
Supporting Bees in Cities: How Bees Are Influenced by Local and Landscape Features


Abstract
:Simple Summary
Abstract
1. Introduction
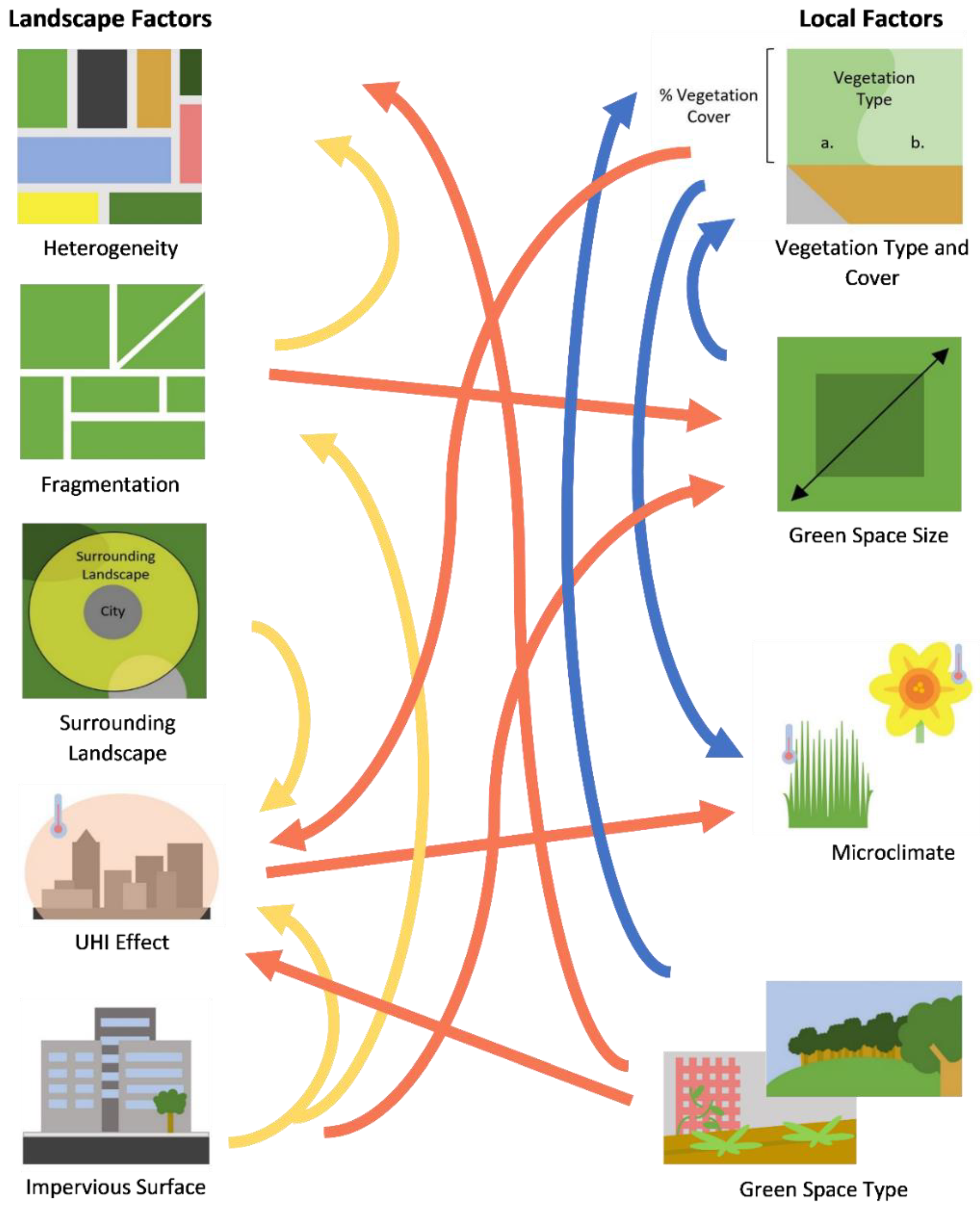
2. Landscape and Local Feature Influences on Pollinators
3. Landscape Features
3.1. Habitat Loss, Fragmentation and Heterogeneity
3.2. Urban Heat Island Effect
3.3. Surrounding Landscape and Impervious Surface
4. Local Features
4.1. Microclimate
4.2. Urban Vegetation
4.3. Green Space Size
4.4. Green Space/Habitat Type
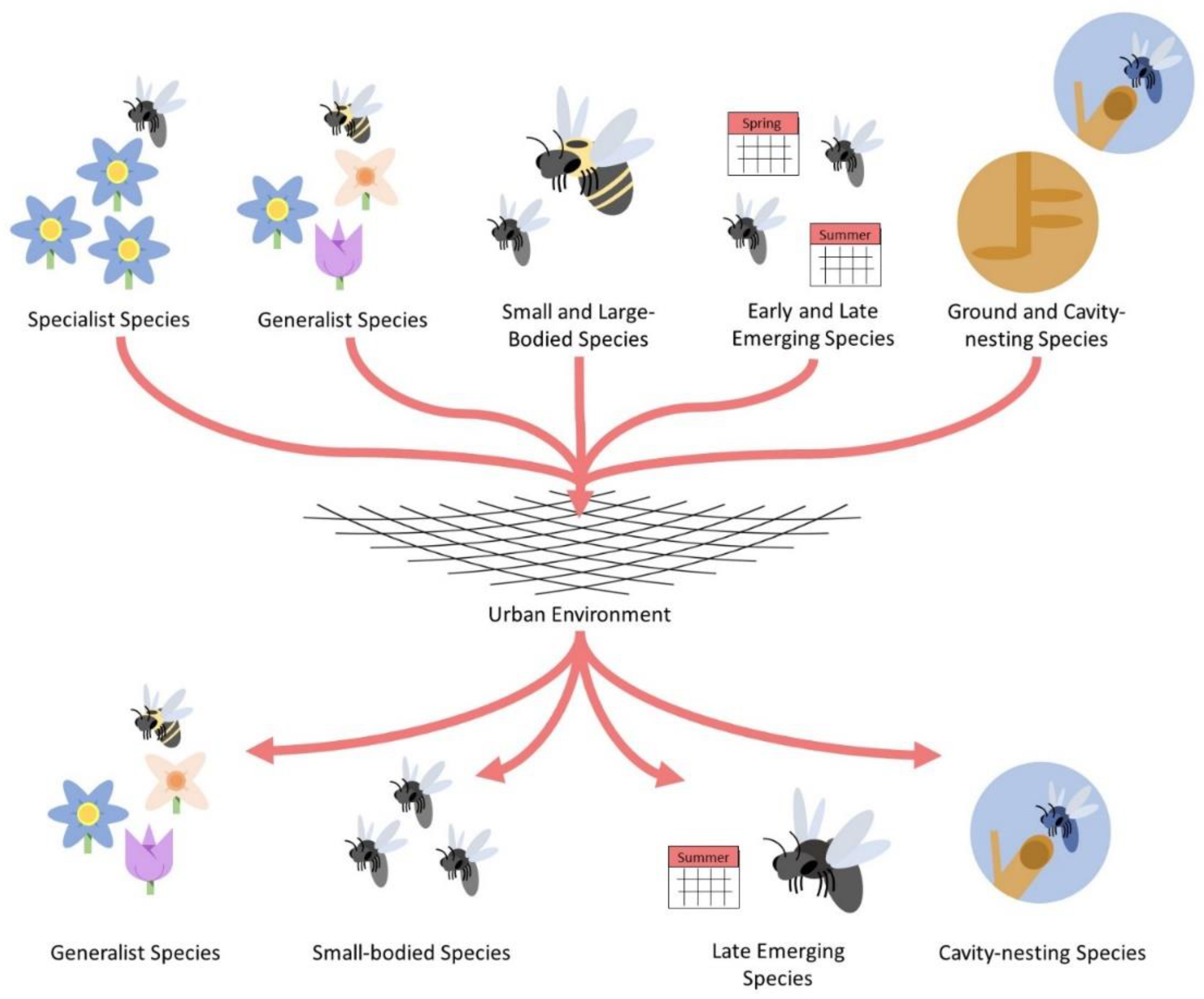
5. Functional Traits Affected by Urbanization
5.1. Dietary Breadth
5.2. Nesting Strategy
5.3. Body Size
5.4. Behavior
5.5. Phylogenetic Diversity

{kind=link}
{kind=link}
| Functional Trait | Favored Strategy | Urban Factors Affecting Traits | References |
|---|---|---|---|
| Body size | Inconclusive (mixed results) | UHI effect, Habitat fragmentation, Vegetation cover and type | [71,121,123,125] |
| Diet strategy | Polylecty | Green space type, Vegetation type and cover | [11,18,70,84,103] |
| Nesting strategy | Cavity nesting | Impervious service, Green space type, Green space size | [26,70,71,77] |
| Phenology | Late emerging | UHI effect, Vegetation type and cover | [15,18,129] |
| Sociality | Inconclusive (mixed results) | Impervious surface, Vegetation cover and type | [15,18,22,111,128] |
| Phylogenetic Diversity | Groups such as Lasioglossum (Dialictus), in some instances Bombus | Impervious surface, Vegetation cover and type, UHI effect, Fragmentation, Green space size | [103,130,131,134,135] |
6. Conservation Aims and Future Directions
7. Conclusions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Wu, J. Urban Ecology and Sustainability: The State-of-the-Science and Future Directions. Landsc. Urban Plan. 2014, 125, 209–221. [Google Scholar] [CrossRef]
- Cardoso, M.C.; Gonçalves, R.B. Reduction by Half: The Impact on Bees of 34 Years of Urbanization. Urban Ecosyst. 2018, 21, 943–949. [Google Scholar] [CrossRef]
- Fenoglio, M.S.; Rossetti, M.R.; Videla, M. Negative Effects of Urbanization on Terrestrial Arthropod Communities: A Meta-Analysis. Glob. Ecol. Biogeogr. 2020, 29, 1412–1429. [Google Scholar] [CrossRef]
- Kurylo, J.S.; Threlfall, C.G.; Parris, K.M.; Ossola, A.; Williams, N.S.G.; Evans, K.L. Butterfly Richness and Abundance along a Gradient of Imperviousness and the Importance of Matrix Quality. Ecol. Appl. 2020, 30. [Google Scholar] [CrossRef] [PubMed]
- Piano, E.; Souffreau, C.; Merckx, T.; Baardsen, L.F.; Backeljau, T.; Bonte, D.; Brans, K.I.; Cours, M.; Dahirel, M.; Debortoli, N.; et al. Urbanization Drives cross-Taxon Declines in Abundance and Diversity at Multiple Spatial Scales. Glob. Chang. Biol. 2019, 26, 1196–1211. [Google Scholar] [CrossRef] [PubMed]
- Sattler, T.; Duelli, P.; Obrist, M.K.; Arlettaz, R.; Moretti, M. Response of Arthropod Species Richness And functional Groups to Urban Habitat Structure and Management. Landsc. Ecol. 2010, 25, 941–954. [Google Scholar] [CrossRef] [Green Version]
- Harrison, T.; Winfree, R. Urban Drivers of Plant-Pollinator Interactions. Funct. Ecol. 2015, 29, 879–888. [Google Scholar] [CrossRef]
- Ballare, K.M.; Neff, J.L.; Ruppel, R.; Jha, S. Multi-Scalar Drivers of Biodiversity: Local Management Mediates Wild Bee Community Response To Regional Urbanization. Ecol. Appl. 2019, 29, e01869. [Google Scholar] [CrossRef]
- Beninde, J.; Veith, M.; Hochkirch, A. Biodiversity in Cities Needs Space: A Meta-Analysis of Factors Determining Intra-Urban Biodiversity Variation. Ecol. Lett. 2015, 18, 581–592. [Google Scholar] [CrossRef]
- Egerer, M.; Arel, C.; Otoshi, M.D.; Quistberg, R.D.; Bichier, P.; Philpott, S.M. Urban Arthropods Respond Variably to Changes in Landscape Context and Spatial Scale. J. Urban Ecol. 2017, 3. [Google Scholar] [CrossRef] [Green Version]
- Geslin, B.; Le Féon, V.; Folschweiller, M.; Flacher, F.; Carmignac, D.; Motard, E.; Perret, S.; Dajoz, I. The Proportion of Impervious Surfaces At the Landscape Scale Structures Wild Bee Assemblages in a Densely Populated Region. Ecol. Evol. 2016, 6, 6599–6615. [Google Scholar] [CrossRef] [PubMed]
- Quistberg, R.D.; Bichier, P.; Philpott, S.M. Landscape and Local Correlates of Bee Abundance and Species Richness in Urban Gardens. Environ. Èntomol. 2016, 45, 592–601. [Google Scholar] [CrossRef] [PubMed]
- Dale, A.G.; Frank, S.D. Urban plants and climate drive unique arthropod interactions with unpredictable consequences. Curr. Opin. Insect Sci. 2018, 29, 27–33. [Google Scholar] [CrossRef] [PubMed]
- Williams, N.S.; Schwartz, M.W.; Vesk, P.A.; McCarthy, M.A.; Hahs, A.K.; Clemants, S.E.; Corlett, R.T.; Duncan, R.P.; Norton, B.A.; Thompson, K.; et al. A Conceptual Framework for Predicting the Effects of Urban Environments on Floras. J. Ecol. 2009, 97, 4–9. [Google Scholar] [CrossRef]
- Wenzel, A.; Grass, I.; Belavadi, V.V.; Tscharntke, T. How Urbanization is Driving Pollinator Diversity and Pollination–A Systematic Review. Biol. Conserv. 2020, 241, 108321. [Google Scholar] [CrossRef]
- Deguines, N.; Julliard, R.; De Flores, M.; Fontaine, C. Functional Homogenization of Flower Visitor Communities with Urbanization. Ecol. Evol. 2016, 6, 1967–1976. [Google Scholar] [CrossRef] [Green Version]
- Knop, E. Biotic Homogenization of Three Insect Groups Due to Urbanization. Glob. Chang. Biol. 2016, 22, 228–236. [Google Scholar] [CrossRef]
- Banaszak-Cibicka, W.; Żmihorski, M. Wild Bees along an Urban Gradient: Winners and Losers. J. Insect Conserv. 2012, 16, 331–343. [Google Scholar] [CrossRef] [Green Version]
- Hamblin, A.L.; Youngsteadt, E.; López-Uribe, M.M.; Frank, S.D. Physiological Thermal Limits Predict Differential Responses of Bees to Urban Heat-Island Effects. Biol. Lett. 2017, 13, 20170125. [Google Scholar] [CrossRef]
- Nagy, D.D.; Magura, T.; Horváth, R.; Debnár, Z.; Tóthmérész, B. Arthropod Assemblages and Functional Responses along an Urbanization Gradient: A Trait-Based Multi-Taxa Approach. Urban For. Urban Green. 2018, 30, 157–168. [Google Scholar] [CrossRef]
- Baldock, K.C.R.; Goddard, M.A.; Hicks, D.M.; Kunin, W.E.; Mitschunas, N.; Osgathorpe, L.M.; Potts, S.G.; Robertson, K.M.; Scott, A.V.; Stone, G.N.; et al. Where is the UK’s Pollinator Biodiversity? The Importance of Urban Areas for Flower-Visiting Insects. Proc. R. Soc. B Boil. Sci. 2015, 282, 20142849. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banaszak-Cibicka, W.; Twerd, L.; Fliszkiewicz, M.; Giejdasz, K.; Langowska, A. City Parks vs. Natural Areas-is it Possible to Preserve a Natural Level of Bee Richness and Abundance in a City Park? Urban Ecosyst. 2018, 21, 599–613. [Google Scholar] [CrossRef] [Green Version]
- Kaluza, B.F.; Wallace, H.; Heard, T.A.; Klein, A.; Leonhardt, S.D. Urban Gardens Promote Bee Foraging over Natural Habitats and Plantations. Ecol. Evol. 2016, 6, 1304–1316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sirohi, M.H.; Jackson, J.; Edwards, M.; Ollerton, J. Diversity and Abundance of Solitary and Primitively Eusocial Bees in an Urban Centre: A Case Study from NORTHAMPTON (England). J. Insect Conserv. 2015, 19, 487–500. [Google Scholar] [CrossRef] [Green Version]
- Tommasi, D.; Miro, A.; Higo, H.A.; Winston, M.L. Bee Diversity and Abundance in an Urban Setting. Can. Èntomol. 2004, 136, 851–869. [Google Scholar] [CrossRef]
- Baldock, K.C.R.; Goddard, M.A.; Hicks, D.M.; Kunin, W.E.; Mitschunas, N.; Morse, H.; Osgathorpe, L.M.; Potts, S.G.; Robertson, K.M.; Scott, A.V.; et al. A Systems Approach Reveals Urban Pollinator Hotspots and Conservation Opportunities. Nat. Ecol. Evol. 2019, 3, 363–373. [Google Scholar] [CrossRef] [Green Version]
- Dylewski, Ł.; Maćkowiak, Ł.; Banaszak-Cibicka, W. Linking pollinators and city flora: How Vegetation Composition and Environmental Features Shapes Pollinators Composition in Urban Environment. Urban For. Urban Green. 2020, 56, 126795. [Google Scholar] [CrossRef]
- Braaker, S.; Obrist, M.K.; Ghazoul, J.; Moretti, M. Habitat Connectivity and Local Conditions Shape Taxonomic and Functional Diversity of Arthropods on Green Roofs. J. Anim. Ecol. 2017, 86, 521–531. [Google Scholar] [CrossRef] [Green Version]
- United Nations. 68% of the World Population Projected to Live in Urban Areas by 2050, Says UN. UN DESA. United Nations Department of Economic and Social Affairs. 2018. Available online: www.un.org (accessed on 15 September 2020).
- Government of Canada, Statistics Canada. Canada’s Population Estimates: Subprovincial Areas, 1 July 2018. Statcan.gc.ca; 2019. Available online: https://www150.statcan.gc.ca/n1/daily-quotidien/190328/dq190328b-eng.htm (accessed on 27 November 2020).
- Center for Sustainable Systems, University of Michigan. U.S. Cities Factsheet; Center for Sustainable Systems, University of Michigan: Ann Arbor, MI, USA, 2020. [Google Scholar]
- Jones, E.L.; Leather, S.R. Invertebrates in Urban Areas: A Review. Eur. J. Èntomol. 2012, 109, 463–478. [Google Scholar] [CrossRef] [Green Version]
- Potts, S.G.; Biesmeijer, J.C.; Kremen, C.; Neumann, P.; Schweiger, O.; Kunin, W.E. Global Pollinator Declines: Trends, Impacts and Drivers. Trends Ecol. Evol. 2010, 25, 345–353. [Google Scholar] [CrossRef]
- Ollerton, J.; Winfree, R.; Tarrant, S. How Many Flowering Plants are Pollinated by Animals? Oikos 2011, 120, 321–326. [Google Scholar] [CrossRef]
- Kearns, C.A.; Inouye, D.W.; Waser, N.M. ENDANGERED MUTUALISMS: The Conservation of Plant-Pollinator Interactions. Annu. Rev. Ecol. Syst. 1998, 29, 83–112. [Google Scholar] [CrossRef]
- Lovell, S.T. Multifunctional Urban Agriculture for Sustainable Land Use Planning in the United States. Sustainability 2010, 2, 2499–2522. [Google Scholar] [CrossRef] [Green Version]
- Siegner, A.B.; Sowerwine, J.; Acey, C. Does Urban Agriculture Improve Food Security? Examining the Nexus of Food Access and Distribution of Urban Produced Foods in the United States: A Systematic Review. Sustainability 2018, 10, 2988. [Google Scholar] [CrossRef] [Green Version]
- Garibaldi, L.A.; Steffan-Dewenter, I.; Winfree, R.; Aizen, M.A.; Bommarco, R.; Cunningham, S.A.; Kremen, C.; Carvalheiro, L.G.; Harder, L.D.; Afik, O.; et al. Wild Pollinators Enhance Fruit Set of Crops Regardless of Honey Bee Abundance. Science 2013, 339, 1608–1611. [Google Scholar] [CrossRef]
- Lowenstein, D.M.; Matteson, K.C.; Minor, E. Diversity of Wild Bees Supports Pollination Services in an Urbanized Landscape. Oecologia 2015, 179, 811–821. [Google Scholar] [CrossRef]
- Southon, G.E.; Jorgensen, A.; Dunnett, N.; Hoyle, H.; Evans, K.L. Biodiverse Perennial Meadows Have Aesthetic Value and Increase Residents’ Perceptions of Site Quality in Urban Green-Space. Landsc. Urban Plan. 2017, 158, 105–118. [Google Scholar] [CrossRef] [Green Version]
- Pawelek, J.C.; Frankie, G.W.; Thorp, R.W.; Przybylski, M. Modification of a Community GArden to Attract Native Bee Pollinators in Urban San Luis Obispo, California. Cities Environ. 2009, 2, 1–20. [Google Scholar] [CrossRef]
- Pardee, G.L.; Philpott, S.M. Native Plants Are the Bee’s Knees: Local and Landscape Predictors of Bee Richness and Abundance in Backyard Gardens. Urban Ecosyst. 2014, 17, 641–659. [Google Scholar] [CrossRef] [Green Version]
- Kennedy, C.M.; Lonsdorf, E.; Neel, M.C.; Williams, N.M.; Ricketts, T.H.; Winfree, R.; Bommarco, R.; Brittain, C.; Burley, A.L.; Cariveau, D.; et al. A global Quantitative Synthesis of Local and Landscape Effects on wild Bee Pollinators in Agroecosystems. Ecol. Lett. 2013, 16, 584–599. [Google Scholar] [CrossRef]
- McKinney, M.L. Effects of urbanization on species richness: A review of plants and animals. Urban Ecosyst. 2008, 11, 161–176. [Google Scholar] [CrossRef]
- Matteson, K.C.; Langellotto, G.A. Determinates of Inner City Butterfly and Bee Species Richness. Urban Ecosyst. 2010, 13, 333–347. [Google Scholar] [CrossRef] [Green Version]
- Williams, N.M.; Winfree, R. Local habitat characteristics but not landscape urbanization drive pollinator visitation and native plant pollination in forest remnants. Biol. Conserv. 2013, 160, 10–18. [Google Scholar] [CrossRef]
- Viana, B.F. How Well do we Understand Landscape Effects on Pollinators and Pollination Services? J. Pollinat. Ecol. 2012, 7, 31–40. [Google Scholar] [CrossRef]
- Williams, N.M.; Kremen, C. Resource Distributions among Habitats Determine solitary bee Offspring Production in a Mosaic Landscape. Ecol. Appl. 2007, 17, 910–921. [Google Scholar] [CrossRef]
- Winfree, R.; Aguilar, R.; Vázquez, D.P.; Lebuhn, G.; Aizen, M.A. A Meta-Analysis of Bees’ Responses to Anthropogenic Disturbance. Ecology 2009, 90, 2068–2076. [Google Scholar] [CrossRef]
- Stenhouse, R.N. Fragmentation and internal disturbance of native vegetation reserves in the Perth metropolitan area, Western Australia. Landsc. Urban Plan. 2004, 68, 389–401. [Google Scholar] [CrossRef]
- Hadley, A.S.; Betts, M.G. The Effects of Landscape Fragmentation on Pollination Dynamics: Absence of Evidence Not Evidence of Absence. Biol. Rev. 2011, 87, 526–544. [Google Scholar] [CrossRef]
- Greenleaf, S.S.; Williams, N.M.; Winfree, R.; Kremen, C. Bee Foraging Ranges and Their Relationship to Body Size. Oecologia 2007, 153, 589–596. [Google Scholar] [CrossRef]
- Hennig, E.I.; Ghazoul, J. Pollinating Animals in the Urban Environment. Urban Ecosyst. 2012, 15, 149–166. [Google Scholar] [CrossRef] [Green Version]
- Oke, T.R. City size and the urban heat island. Atmos. Environ. 1973, 7, 769–779. [Google Scholar] [CrossRef]
- Kłysik, K.; Fortuniak, K. Temporal and Spatial Characteristics of the Urban Heat Island of Łódź, Poland. Atmos. Environ. 1999, 33, 3885–3895. [Google Scholar] [CrossRef]
- Polidori, C.; García-Gila, J.; Blasco-Aróstegui, J.; Gil-Tapetado, D. Urban areas are favouring the spread of an alien mud-dauber wasp into climatically non-optimal latitudes. Acta Oecologica 2021, 110, 103678. [Google Scholar] [CrossRef]
- Fitch, G.; Wilson, C.J.; Glaum, P.; Vaidya, C.; Simao, M.-C.; Jamieson, M.A. Does urbanization favour exotic bee species? Implications for the conservation of native bees in cities. Biol. Lett. 2019, 15, 20190574. [Google Scholar] [CrossRef] [PubMed]
- Wilson, C.J.; Jamieson, M.A. The Effects of Urbanization on Bee Communities Depends on Floral Resource Availability and Bee Functional Traits. PLoS ONE 2019, 14, e0225852. [Google Scholar] [CrossRef] [Green Version]
- McIntyre, N.; Rango, J.; Fagan, W.; Faeth, S. Ground Arthropod Community Structure in a Heterogeneous Urban Environment. Landsc. Urban Plan. 2001, 52, 257–274. [Google Scholar] [CrossRef]
- Kaiser, A.; Merckx, T.; Van Dyck, H. The Urban Heat Island and Its Spatial Scale Dependent Impact on Survival and Development in Butterflies of Different Thermal Sensitivity. Ecol. Evol. 2016, 6, 4129–4140. [Google Scholar] [CrossRef] [Green Version]
- Burdine, J.D.; McCluney, K.E. Differential Sensitivity of Bees to Urbanization-Driven Changes in Body Temperature and Water Content. Sci. Rep. 2019, 9, 1–10. [Google Scholar] [CrossRef]
- Hamblin, A.L.; Youngsteadt, E.; Frank, S.D. Wild Bee Abundance Declines with Urban Warming, Regardless of Floral Density. Urban Ecosyst. 2018, 21, 419–428. [Google Scholar] [CrossRef]
- Bowler, D.E.; Buyung-Ali, L.; Knight, T.M.; Pullin, A.S. Urban greening to cool towns and cities: A systematic review of the empirical evidence. Landsc. Urban Plan. 2010, 97, 147–155. [Google Scholar] [CrossRef]
- Chang, C.-R.; Li, M.-H.; Chang, S.-D. A preliminary study on the local cool-island intensity of Taipei city parks. Landsc. Urban Plan. 2007, 80, 386–395. [Google Scholar] [CrossRef]
- Gonçalves, A.; Ornellas, G.; Ribeiro, A.C.; Maia, F.; Rocha, A.; Feliciano, M. Urban Cold and Heat Island in the City of Bragança (Portugal). Climate 2018, 6, 70. [Google Scholar] [CrossRef] [Green Version]
- Klein, A.-M.; Brittain, C.; Hendrix, S.D.; Thorp, R.; Williams, N.M.; Kremen, C. Wild Pollination Services to California Almond Rely on Semi-Natural Habitat: Wild Pollination Services to California Almond. J. Appl. Ecol. 2012, 49, 723–732. [Google Scholar] [CrossRef]
- Ricketts, T.H.; Regetz, J.; Steffan-Dewenter, I.; Cunningham, S.A.; Kremen, C.; Bogdanski, A.; Gemmill-Herren, B.; Greenleaf, S.S.; Klein, A.M.; Mayfield, M.M.; et al. Landscape Effects on Crop Pollination Services: Are There General Patterns? Ecol. Lett. 2008, 11, 499–515. [Google Scholar] [CrossRef] [PubMed]
- Tonietto, R.; Fant, J.; Ascher, J.; Ellis, K.; Larkin, D. A Comparison of Bee Communities of Chicago Green Roofs, Parks and Prairies. Landsc. Urban Plan. 2011, 103, 102–108. [Google Scholar] [CrossRef]
- Phillips, B.W.; Gardiner, M.M. Use of Video Surveillance to Measure the Influences of Habitat Management and Landscape Composition on Pollinator Visitation and Pollen Deposition in Pumpkin (Cucurbita pepo) Agroecosystems. PeerJ 2015, 3, e1342. [Google Scholar] [CrossRef] [Green Version]
- Bogusch, P.; Hlaváčková, L.; Gasol, N.R.; Heneberg, P. Near-natural habitats near almond orchards with presence of empty gastropod shells are important for solitary shell-nesting bees and wasps. Agric. Ecosyst. Environ. 2020, 299, 106949. [Google Scholar] [CrossRef]
- Senapathi, D.; Goddard, M.A.; Kunin, W.E.; Baldock, K.C.R. Landscape Impacts on Pollinator Communities in Temperate Systems: Evidence and Knowledge Gaps. Funct. Ecol. 2017, 31, 26–37. [Google Scholar] [CrossRef] [Green Version]
- Kuussaari, M.; Toivonen, M.; Heliölä, J.; Pöyry, J.; Mellado, J.; Ekroos, J.; Hyyryläinen, V.; Vähä-Piikkiö, I.; Tiainen, J. Butterfly Species’ Responses to Urbanization: Differing Effects of Human Population Density and Built-Up Area. Urban Ecosyst. 2020. [Google Scholar] [CrossRef]
- Cane, J.H.; Minckley, R.L.; Kervin, L.J.; Roulston, T.H.; Williams, N.M. Complex Responses within A Desert Bee Guild (Hymenoptera: Apiformes) To Urban Habitat Fragmentation. Ecol. Appl. 2006, 16, 632–644. [Google Scholar] [CrossRef] [Green Version]
- Pincebourde, S.; Murdock, C.C.; Vickers, M.; Sears, M.W. Fine-Scale Microclimatic Variation Can Shape the Responses of Organisms to Global Change in Both Natural and Urban Environments. Integr. Comp. Biol. 2016, 56, 45–61. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fisogni, A.; Hautekèete, N.; Piquot, Y.; Brun, M.; Vanappelghem, C.; Michez, D.; Massol, F. Urbanization drives an early spring for plants but not for pollinators. Oikos 2020, 129, 1681–1691. [Google Scholar] [CrossRef]
- Crum, S.M.; Jenerette, G.D. Microclimate Variation among Urban Land Covers: The Importance of Vertical and Horizontal Structure in Air and Land Surface Temperature Relationships. J. Appl. Meteorol. Clim. 2017, 56, 2531–2543. [Google Scholar] [CrossRef]
- Herrera, C.M. Floral Biology, Microclimate, and Pollination by Ectothermic Bees in an Early-Blooming Herb. Ecology 1995, 76, 218–228. [Google Scholar] [CrossRef]
- Richards, M.H.; Garate, A.C.; Shehata, M.; Groom, D.; Tattersall, G.J.; Welch, K.C. Effect of Nest Microclimate Temperatures on Metabolic Rates of Small Carpenter Bees, Ceratina calcarata (Hymenoptera: Apidae). Can. Èntomol. 2020, 152, 772–782. [Google Scholar] [CrossRef]
- Radmacher, S.; Strohm, E. Factors Affecting Offspring Body Size in the Solitary Bee Osmia bicornis (Hymenoptera, Megachilidae). Apidologie 2009, 41, 169–177. [Google Scholar] [CrossRef] [Green Version]
- WallisDeVries, M.F.; Van Swaay, C.A.M. Global Warming and Excess Nitrogen May Induce Butterfly Decline by Microclimatic Cooling: Microclimate Cooling and Butterfly Decline. Glob. Chang. Biol. 2006, 12, 1620–1626. [Google Scholar] [CrossRef]
- Threlfall, C.G.; Ossola, A.; Hahs, A.K.; Williams, N.S.G.; Wilson, L.; Livesley, S.J. Variation in Vegetation Structure and Composition across Urban Green Space Types. Front. Ecol. Evol. 2016, 4. [Google Scholar] [CrossRef] [Green Version]
- Leston, L.; Koper, N. Urban rights-of-way as extensive butterfly habitats: A Case Study from Winnipeg, Canada. Landsc. Urban Plan. 2017, 157, 56–62. [Google Scholar] [CrossRef]
- Martins, K.T.; Gonzalez, A.; Lechowicz, M.J. Patterns of Pollinator Turnover and Increasing Diversity Associated with Urban Habitats. Urban Ecosyst. 2017, 20, 1359–1371. [Google Scholar] [CrossRef]
- Lanner, J.; Kratschmer, S.; Petrović, B.; Gaulhofer, F.; Meimberg, H.; Pachinger, B. City Dwelling Wild Bees: How Communal Gardens Promote Species Richness. Urban Ecosyst. 2019, 23, 271–288. [Google Scholar] [CrossRef]
- Stewart, A.B.; Sritongchuay, T.; Teartisup, P.; Kaewsomboon, S.; Bumrungsri, S. Habitat and landscape Factors Influence Pollinators in a Tropical Megacity, Bangkok, Thailand. PeerJ 2018, 6, e5335. [Google Scholar] [CrossRef] [PubMed]
- Werrell, P.A.; Langellotto, G.A.; Morath, S.U.; Matteson, K.C. The Influence of Garden Size and Floral Cover on Pollen Deposition in Urban Community Gardens. Cities Environ. 2009, 2, 1–16. [Google Scholar] [CrossRef]
- Morales, C.L.; Traveset, A. A meta-analysis of impacts of alien vs. native plants on pollinator visitation and reproductive success of co-flowering native plants. Ecol. Lett. 2009, 12, 716–728. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hostetler, N.E.; McIntyre, M.E. Effects of Urban Land Use on Pollinator (Hymenoptera: Apoidea) Communities in a Desert Metropolis. Basic Appl. Ecol. 2001, 2, 209–218. [Google Scholar] [CrossRef] [Green Version]
- Mach, B.M.; Potter, D.A. Quantifying Bee Assemblages and Attractiveness of Flowering Woody Landscape Plants for Urban Pollinator Conservation. PLoS ONE 2018, 13, e0208428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Giovanetti, M.; Giuliani, C.; Boff, S.; Fico, G.; Lupi, D. A Botanic Garden as a Tool to Combine Public Perception of Nature and Life-Science Investigations on Native/Exotic Plants Interactions with Local Pollinators. PLoS ONE 2020, 15, e0228965. [Google Scholar] [CrossRef] [Green Version]
- Rollings, R.; Goulson, D. Quantifying the Attractiveness of Garden Flowers for Pollinators. J. Insect Conserv. 2019, 23, 803–817. [Google Scholar] [CrossRef] [Green Version]
- Salisbury, A.; Armitage, J.; Bostock, H.; Perry, J.; Tatchell, M.; Thompson, K. EDITOR’S CHOICE: Enhancing Gardens as Habitats for Flower-Visiting Aerial Insects (pollinators): Should We Plant Native or Exotic Species? J. Appl. Ecol. 2015, 52, 1156–1164. [Google Scholar] [CrossRef]
- Lowenstein, D.M.; Matteson, K.C.; Minor, E. Evaluating the Dependence of Urban Pollinators on Ornamental, Non-native, and ‘Weedy’ Floral Resources. Urban Ecosyst. 2019, 22, 293–302. [Google Scholar] [CrossRef]
- Aronson, M.F.J.; Lepczyk, C.; Evans, K.L.; Goddard, M.; Lerman, S.B.; MacIvor, J.S.; Nilon, C.H.; Vargo, T. Biodiversity in the City: Key Challenges for Urban Green Space Management. Front. Ecol. Environ. 2017, 15, 189–196. [Google Scholar] [CrossRef] [Green Version]
- Lepczyk, C.; Aronson, M.F.J.; Evans, K.L.; Goddard, M.A.; Lerman, S.B.; MacIvor, J.S. Biodiversity in the City: Fundamental Questions for Understanding the Ecology of Urban Green Spaces for Biodiversity Conservation. BioScience 2017, 67, 799–807. [Google Scholar] [CrossRef] [Green Version]
- Konvicka, M.; Kadlec, T. How to Increase The Value of Urban Areas for Butterfly Conservation? A Lesson from Prague Nature Reserves and Parks. Eur. J. Èntomol. 2011, 108, 219–229. [Google Scholar] [CrossRef] [Green Version]
- Tzortzakaki, O.; Kati, V.; Panitsa, M.; Tzanatos, E.; Giokas, S. Butterfly Diversity along the Urbanization Gradient in a Densely-Built Mediterranean City: Land Cover is More Decisive than Resources in Structuring Communities. Landsc. Urban Plan. 2019, 183, 79–87. [Google Scholar] [CrossRef]
- Jauker, B.; Krauss, J.; Jauker, F.; Steffan-Dewenter, I. Linking Life History Traits to Pollinator Loss in Fragmented Calcareous Grasslands. Landsc. Ecol. 2012, 28, 107–120. [Google Scholar] [CrossRef] [Green Version]
- Gunnarsson, B.; Federsel, L.M. Bumblebees in the city: Abundance, Species Richness and Diversity in Two Urban Habitats. J. Insect Conserv. 2014, 18, 1185–1191. [Google Scholar] [CrossRef] [Green Version]
- Zurbuchen, A.; Landert, L.; Klaiber, J.; Müller, A.; Hein, S.; Dorn, S. Maximum Foraging Ranges in Solitary Bees: Only Few Individuals Have the Capability to Cover Long Foraging Distances. Biol. Conserv. 2010, 143, 669–676. [Google Scholar] [CrossRef]
- Lerman, S.B.; Contosta, A.R.; Milam, J.; Bang, C. To Mow or to Mow Less: Lawn Mowing Frequency Affects Bee Abundance and Diversity in Suburban Yards. Biol. Conserv. 2018, 221, 160–174. [Google Scholar] [CrossRef]
- Dylewski, Ł.; Maćkowiak, Ł.; Banaszak-Cibicka, W. Are All Urban Green Spaces a Favourable Habitat for Pollinator Communities? Bees, Butterflies and Hoverflies in Different Urban Green Areas. Ecol. Èntomol. 2019, 44, 678–689. [Google Scholar] [CrossRef]
- Normandin, É.É.; Vereecken, N.J.; Buddle, C.C.; Fournier, V.V. Taxonomic and Functional Trait Diversity of Wild Bees in Different Urban Settings. PeerJ 2017, 5, e3051. [Google Scholar] [CrossRef] [Green Version]
- Daniels, B.; Jedamski, J.; Ottermanns, R.; Ross-Nickoll, M. A “plan bee” for cities: Pollinator diversity and plant-pollinator interactions in urban green spaces. PLoS ONE 2020, 15, e0235492. [Google Scholar] [CrossRef] [PubMed]
- Middle, I.; Dzidic, P.; Buckley, A.; Bennett, D.; Tye, M.; Jones, R. Integrating Community Gardens into Public Parks: An Innovative Approach for Providing Ecosystem Services in Urban Areas. Urban For. Urban Green. 2014, 13, 638–645. [Google Scholar] [CrossRef] [Green Version]
- Fischer, L.K.; Eichfeld, J.; Kowarik, I.; Buchholz, S. Disentangling Urban Habitat and Matrix Effects on Wild Bee Species. PeerJ 2016, 4, e2729. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hunter, P. Brown is the new green: Brownfield sites often harbour a surprisingly large amount of biodiversity. EMBO Rep. 2014, 15, 1238–1242. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Twerd, L.; Banaszak-Cibicka, W. Wastelands: Their attractiveness and importance for preserving the diversity of wild bees in urban areas. J. Insect Conserv. 2019, 23, 573–588. [Google Scholar] [CrossRef] [Green Version]
- Turo, K.J.; Spring, M.R.; Sivakoff, F.S.; De La Flor, Y.A.D.; Gardiner, M.M. Conservation in post-industrial cities: How does vacant land management and landscape configuration influence urban bees? J. Appl. Ecol. 2021, 58, 58–69. [Google Scholar] [CrossRef]
- Baldock, K.C. Opportunities and Threats for Pollinator Conservation in Global Towns and Cities. Curr. Opin. Insect Sci. 2020, 38, 63–71. [Google Scholar] [CrossRef]
- Buchholz, S.; Egerer, M. Functional Ecology of Wild Bees in Cities: Towards a Better Understanding of trait-Urbanization Relationships. Biodivers. Conserv. 2020, 29, 2779–2801. [Google Scholar] [CrossRef]
- Cariveau, D.P.; Winfree, R. Causes of Variation in Wild Bee Responses to Anthropogenic Drivers. Curr. Opin. Insect Sci. 2015, 10, 104–109. [Google Scholar] [CrossRef]
- Hernandez, J.L.; Frankie, G.W.; Thorp, R.W. Ecology of Urban Bees: A Review of Current Knowledge and Directions for Future Study. Cities Environ. 2009, 2, 1–15. [Google Scholar] [CrossRef]
- Da Rocha-Filho, L.C.; Ferreira-Caliman, M.J.; Garófalo, C.A.; Augusto, S.C. A Specialist in an Urban Area: Are Cities Suitable to Harbour Populations of the Oligolectic Bee Centris (Melacentris) collaris (Apidae: Centridini)? Ann. Zool. Fenn. 2018, 55, 135–149. [Google Scholar] [CrossRef]
- Garbuzov, M.; Alton, K.; Ratnieks, F.L. Most Ornamental Plants on Sale in Garden Centres are Unattractive to Flower-Visiting Insects. PeerJ 2017, 5, e3066. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frankie, G.W.; Thorp, R.W.; Schindler, M.; Hernandez, J.; Ertter, B.; Rizzardi, M. Ecological Patterns of Bees and Their Host Ornamental Flowers in Two Northern California Cities. J. Kans. Èntomol. Soc. 2005, 78, 227–246. [Google Scholar] [CrossRef]
- Geslin, B.; Gauzens, B.; Thébault, E.; Dajoz, I. Plant Pollinator Networks along a Gradient of Urbanisation. PLoS ONE 2013, 8, e63421. [Google Scholar] [CrossRef] [PubMed]
- Michener, C.D. The Bees of the World, 2nd ed.; Johns Hopkins University Press: Baltimore, MD, USA, 2007. [Google Scholar]
- Verboven, H.A.F.; Uyttenbroeck, R.; Brys, R.; Hermy, M. Different Responses of Bees and Hoverflies to Land Use in an Urban–Rural Gradient Show the Importance of the Nature of the Rural Land Use. Landsc. Urban Plan. 2014, 126, 31–41. [Google Scholar] [CrossRef]
- Hurd, P.D. An Annotated Catalogue of the Carpenter Bees (Genus Xylocopa Latrielle) of the Western Hemisphere (Hymenoptera: Anthophoridae); Smithsonian Institute Press: Washington, DC, USA, 1978. [Google Scholar]
- Fortel, L.; Henry, M.; Guilbaud, L.; Guirao, A.L.; Kuhlmann, M.; Mouret, H.; Rollin, O.; Vaissière, B.E. Decreasing Abundance, Increasing Diversity and Changing Structure of the Wild Bee Community (Hymenoptera: Anthophila) along an Urbanization Gradient. PLoS ONE 2014, 9, e104679. [Google Scholar] [CrossRef]
- Wray, J.C.; Neame, L.A.; Elle, E. Floral Resources, Body Size, and Surrounding Landscape Influence Bee Community Assemblages in Oak-Savannah Fragments: Bee Communities in Oak-Savannah Fragments. Ecol. Èntomol. 2014, 39, 83–93. [Google Scholar] [CrossRef]
- Eggenberger, H.; Frey, D.; Pellissier, L.; Ghazoul, J.; Fontana, S.; Moretti, M. Urban Bumblebees Are Smaller and More Phenotypically Diverse than Their Rural Counterparts. J. Anim. Ecol. 2019, 88, 1522–1533. [Google Scholar] [CrossRef]
- Merckx, T.; Kaiser, A.; Van Dyck, H. Increased Body Size along Urbanization Gradients at Both Community and Intraspecific Level in Macro-Moths. Glob. Chang. Biol. 2018, 24, 3837–3848. [Google Scholar] [CrossRef]
- Merckx, T.; Souffreau, C.; Kaiser, A.; Baardsen, L.F.; Backeljau, T.; Bonte, D.; Brans, K.I.; Cours, M.; Dahirel, M.; Debortoli, N.; et al. Body-Size Shifts in Aquatic and Terrestrial Urban Communities. Nat. Cell Biol. 2018, 558, 113–116. [Google Scholar] [CrossRef]
- Wilson, E.S.; Murphy, C.E.; Rinehart, J.P.; Yocum, G.; Bowsher, J.H. Microclimate Temperatures Impact Nesting Preference in Megachile rotundata (Hymenoptera: Megachilidae). Environ. Èntomol. 2020, 49, 296–303. [Google Scholar] [CrossRef] [PubMed]
- Buchholz, S.; Kowarik, I. Urbanisation Modulates Plant-Pollinator Interactions in Invasive vs. Native Plant Species. Sci. Rep. 2019, 9, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Harrison, T.; Gibbs, J.; Winfree, R. Forest Bees Are Replaced in Agricultural and Urban Landscapes by Native Species with Different Phenologies and Life-History Traits. Glob. Chang. Biol. 2017, 24, 287–296. [Google Scholar] [CrossRef] [PubMed]
- Goulson, D.; Nicholls, E.; Botías, C.; Rotheray, E.L. Bee Declines Driven by Combined Stress from Parasites, Pesticides, and Lack of Flowers. Science 2015, 347, 1255957. [Google Scholar] [CrossRef]
- Harrison, T.; Gibbs, J.; Winfree, R. Phylogenetic Homogenization of Bee Communities across Ecoregions: XXXX. Glob. Ecol. Biogeogr. 2018, 27, 1457–1466. [Google Scholar] [CrossRef]
- Odanaka, K.A.; Rehan, S.M. Impact Indicators: Effects of Land Use Management on Functional Trait and Phylogenetic Diversity of Wild Bees. Agric. Ecosyst. Environ. 2019, 286, 106663. [Google Scholar] [CrossRef]
- Xu, J.; Dang, H.; Wang, M.; Chai, Y.; Guo, Y.; Chen, Y.; Zhang, C.; Yue, M. Is Phylogeny More Useful than Functional Traits for Assessing Diversity Patterns Under Community Assembly Processes? Forests 2019, 10, 1159. [Google Scholar] [CrossRef] [Green Version]
- Baiser, B.; Lockwood, J.L. The relationship between functional and taxonomic homogenization: Functional and taxonomic homogenization. Glob. Ecol. Biogeogr. 2010, 20, 134–144. [Google Scholar] [CrossRef]
- Grab, H.; Branstetter, M.G.; Amon, N.; Urban-Mead, K.R.; Park, M.G.; Gibbs, J.; Blitzer, E.J.; Poveda, K.; Loeb, G.; Danforth, B.N. Agriculturally dominated landscapes reduce bee phylogenetic diversity and pollination services. Science 2019, 363, 282–284. [Google Scholar] [CrossRef]
- Matteson, K.C.; Ascher, J.S.; Gail, A.L. Bee Richness and Abundance in New York City Urban Gardens. Ann. Entomol. Soc. Am. 2008, 101, 140–150. [Google Scholar] [CrossRef]
- Sheffield, C.S.; Pindar, A.; Packer, L.; Kevan, P.G. The Potential of Cleptoparasitic Bees as Indicator Taxa for Assessing Bee Communities. Apidologie 2013, 44, 501–510. [Google Scholar] [CrossRef] [Green Version]
- Turo, K.J.; Gardiner, M.M. From Potential to Practical: Conserving Bees in Urban Public Green Spaces. Front. Ecol. Environ. 2019, 17, 167–175. [Google Scholar] [CrossRef]
- Howes, M.; Wortley, L.; Potts, R.; Dedekorkut-Howes, A.; Serrao-Neumann, S.; Davidson, J.; Smith, T.; Nunn, P. Environmental Sustainability: A Case of Policy Implementation Failure? Sustainability 2017, 9, 165. [Google Scholar] [CrossRef] [Green Version]
- Burr, A.; Hall, D.M.; Schaeg, N. The Perfect Lawn: Exploring Neighborhood Socio-Cultural Drivers for Insect Pollinator Habitat. Urban Ecosyst. 2018, 21, 1123–1137. [Google Scholar] [CrossRef]
- Byrne, L. Of Looks, Laws and Lawns: How Human Aesthetic Preferences Influence Landscape Management, Public Policies, and Urban Ecosystems. In Proceedings of the Urban Rural Interface Conference Proceedings, Atlanta, GA, USA, 13–16 March 2005; pp. 42–46. [Google Scholar]
- Zhao, C.; Sander, H.A.; Hendrix, S.D. Wild Bees and Urban Agriculture: Assessing Pollinator Supply and Demand across Urban Landscapes. Urban Ecosyst. 2019, 22, 455–470. [Google Scholar] [CrossRef]
- Hunter, R.; Cleland, C.; Cleary, A.; Droomers, M.; Wheeler, B.; Sinnett, D.; Nieuwenhuijsen, M.J.; Braubach, M. Environmental, Health, Wellbeing, Social and Equity Effects of Urban Green Space Interventions: A Meta-Narrative Evidence Synthesis. Environ. Int. 2019, 130, 104923. [Google Scholar] [CrossRef]
- Keeler, B.; Hamel, P.; McPhearson, T.; Hamann, M.H.; Donahue, M.L.; Prado, K.A.M.; Arkema, K.K.; Bratman, G.N.; Brauman, K.A.; Finlay, J.; et al. Social-Ecological and Technological Factors Moderate the Value of Urban Nature. Nat. Sustain. 2019, 2, 29–38. [Google Scholar] [CrossRef]
- Wolch, J.R.; Byrne, J.; Newell, J.P. Urban Green Space, Public Health, and Environmental Justice: The Challenge of Making Cities ‘Just Green Enough’. Landsc. Urban Plan. 2014, 125, 234–244. [Google Scholar] [CrossRef] [Green Version]
- Kondo, M.C.; Fluehr, J.M.; McKeon, T.P.; Branas, C.C. Urban Green Space and Its Impact on Human Health. Int. J. Environ. Res. Public Health 2018, 15, 445. [Google Scholar] [CrossRef] [Green Version]
- Duchemin, E.; Wegmuller, F.; Legault, A.-M. Urban Agriculture: Multi-Dimensional Tools for Social Development in Poor Neibourghoods. Field Actions Sci. Rep. 2009, 2, 1–8. [Google Scholar] [CrossRef]
- Tibesigwa, B.; Ntuli, H.; Lokina, R. Valuing Recreational Ecosystem Services in Developing Cities: The Case of Urban Parks in Dar Es Salaam, Tanzania. Cities 2020, 106, 102853. [Google Scholar] [CrossRef]
- Aguilera, G.; Ekroos, J.; Persson, A.S.; Pettersson, L.B.; Öckinger, E. Intensive Management Reduces Butterfly Diversity Over Time in Urban Green Spaces. Urban Ecosyst. 2018, 22, 335–344. [Google Scholar] [CrossRef] [Green Version]
- MacIvor, J.S.; Cabral, J.M.; Packer, L. Pollen Specialization by Solitary Bees in an Urban Landscape. Urban Ecosyst. 2013, 17, 139–147. [Google Scholar] [CrossRef]
- Harmon-Threatt, A.N.; Hendrix, S.D. Prairie Restorations and Bees: The Potential Ability of Seed Mixes to Foster Native Bee Communities. Basic Appl. Ecol. 2015, 16, 64–72. [Google Scholar] [CrossRef]
- Sikora, A.; Michołap, P.; Sikora, M. What Kind of Flowering Plants Are Attractive for Bumblebees in urban Green Areas? Urban For. Urban Green. 2020, 48, 126546. [Google Scholar] [CrossRef]
- Tucker, E.M.; Rehan, S.M. Wild Bee Pollination Networks in Northern New England. J. Insect Conserv. 2016, 20, 325–337. [Google Scholar] [CrossRef]
- Hall, D.M.; Steiner, R. Insect Pollinator Conservation Policy Innovations at Subnational Levels: Lessons for Lawmakers. Environ. Sci. Policy 2019, 93, 118–128. [Google Scholar] [CrossRef]
- City of Toronto. Toronto Pollinator Protection Strategy. 2018. Available online: https://www.toronto.ca/wp-content/uploads/2018/05/9676-A1802734_pollinator-protection-strategy-booklet.pdf (accessed on 22 November 2020).
- Xerces Society. c 2020. Bee City USA [Internet]. Xerces Society. Available online: https://www.beecityusa.org/ (accessed on 20 November 2020).


Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ayers, A.C.; Rehan, S.M. Supporting Bees in Cities: How Bees Are Influenced by Local and Landscape Features. Insects 2021, 12, 128. https://doi.org/10.3390/insects12020128
Ayers AC, Rehan SM. Supporting Bees in Cities: How Bees Are Influenced by Local and Landscape Features. Insects. 2021; 12(2):128. https://doi.org/10.3390/insects12020128
Chicago/Turabian StyleAyers, Anthony C., and Sandra M. Rehan. 2021. "Supporting Bees in Cities: How Bees Are Influenced by Local and Landscape Features" Insects 12, no. 2: 128. https://doi.org/10.3390/insects12020128