguez-Gómez, C.A.; Longo, A.V.; Joglar, R.L. Green iguanas (Iguana iguana) in Puerto Rico: Is it time for management? Biol. Invasions 2012, 14, 35–45. [Google Scholar] [CrossRef]
- García-Quijano, C.; Carlo, T.; Arce-Nazario, J. Human ecology of a species introduction: Interactions between humans and introduced green iguanas in a Puerto Rican urban estuary. Hum. Organ. 2011, 70, 164–178. [Google Scholar]
- Barrera, R.; Bingham, A.M.; Hassan, H.K.; Amador, M.; Mackay, A.J.; Unnasch, T.R. Vertebrate hosts of Aedes aegypti and Aedes mediovittatus (Diptera: Culicidae) in rural Puerto Rico. J. Med. Entomol. 2012, 49, 917–921. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ponlawat, A.; Harrington, L.C. Blood feeding Patterns of Aedes aegypti and Aedes albopictus in Thailand. J. Med. Entomol. 2005, 42, 844–849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Morens, D.M.; Fauci, A.S. Dengue and hemorrhagic fever: A potential threat to public health in the United States. JAMA 2008, 299, 214–216. [Google Scholar] [CrossRef] [PubMed]
- Sharp, T.M.; Roth, N.M.; Torres, J.; Ryff, K.R.; Rodríguez, N.M.P.; Mercado, C.; Padró, M.d.P.D.; Ramos, M.; Phillips, R.; Lozier, M. Chikungunya cases identified through passive surveillance and household investigations—Puerto Rico, May 5–August 12, 2014. Morb. Mortal. Wkly. Rep. 2014, 63, 1121. [Google Scholar]
- Sharp, T.M.; Hunsperger, E.; Santiago, G.A.; Muñoz-Jordan, J.L.; Santiago, L.M.; Rivera, A.; Rodríguez-Acosta, R.L.; Gonzalez Feliciano, L.; Margolis, H.S.; Tomashek, K.M. Virus-specific differences in rates of disease during the 2010 Dengue epidemic in Puerto Rico. PLoS Negl. Trop. Dis. 2013, 7, e2159. [Google Scholar] [CrossRef] [Green Version]
- Dupuis, A.P.; Marra, P.P.; Reitsma, R.; Jones, M.J.; Louie, K.L.; Kramer, L.D. Serologic evidence for West Nile virus transmission in Puerto Rico and Cuba. Am. J. Trop. Med. Hyg. 2005, 73, 474–476. [Google Scholar] [CrossRef]
- Barrera, R.; Hunsperger, E.; Muñoz-Jordán, J.L.; Amador, M.; Diaz, A.; Smith, J.; Bessoff, K.; Beltran, M.; Vergne, E.; Verduin, M.; et al. First isolation of West Nile virus in the Caribbean. Am. J. Trop. Med. Hyg. 2008, 78, 666–668. [Google Scholar] [CrossRef]
- Torres-Aponte, J.M.; Luce, R.R.; Hunsperger, E.; Muñoz-Jordan, J.L.; Beltrán, M.; Vergne, E.; Argüello, D.F.; García, E.J.; Sun, W.; Tomashek, K.M. Enhanced West Nile virus surveillance in a dengue-endemic area—Puerto Rico, 2007. Am. J. Trop. Med. Hyg. 2013, 88, 997–1002. [Google Scholar] [CrossRef] [Green Version]
- Komar, N.; Bessoff, K.; Diaz, A.; Amador, M.; Young, G.; Seda, R.; Perez, T.; Hunsperger, E. Avian hosts of West Nile virus in Puerto Rico. Vector-Borne Zoonotic Dis. 2011, 12, 47–54. [Google Scholar] [CrossRef]
- Fitzgerald, S.D. Overview of West Nile Virus Infection in Poultry. Available online: https://www.merckvetmanual.com/poultry/west-nile-virus-infection-in-poultry/overview-of-west-nile-virus-infection-in-poultry (accessed on 14 December 2020).
- Ostfeld, R.S.; Keesing, F. Biodiversity series: The function of biodiversity in the ecology of vector-borne zoonotic diseases. Can. J. Zool. 2000, 78, 2061–2078. [Google Scholar] [CrossRef]
- Swaddle, J.P.; Calos, S.E. Increased avian diversity Is associated with lower incidence of human West Nile infection: Observation of the dilution effect. PLoS ONE 2008, 3, e2488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keesing, F.; Holt, R.D.; Ostfeld, R.S. Effects of species diversity on disease risk. Ecol. Lett. 2006, 9, 485–498. [Google Scholar] [CrossRef] [PubMed]
- Rohr, J.R.; Civitello, D.J.; Halliday, F.W.; Hudson, P.J.; Lafferty, K.D.; Wood, C.L.; Mordecai, E.A. Towards common ground in the biodiversity–disease debate. Nat. Ecol. Evol. 2020, 4, 24–33. [Google Scholar] [CrossRef] [PubMed]
- Olden, J.D.; LeRoy Poff, N.; Douglas, M.R.; Douglas, M.E.; Fausch, K.D. Ecological and evolutionary consequences of biotic homogenization. Trends Ecol. Evol. 2004, 19, 18–24. [Google Scholar] [CrossRef] [PubMed]
- Brown, H.E.; Childs, J.E.; Diuk-Wasser, M.A.; Fish, D. Ecologic factors associated with West Nile virus transmission, northeastern United States. Emerg. Infect. Dis. 2008, 14, 1539. [Google Scholar] [CrossRef] [PubMed]
- van Riper III, C.; van Riper, S.G.; Goff, M.L.; Laird, M. The epizootiology and ecological significance of malaria in Hawaiian land birds. Ecol. Monogr. 1986, 56, 327–344. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Variable | CA | MP | PN | RP | TO | VM | VV |
|---|---|---|---|---|---|---|---|
| Number of abandoned homes | 0.769 | 0.769 | 0.461 | 0.080 | 0.538 | 0.308 | 0.308 |
| Number of parks | 0.000 | 0.308 | 0.000 | 0.167 | 0.077 | 0.154 | 0.000 |
| Number of freshwater bodies | 0.231 | 0.385 | 0.000 | 0.000 | 0.000 | 0.153 | 0.000 |
| Number of litter items | 22.5 | 41.5 | 20.8 | 5.1 | 19.4 | 15.3 | 4.1 |
| Human population density per mi² | 1509 | 681 | 629 | 630 | 1433 | 1001 | 641 |
| Proportion unemployment | 0.180 | 0.447 | 0.306 | 0.332 | 0.586 | 0.293 | 0.167 |
| Proportion with college education | 0.227 | 0.164 | 0.386 | 0.687 | 0.204 | 0.519 | 0.614 |
| Proportion below poverty | 0.599 | 0.626 | 0.445 | 0.246 | 0.536 | 0.246 | 0.147 |
| No health insurance | 118 | 114 | 143 | 70 | 256 | 162 | 73 |
| Median household income (USD) | 10,000 | 10,000 | 10,000 | 15,000 | 10,000 | 25,000 | 65,000 |
| Class | Species | Culex quinquefasciatus | Aedes aegypti | ||
|---|---|---|---|---|---|
| n | Frequency | n | Frequency | ||
| Aves | Agapornis fischeri | 1 | 0.002 | ||
| Aves | Butorides virescens | 1 | 0.056 | ||
| Aves | Cairina moschata | 1 | 0.002 | ||
| Aves | Coereba flaveola | 3 | 0.005 | ||
| Aves | Columbidae | 1 | 0.002 | ||
| Aves | Columbina passerina | 1 | 0.002 | ||
| Aves | Gallus gallus | 443 | 0.756 | 1 | 0.056 |
| Aves | Nyctanassa violacea | 5 | 0.009 | ||
| Aves | Nycticorax nycticorax | 1 | 0.002 | ||
| Aves | Nymphicus hollandicus | 1 | 0.002 | ||
| Aves | Passer domesticus | 4 | 0.007 | ||
| Aves | Quiscalus niger | 4 | 0.007 | ||
| Aves | Turdus sp. | 1 | 0.002 | ||
| Aves | Tyrannus dominicensis | 1 | 0.002 | ||
| Aves | Zenaida asiatica | 4 | 0.007 | ||
| Aves | Zenaida aurita | 1 | 0.002 | ||
| Aves | Zenaida macroura | 1 | 0.002 | ||
| Aves | Zenaida sp. | 3 | 0.005 | ||
| Total Aves | 476 | 2 | |||
| Mammalia | Bos taurus | 1 | 0.002 | ||
| Mammalia | Canis lupus familiaris | 60 | 0.102 | 8 | 0.444 |
| Mammalia | Felis catus | 11 | 0.019 | 1 | 0.056 |
| Mammalia | Homo sapiens | 20 | 0.034 | 7 | 0.389 |
| Mammalia | Mus musculus | 8 | 0.014 | ||
| Mammalia | Rattus rattus | 1 | 0.002 | ||
| Mammalia | Sus scrofa | 4 | 0.007 | ||
| Total Mammalia | 105 | 16 | |||
| Reptilia | Iguana sp. | 5 | 0.009 | ||
| Total | 586 | 18 | |||
| Month/Year | Culex quinquefasciatus | Aedes aegypti | ||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Total | Class | n | Frequency | Species | n | Freq | Total | Class | n | Frequency | Species | n | Freq | |
| 01/2018 | 31 | Aves | 24 | 0.77 | Coereba flaveola | 1 | 0.03 | |||||||
| Gallus gallus | 23 | 0.74 | ||||||||||||
| Mammalia | 7 | 0.23 | Canis lupus familiaris | 6 | 0.19 | |||||||||
| Homo sapiens | 1 | 0.03 | ||||||||||||
| 03/2018 | 67 | Aves | 52 | 0.78 | Agapornis fischeri | 1 | 0.01 | 2 | Mammalia | 2 | 1.00 | Canis lupus familiaris | 1 | 0.50 |
| Columbidae | 1 | 0.01 | Homo sapiens | 1 | 0.50 | |||||||||
| Gallus gallus | 49 | 0.73 | ||||||||||||
| Nyctanassa violacea | 1 | 0.01 | ||||||||||||
| Mammalia | 15 | 0.22 | Canis lupus familiaris | 10 | 0.15 | |||||||||
| Felis catus | 1 | 0.01 | ||||||||||||
| Homo sapiens | 4 | 0.06 | ||||||||||||
| 05/2018 | 168 | Aves | 138 | 0.82 | Cairina moschata | 1 | 0.01 | 2 | Mammalia | 2 | 1.00 | Canis lupus familiaris | 2 | 1.00 |
| Coereba flaveola | 2 | 0.01 | ||||||||||||
| Columbina passerina | 1 | 0.01 | ||||||||||||
| Gallus gallus | 128 | 0.76 | ||||||||||||
| Nyctanassa violacea | 1 | 0.01 | ||||||||||||
| Passer domesticus | 1 | 0.01 | ||||||||||||
| Quiscalus niger | 3 | 0.02 | ||||||||||||
| Zenaida asiatica | 1 | 0.01 | ||||||||||||
| Mammalia | 30 | 0.18 | Bos taurus | 1 | 0.01 | |||||||||
| Canis lupus familiaris | 13 | 0.08 | ||||||||||||
| Felis catus | 6 | 0.04 | ||||||||||||
| Homo sapiens | 4 | 0.02 | ||||||||||||
| Mus musculus | 5 | 0.03 | ||||||||||||
| Sus scrofa | 1 | 0.01 | ||||||||||||
| 10/2018 | 73 | Aves | 61 | 0.84 | Gallus gallus | 59 | 0.81 | 6 | Aves | 1 | 0.17 | Butorides virescens | 1 | 0.17 |
| Passer domesticus | 1 | 0.01 | Mammalia | 5 | 0.83 | Canis lupus familiaris | 2 | 0.33 | ||||||
| Tyrannus dominicensis | 1 | 0.01 | Homo sapiens | 3 | 0.50 | |||||||||
| Mammalia | 12 | 0.16 | Canis lupus familiaris | 5 | 0.07 | |||||||||
| Felis catus | 1 | 0.01 | ||||||||||||
| Homo sapiens | 4 | 0.05 | ||||||||||||
| Sus scrofa | 2 | 0.03 | ||||||||||||
| 01/2019 | 103 | Aves | 90 | 0.87 | Gallus gallus | 87 | 0.84 | 2 | Mammalia | 2 | 1.00 | Homo sapiens | 2 | 1.00 |
| Nyctanassa violacea | 1 | 0.01 | ||||||||||||
| Passer domesticus | 1 | 0.01 | ||||||||||||
| Zenaida aurita | 1 | 0.01 | ||||||||||||
| Mammalia | 11 | 0.11 | Canis lupus familiaris | 11 | 0.11 | |||||||||
| Reptilia | 2 | 0.02 | Iguana sp. | 2 | 0.02 | |||||||||
| 05/2019 | 144 | Aves | 111 | 0.77 | Gallus gallus | 97 | 0.67 | 6 | Aves | 1 | 0.17 | Canis lupus familiaris | 3 | 0.50 |
| Nyctanassa violacea | 2 | 0.01 | Mammalia | 5 | 0.83 | Felis catus | 1 | 0.17 | ||||||
| Nycticorax nycticorax | 1 | 0.01 | Gallus gallus | 1 | 0.17 | |||||||||
| Nymphicus hollandicus | 1 | 0.01 | Homo sapiens | 1 | 0.17 | |||||||||
| Passer domesticus | 1 | 0.01 | ||||||||||||
| Quiscalus niger | 1 | 0.01 | ||||||||||||
| Turdus sp | 1 | 0.01 | ||||||||||||
| Zenaida asiatica | 3 | 0.02 | ||||||||||||
| Zenaida macroura | 1 | 0.01 | ||||||||||||
| Zenaida sp. | 3 | 0.02 | ||||||||||||
| Mammalia | 30 | 0.21 | Canis lupus familiaris | 15 | 0.10 | |||||||||
| Felis catus | 3 | 0.02 | ||||||||||||
| Mus musculus | 3 | 0.02 | ||||||||||||
| Rattus rattus | 1 | 0.01 | ||||||||||||
| Sus scrofa | 1 | 0.01 | ||||||||||||
| Homo sapiens | 7 | 0.05 | ||||||||||||
| Reptilia | 3 | 0.02 | Iguana sp. | 3 | 0.02 | |||||||||
| Culex quinquefasciatus Blood Meal Identification | Aedes aegypti Blood Meal Identification | ||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Site | Total n | Class | n | Frequency | Species | n | Frequency | Site | Total n | Class | n | Frequency | Species | n | Frequency |
| C | 1 | Aves | 1 | 1.000 | Coereba flaveola | 1 | 1.000 | C | 2 | Mammalia | 2 | 1.000 | Homo sapiens | 2 | 1.00 |
| CA | 237 | Aves | 195 | 0.823 | Coereba flaveola | 2 | 0.008 | CA | 3 | Mammalia | 3 | 1.000 | Homo sapiens | 3 | 1.00 |
| Columbina passerina | 1 | 0.004 | |||||||||||||
| Gallus gallus | 189 | 0.797 | |||||||||||||
| Passer domesticus | 2 | 0.008 | |||||||||||||
| Tyrannus dominicensis | 1 | 0.004 | |||||||||||||
| Mammalia | 38 | 0.160 | Bos taurus | 1 | 0.004 | ||||||||||
| Canis lupus familiaris | 19 | 0.080 | |||||||||||||
| Homo sapiens | 9 | 0.038 | |||||||||||||
| Mus musculus | 6 | 0.025 | |||||||||||||
| Sus scrofa | 3 | 0.013 | |||||||||||||
| Reptilia | 4 | 0.017 | Iguana sp. | 4 | 0.017 | ||||||||||
| MP | 32 | Aves | 32 | 1.000 | Gallus gallus | 30 | 0.938 | MP | 2 | Mammalia | 2 | 1.000 | Canis lupus familiaris | 1 | 0.50 |
| Nycticorax nycticorax | 1 | 0.031 | Homo sapiens | 1 | 0.50 | ||||||||||
| Turdus sp. | 1 | 0.031 | |||||||||||||
| PN | 6 | Aves | 3 | 0.500 | Columbidae | 1 | 0.167 | ||||||||
| Gallus gallus | 1 | 0.167 | |||||||||||||
| Passer domesticus | 1 | 0.167 | |||||||||||||
| Mammalia | 3 | 0.500 | Canis lupus familiaris | 1 | 0.167 | ||||||||||
| Homo sapiens | 2 | 0.333 | |||||||||||||
| RP | 4 | Aves | 2 | 0.500 | Gallus gallus | 1 | 0.250 | RP | 4 | Mammalia | 4 | 1.000 | Canis lupus familiaris | 3 | 0.75 |
| Nymphicus hollandicus | 1 | 0.250 | Felis catus | 1 | 0.25 | ||||||||||
| Mammalia | 2 | 0.500 | Canis lupus familiaris | 2 | 0.500 | ||||||||||
| T | 296 | Aves | 237 | 0.801 | Cairina moschata | 1 | 0.003 | T | 3 | Aves | 1 | 0.333 | Gallus gallus | 1 | 0.33 |
| Gallus gallus | 218 | 0.736 | Mammalia | 2 | 0.667 | Canis lupus familiaris | 1 | 0.33 | |||||||
| Nyctanassa violacea | 5 | 0.017 | Homo sapiens | 1 | 0.33 | ||||||||||
| Passer domesticus | 1 | 0.003 | |||||||||||||
| Quiscalus niger | 4 | 0.014 | |||||||||||||
| Zenadia asiatica | 4 | 0.014 | |||||||||||||
| Zenaida macroura | 1 | 0.003 | |||||||||||||
| Zenaida sp. | 3 | 0.010 | |||||||||||||
| Mammalia | 58 | 0.196 | Canis lupus familiaris | 34 | 0.115 | ||||||||||
| Felis catus | 11 | 0.037 | |||||||||||||
| Homo sapiens | 9 | 0.030 | |||||||||||||
| Mus musculus | 2 | 0.007 | |||||||||||||
| Rattus rattus | 1 | 0.003 | |||||||||||||
| Sus scrofa | 1 | 0.003 | |||||||||||||
| Reptilia | 1 | 0.003 | Iguana sp. | 1 | 0.003 | ||||||||||
| VM/VV | 10 | Aves | 6 | 0.600 | Agapornis fischeri | 1 | 0.100 | V | 1 | Aves | 1 | 1.000 | Butorides virescens | 1 | 1.00 |
| Gallus gallus | 4 | 0.400 | 3 | Mammalia | 3 | 1.000 | Canis lupus familiaris | 1 | 1.00 | ||||||
| Zenaida aurita | 1 | 0.100 | |||||||||||||
| Mammalia | 4 | 0.400 | Canis lupus familiaris | 4 | 0.400 | ||||||||||
| Date | H′ | D | SR |
|---|---|---|---|
| 01/2018 | 0.76 | 0.59 | 4.00 |
| 03/2018 | 1.05 | 0.47 | 4.34 |
| 05/2018 | 1.06 | 0.53 | 5.94 |
| 10/2018 | 0.76 | 0.62 | 4.39 |
| 01/2019 | 0.85 | 0.58 | 3.40 |
| 05/2019 | 0.88 | 0.61 | 7.08 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hopken, M.W.; Reyes-Torres, L.J.; Scavo, N.; Piaggio, A.J.; Abdo, Z.; Taylor, D.; Pierce, J.; Yee, D.A. Temporal and Spatial Blood Feeding Patterns of Urban Mosquitoes in the San Juan Metropolitan Area, Puerto Rico. Insects 2021, 12, 129. https://doi.org/10.3390/insects12020129
Hopken MW, Reyes-Torres LJ, Scavo N, Piaggio AJ, Abdo Z, Taylor D, Pierce J, Yee DA. Temporal and Spatial Blood Feeding Patterns of Urban Mosquitoes in the San Juan Metropolitan Area, Puerto Rico. Insects. 2021; 12(2):129. https://doi.org/10.3390/insects12020129
Chicago/Turabian StyleHopken, Matthew W., Limarie J. Reyes-Torres, Nicole Scavo, Antoinette J. Piaggio, Zaid Abdo, Daniel Taylor, James Pierce, and Donald A. Yee. 2021. "Temporal and Spatial Blood Feeding Patterns of Urban Mosquitoes in the San Juan Metropolitan Area, Puerto Rico" Insects 12, no. 2: 129. https://doi.org/10.3390/insects12020129