
Identification and Characterization Analysis of Transient Receptor Potential Mucolipin Protein of Laodelphax striatellus Fallén


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insects and Cells
2.2. Bioinformatics
2.3. RNA Extraction and RT-qPCR Assay
2.4. Baculovirus Expression of Ls-TRPML in Sf9 Cells
2.5. Antibody Preparation
2.6. Immunofluorescence Microscopy
2.7. Plasmid Construction
2.8. Protein Expression and Purification
2.9. SDS–PAGE and Western Blot Assay
2.10. Lipid Binding Assay
2.11. Statistical Analysis
3. Results
3.1. Identification of TRPML in L. striatellus
3.2. The Expression Profile Analysis of Ls-TRPML
3.3. The Subcellular Localization of Ls-TRPML in Sf9 Cells
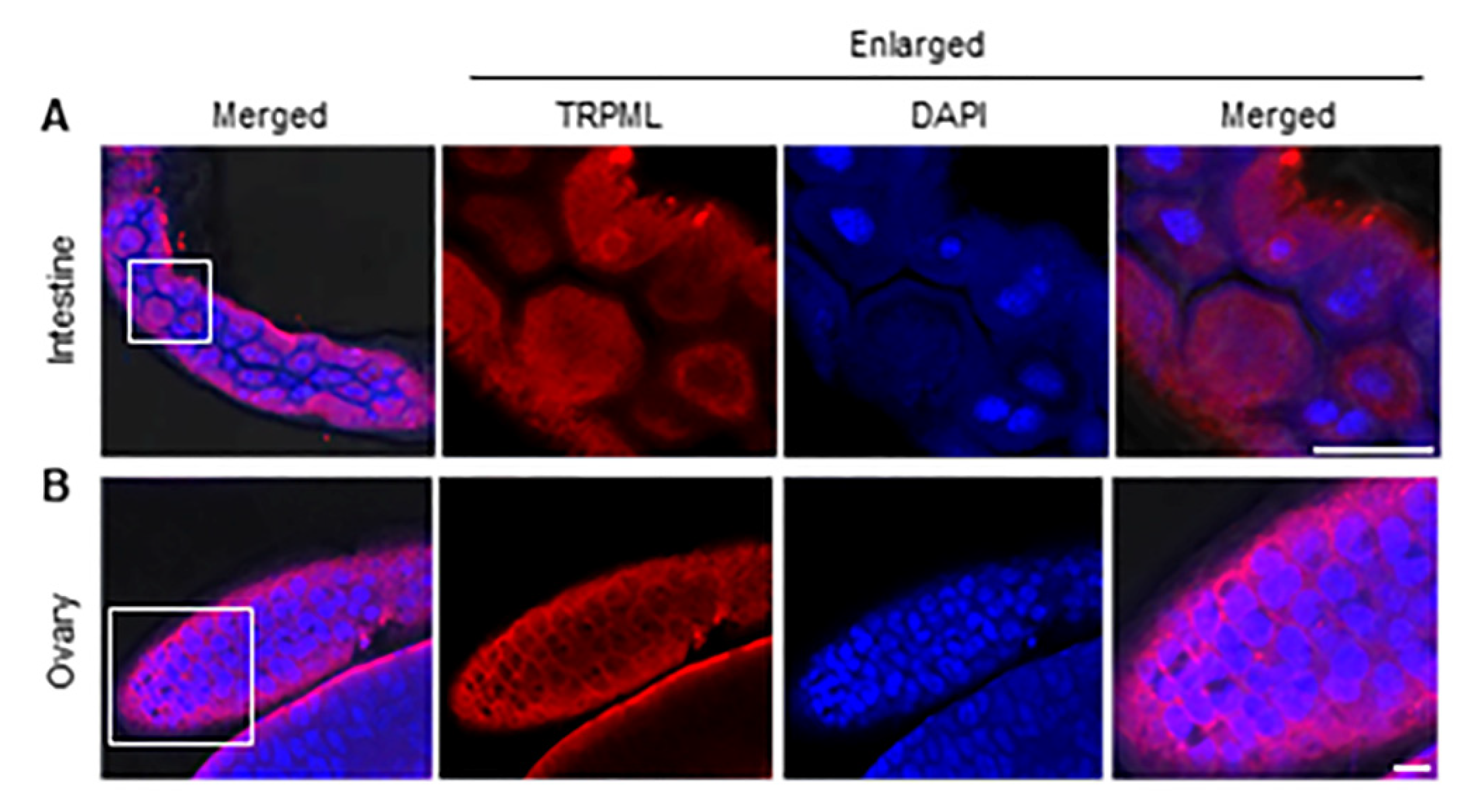
3.4. The Subcellular Localization of Ls-TRPML in L. striatellus
3.5. The Specific Binding of Ls-TRPML with PI(3,5)P2
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Montell, C. Physiology, phylogeny, and functions of the TRP superfamily of cation channels. Sci. STKE 2001, 90, re1. [Google Scholar] [CrossRef]
- Montell, C. The TRP superfamily of cation channels. Sci. STKE 2005, 2005, re3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramsey, I.S.; Delling, M.; Clapham, D.E. An introduction to TRP channels. Annu. Rev. Physiol. 2006, 68, 619–647. [Google Scholar] [CrossRef] [Green Version]
- Moran, M.M.; Xu, H.; Clapham, D.E. TRP ion channels in the nervous system. Curr. Opin. Neurobiol. 2004, 14, 362–369. [Google Scholar] [CrossRef]
- Colletti, G.A.; Kiselyov, K. TRPML1. Adv. Exp. Med. Biol. 2011, 704, 209–219. [Google Scholar]
- Cheng, X.; Shen, D.; Samie, M.; Xu, H. Mucolipins: Intracellular TRPML1-3 channels. FEBS Lett. 2010, 584, 2013–2021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Feng, X.; Huang, Y.; Lu, Y.; Xiong, J.; Wong, C.O.; Yang, P.; Xia, J.; Chen, D.; Du, G.; Venkatachalam, K.; et al. Drosophila TRPML forms PI(3,5)P2-activated cation channels in both endolysosomes and plasma membrane. J. Biol. Chem. 2014, 289, 4262–4272. [Google Scholar] [CrossRef] [Green Version]
- Feng, X.; Xiong, J.; Lu, Y.; Xia, X.; Zhu, M.X. Differential mechanisms of action of the mucolipin synthetic agonist, ML-SA1, on insect TRPML and mammalian TRPML1. Cell Calcium 2014, 56, 446–456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkatachalam, K.; Wong, C.O.; Zhu, M.X. The role of TRPMLs in endolysosomal trafficking and function. Cell Calcium 2015, 58, 48–56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.X.; Ren, D. Lysosomal physiology. Annu. Rev. Physiol. 2015, 77, 57–80. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di-Paolo, G.; De-Camilli, P. Phosphoinositides in cell regulation and membrane dynamics. Nature 2006, 443, 651–657. [Google Scholar] [CrossRef]
- Dong, X.P.; Shen, D.B.; Wang, X.; Dawson, T.; Li, X.R.; Zhang, Q.; Cheng, X.P.; Zhang, Y.L.; Weisman, L.S.; Delling, M.; et al. PI(3,5)P(2) controls membrane trafficking by direct activation of mucolipin Ca(2+) release channels in the endolysosome. Nat. Commun. 2010, 1, 38. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.L.; Cheng, X.P.; Yu, L.; Yang, J.S.; Calvo, R.; Patnaik, S.; Hu, X.; Gao, Q.; Yang, M.M.; Lawas, M.; et al. MCOLN1 is a ROS sensor in lysosomes that regulates autophagy. Nat. Commun. 2016, 7, 12109. [Google Scholar] [CrossRef] [Green Version]
- Zhang, X.L.; Yu, L.; Xu, H.X. Lysosome calcium in ROS regulation of autophagy. Autophagy 2016, 12, 1954–1955. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, X.R.; Garrity, A.G.; Xu, H.X. Regulation of membrane trafficking by signalling on endosomal and lysosomal membranes. J. Physiol. 2013, 591, 4389–4401. [Google Scholar] [CrossRef] [Green Version]
- Venkatachalam, K.; Long, A.A.; Elsaesser, R.; Nikolaeva, D.; Broadie, K.; Montell, C. Motor deficit in a Drosophila model of mucolipidosis type IV due to defective clearance of apoptotic cells. Cell 2008, 135, 838–851. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Edwards-Jorquera, S.S.; Bosveld, F.; Bellaïche, Y.A.; Lennon-Duménil, A.M.; Glavic, A. Trpml controls actomyosin contractility and couples migration to phagocytosis in fly macrophages. J. Cell Biol. 2020, 219, e201905228. [Google Scholar] [CrossRef] [PubMed]
- Fares, H.; Greenwald, I. Regulation of endocytosis by CUP-5, the Caenorhabditis elegans mucolipin-1 homolog. Nat. Genet. 2001, 28, 64–68. [Google Scholar] [CrossRef] [PubMed]
- Treusch, S.; Knuth, S.; Slaugenhaupt, S.A.; Goldin, E.; Grant, B.D.; Fares, H. Caenorhabditis elegans functional orthologue of human protein h-mucolipin-1 is required for lysosome biogenesis. Proc. Natl. Acad. Sci. USA 2004, 101, 4483–4488. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hersh, B.M.; Hartwieg, E.; Horvitz, H.R. The Caenorhabditis elegans mucolipin-like gene cup-5 is essential for viability and regulates lysosomes in multiple cell types. Proc. Natl. Acad. Sci. USA 2002, 99, 4355–4360. [Google Scholar] [CrossRef] [Green Version]
- Schaheen, L.; Dang, H.; Fares, H. Basis of lethality in C. elegans lacking CUP-5, the Mucolipidosis Type IV orthologue. Dev. Biol. 2006, 293, 382–391. [Google Scholar] [CrossRef] [Green Version]
- Campbell, E.M.; Fares, H. Roles of CUP-5, the Caenorhabditis elegans orthologue of human TRPML1, in lysosome and gut granule biogenesis. BMC Cell Biol. 2010, 11, 40. [Google Scholar] [CrossRef] [Green Version]
- Sun, T.; Wang, X.; Lu, Q.; Ren, H.; Zhang, H. CUP-5, the C. elegans ortholog of the mammalian lysosomal channel protein MLN1/TRPML1, is required for proteolytic degradation in autolysosomes. Autophagy 2011, 7, 1308–1315. [Google Scholar] [CrossRef] [Green Version]
- Luisoni, E.; Lovisolo, O.; Kitagawa, Y.; Shikata, E. Serological relationship between maize rough dwarf virus and rice black-streaked dwarf virus. Virology 1973, 52, 281–283. [Google Scholar] [CrossRef]
- Xu, Y.; Fu, S.; Tao, X.R.; Zhou, X.P. Rice stripe virus: Exploring molecular weapons in the arsenal of a negative-sense RNA Virus. Annu. Rev. Phytopathol. 2021, 59, 351–371. [Google Scholar] [CrossRef] [PubMed]
- Di, D.P.; Zhang, Y.L.; Yan, C.; Yan, T.; Zhang, A.H.; Yang, F.; Cao, X.L.; Li, D.W.; Lu, Y.G.; Wang, X.B.; et al. First report of barley yellow striate mosaic virus on wheat in China. Plant Dis. 2014, 98, 1450. [Google Scholar] [CrossRef]
- Wu, W.; Liu, H.Q.; Dong, Y.; Zhang, Y.; Wong, X.M.; Wang, C.C.; Zhou, Y.J.; Xu, Q.F. Determination of suitable RT-qPCR reference genes for studies of gene functions in Laodelphax striatellus (Fallén). Genes 2014, 10, 887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Venkatachalam, K.; Wong, C.O.; Montell, C. Feast or famine: Role of TRPML in preventing cellular amino acid starvation. Autophagy 2013, 9, 98–100. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, X.L.; Li, X.R.; Xu, H.X. Phosphoinositide isoforms determine compartments pecific ion channel activity. Proc. Natl. Acad. Sci. USA 2012, 109, 11384–11389. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, C.C.; Keller, M.; Hess, M.; Schiffmann, R.; Urban, N.; Wolfgardt, A.; Schaefer, M.; Brecher, F.; Biel, M.; Wahl-Schott, C.; et al. A small molecule restores function to TRPML1 mutant isoforms responsible for mucolipidosis type IV. Nat. Commun. 2014, 5, 4681. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grimm, C.; Jörs, S.; Saldanha, S.A.; Obukhov, A.G.; Pan, B.; Oshima, K.; Cuajungco, M.P.; Chase, P.; Hodder, P.; Heller, S. Small molecule activators of TRPML 3. Chem. Biol. 2010, 17, 135–148. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mochizuki, T.; Wu, G.; Hayashi, T.; Xenophontos, S.L.; Veldhuisen, B.; Saris, J.J.; Reynolds, D.M.; Cai, Y.; Gabow, P.A.; Pierides, A.; et al. PKD2, a gene for polycystic kidney disease that encodes an integral membrane protein. Science 1996, 272, 1339–1342. [Google Scholar] [CrossRef] [PubMed]
- Santoni, G.; Morelli, M.B.; Amantini, C.; Nabissi, M.; Santoni, M.; Santoni, A. Involvement of the TRPML mucolipin channels in viral infections and anti-viral innate immune responses. Front. Immunol. 2020, 11, 739. [Google Scholar] [CrossRef]
- Wong, C.O.; Li, R.; Montell, C.; Venkatachalam, K. Drosophila TRPML is required for TORC1 activation. Curr. Biol. 2012, 22, 1616–1621. [Google Scholar] [CrossRef] [Green Version]
- Björkgren, I.; Lishko, P.V. Fertility and TRP Channels. In Neurobiology of TRP Channels; Emir, T.L.R., Ed.; CRC Press/Taylor & Francis: Boca Raton, FL, USA, 2017; Chapter 11. [Google Scholar]
- Whitaker, M. Calcium at fertilization and in early development. Physiol. Rev. 2006, 86, 25–88. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Machaty, Z. Calcium influx in mammalian eggs. Reproduction 2013, 145, R97–R105. [Google Scholar] [CrossRef] [Green Version]
- Kaneuchi, T.; Sartain, C.V.; Takeo, S.; Horner, V.L.; Buehner, N.A.; Aigaki, T.; Wolfner, M.F. Calcium waves occur as Drosophila oocytes activate. Proc. Natl. Acad. Sci. USA 2015, 112, 791–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cao, Q.; Yang, Y.; Zhong, X.Z.; Dong, X.P. The lysosomal Ca(2+) release channel TRPML1 regulates lysosome size by activating calmodulin. J. Biol. Chem. 2017, 292, 8424–8435. [Google Scholar] [CrossRef] [Green Version]
- Ho, C.Y.; Alghamdi, T.A.; Botelho, R.J. Phosphatidylinositol-3,5-bisphosphate: No longer the poor PIP2. Traffic 2012, 13, 1–8. [Google Scholar] [CrossRef]
- Kingsolver, M.B.; Huang, Z.J.; Hardy, R.W. Insect antiviral innate immunity: Pathways, effectors, and connections. J. Mol. Biol. 2013, 425, 4921–4936. [Google Scholar] [CrossRef] [Green Version]
- Wei, J.; Jia, D.S.; Mao, Q.Z.; Zhang, X.F.; Chen, Q.; Wu, W.; Chen, H.Y.; Wei, T.Y. Complex interactions between insect-borne rice viruses and their vectors. Curr. Opin. Virol. 2018, 33, 18–23. [Google Scholar] [CrossRef] [PubMed]
- Schoggins, J.W.; Rice, C.M. Interferon-stimulated genes and their antiviral effector functions. Curr. Opin. Virol. 2011, 1, 519–525. [Google Scholar] [CrossRef] [PubMed]
- Schoggins, J.W. Interferon-stimulated genes: Roles in viral pathogenesis. Curr. Opin. Virol. 2014, 6, 40–46. [Google Scholar] [CrossRef] [PubMed]
- Rinkenberger, N.; Schoggins, J.W. Mucolipin-2 cation channel increases trafficking efficiency of endocytosed viruses. MBio 2018, 9, e2314–e2317. [Google Scholar] [CrossRef] [Green Version]
- Sun, L.; Hua, Y.; Vergarajauregui, S.; Diab, H.I.; Puertollano, R. Novel role of TRPML2 in the regulation of the innate immune response. J. Immunol. 2015, 195, 4922–4932. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Valadez, J.A.; Cuajungco, M.P. PAX5 is the transcriptional activator of mucolipin-2 (MCOLN2) gene. Gene 2015, 555, 194–202. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cuajungco, M.P.; Silva, J.; Habibi, A.; Valadez, J.A. The mucolipin-2 (TRPML2) ion channel: A tissue-specific protein crucial to normal cell function. Pflügers Arch. Eur. J. Physiol. 2016, 468, 177–192. [Google Scholar] [CrossRef] [Green Version]





Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, H.; Dong, Y.; Wan, B.; Ji, Y.; Xu, Q. Identification and Characterization Analysis of Transient Receptor Potential Mucolipin Protein of Laodelphax striatellus Fallén. Insects 2021, 12, 1107. https://doi.org/10.3390/insects12121107
Wang H, Dong Y, Wan B, Ji Y, Xu Q. Identification and Characterization Analysis of Transient Receptor Potential Mucolipin Protein of Laodelphax striatellus Fallén. Insects. 2021; 12(12):1107. https://doi.org/10.3390/insects12121107
Chicago/Turabian StyleWang, Haitao, Yan Dong, Baijie Wan, Yinghua Ji, and Qiufang Xu. 2021. "Identification and Characterization Analysis of Transient Receptor Potential Mucolipin Protein of Laodelphax striatellus Fallén" Insects 12, no. 12: 1107. https://doi.org/10.3390/insects12121107