
Deep Conservation of Hid-Like RHG Gene Family Homologs in Winged Insects Revealed by “Taxon Hopping” BLAST

Abstract
:Simple Summary
Abstract
1. Introduction
2. Results
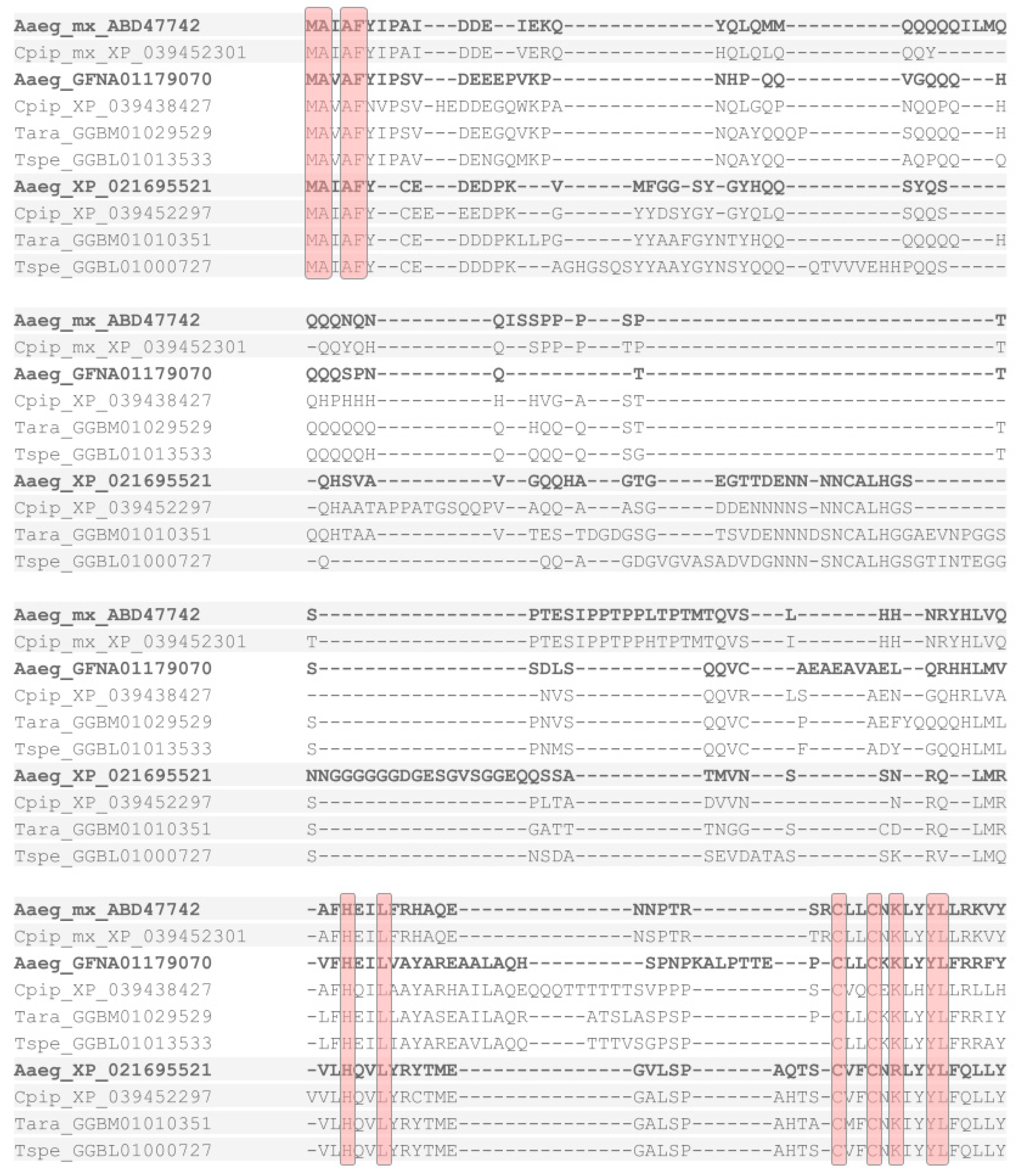
2.1. RHG Homologs from an Extended Range of Winged Insects
2.2. Protein Sequence Conservation Differences within and between Orders
2.3. Deeply Conserved N- and C-Terminal Amino Acid Residues
2.4. Michelob_x Constitutes an Independent RHG Gene Family Expansion in Mosquitoes
2.5. Gene Structure Conservation
2.6. A Conserved RHG Isoform in the Lepidoptera
2.7. Exceptional RHG Sequence Divergence in Aphids
3. Discussion
4. Materials and Methods
4.1. Homolog Searches
4.2. Multiple Sequence Alignments
4.3. Gene Structure Analyses
Supplementary Materials
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yuan, J.; Shaham, S.; Ledoux, S.; Ellis, H.M.; Horvitz, H.R. The C. elegans cell death gene Ced-3 encodes a protein similar to mammalian Interleukin-1 Beta-Converting enzyme. Cell 1993, 75, 641–652. [Google Scholar] [CrossRef]
- Crawford, E.D.; Seaman, J.E.; Barber, A.E.; David, D.C.; Babbitt, P.C.; Burlingame, A.L.; Wells, J.A. Conservation of Caspase substrates across metazoans suggests hierarchical importance of signaling pathways over specific targets and cleavage site motifs in apoptosis. Cell Death Differ. 2012, 19, 2040–2048. [Google Scholar] [CrossRef] [Green Version]
- Cory, S.; Adams, J.M. The Bcl2 Family: Regulators of the cellular Life-or-Death switch. Nat. Rev. Cancer 2002, 2, 647–656. [Google Scholar] [CrossRef]
- Lettre, G.; Hengartner, M.O. Developmental apoptosis in C. elegans: A complex CEDnario. Nat. Rev. Mol. Cell Biol. 2006, 7, 97–108. [Google Scholar] [CrossRef]
- Wang, S.L.; Hawkins, C.J.; Yoo, S.J.; Müller, H.A.; Hay, B.A. The Drosophila Caspase Inhibitor DIAP1 Is essential for cell survival and is negatively regulated by HID. Cell 1999, 98, 453–463. [Google Scholar] [CrossRef] [Green Version]
- Conradt, B. Genetic Control of Programmed Cell Death during Animal Development. Annu. Rev. Genet. 2009, 43, 493–523. [Google Scholar] [CrossRef] [Green Version]
- White, K.; Grether, M.E.; Abrams, J.M.; Young, L.; Farrell, K.; Steller, H. Genetic control of programmed cell death in Drosophila. Science 1994, 264, 677–683. [Google Scholar] [CrossRef] [PubMed]
- Grether, M.E.; Abrams, J.M.; Agapite, J.; White, K.; Steller, H. The head involution defective gene of Drosophila melanogaster functions in programmed cell death. Genes Dev. 1995, 9, 1694–1708. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, P.; Nordstrom, W.; Gish, B.; Abrams, J.M. Grim, a novel cell death gene in Drosophila. Genes Dev. 1996, 10, 1773–1782. [Google Scholar] [CrossRef] [Green Version]
- Wing, J.P.; Karres, J.S.; Ogdahl, J.L.; Zhou, L.; Schwartz, L.M.; Nambu, J.R. Drosophila sickle is a novel Grim-Reaper cell death activator. Curr. Biol. 2002, 12, 131–135. [Google Scholar] [CrossRef] [Green Version]
- Srinivasula, S.M.; Datta, P.; Kobayashi, M.; Wu, J.W.; Fujioka, M.; Hegde, R.; Zhang, Z.; Mukattash, R.; Fernandes-Alnemri, T.; Shi, Y.; et al. Sickle, a novel Drosophila death gene in the reaper/hid/grim region, encodes an IAP-Inhibitory protein. Curr. Biol. 2002, 12, 125–130. [Google Scholar] [CrossRef] [Green Version]
- Christich, A.; Kauppila, S.; Chen, P.; Sogame, N.; Ho, S.I.; Abrams, J.M. The damage-responsive Drosophila gene sickle encodes a novel IAP binding protein similar to but distinct from reaper, grim, and hid. Curr. Biol. 2002, 12, 137–140. [Google Scholar] [CrossRef] [Green Version]
- Zhou, L. The “unique Key”feature of the Iap-Binding Motifs in RHG Proteins. Cell Death Differ. 2005, 12, 1148–1151. [Google Scholar] [CrossRef] [PubMed]
- Shi, Y. A Conserved Tetrapeptide Motif: Potentiating Apoptosis through IAP-Binding. Cell Death Differ. 2002, 9, 93–95. [Google Scholar] [CrossRef] [PubMed]
- Berthelet, J.; Dubrez, L. Regulation of apoptosis by Inhibitors of Apoptosis (IAPs). Cells 2013, 2, 163–187. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zachariou, A.; Tenev, T.; Goyal, L.; Agapite, J.; Steller, H.; Meier, P. IAP-antagonists exhibit non-redundant modes of action through differential DIAP1 binding. EMBO J. 2003, 22, 6642–6652. [Google Scholar] [CrossRef] [Green Version]
- Wing, J.P.; Schwartz, L.M.; Nambu, J.R. The RHG Motifs of Drosophila Reaper and Grim are important for their distinct cell death-inducing abilities. Mech. Dev. 2001, 102, 193–203. [Google Scholar] [CrossRef]
- Clavería, C.; Caminero, E.; Martínez-A, C.; Campuzano, S.; Torres, M. GH3, a novel proapoptotic domain in Drosophila Grim, promotes a mitochondrial death pathway. EMBO J. 2002, 21, 3327–3336. [Google Scholar] [CrossRef] [Green Version]
- Ribeiro Lopes, M.; Parisot, N.; Callaerts, P.; Calevro, F. Genetic diversity of the apoptotic pathway in insects. In Evolution, Origin of Life, Concepts and Methods; Pontarotti, P., Ed.; Springer International Publishing: Cham, Switzerland, 2019; pp. 253–285. ISBN 9783030303631. [Google Scholar]
- Yoo, S.; Lam, H.; Lee, C.; Lee, G.; Park, J.H. Cloning and functional characterizations of an apoptogenic hid gene in the scuttle fly, Megaselia scalaris (Diptera; Phoridae). Gene 2017, 604, 9–21. [Google Scholar] [CrossRef] [Green Version]
- Chen, P.; Ho, S.-I.; Shi, Z.; Abrams, J.M. Bifunctional killing activity encoded by conserved Reaper proteins. Cell Death Differ. 2004, 11, 704–713. [Google Scholar] [CrossRef]
- Edman, R.M.; Linger, R.J.; Belikoff, E.J.; Li, F.; Sze, S.-H.; Tarone, A.M.; Scott, M.J. Functional characterization of calliphorid cell death genes and cellularization gene promoters for controlling gene Expression and cell viability in early embryos. Insect Mol. Biol. 2015, 24, 58–70. [Google Scholar] [CrossRef]
- Schetelig, M.F.; Nirmala, X.; Handler, A.M. Pro-apoptotic cell death Genes, hid and reaper, from the tephritid pest Sspecies, Anastrepha suspensa. Apoptosis 2011, 16, 759–768. [Google Scholar] [CrossRef] [PubMed]
- Bryant, B.; Zhang, Y.; Zhang, C.; Santos, C.P.; Clem, R.J.; Zhou, L. A lepidopteran orthologue of reaper reveals functional conservation and evolution of IAP antagonists. Insect Mol. Biol. 2009, 18, 341–351. [Google Scholar] [CrossRef] [Green Version]
- Wu, Y.; Wu, Y.; Hui, T.; Wu, H.; Wu, Y.; Wang, W. Reaper homologue IBM1 in silkworm Bombyx mori induces apoptosis upon baculovirus infection. FEBS Lett. 2013, 587, 600–606. [Google Scholar] [CrossRef] [Green Version]
- Shu, B.; Zhang, J.; Veeran, S.; Zhong, G. Pro-apoptotic function analysis of the reaper homologue IBM1 in Spodoptera frugiperda. Int. J. Mol. Sci. 2020, 21, 2729. [Google Scholar] [CrossRef] [Green Version]
- Misof, B.; Liu, S.; Meusemann, K.; Peters, R.S.; Donath, A.; Mayer, C.; Frandsen, P.B.; Ware, J.; Flouri, T.; Beutel, R.G.; et al. Phylogenomics resolves the timing and pattern of insect evolution. Science 2014, 346, 763–767. [Google Scholar] [CrossRef]
- Lopes, M.R.; Parisot, N.; Gaget, K.; Huygens, C.; Peignier, S.; Duport, G.; Orlans, J.; Charles, H.; Baatsen, P.; Jousselin, E.; et al. Evolutionary novelty in the apoptotic pathway of aphids. Proc. Natl. Acad. Sci. USA 2020, 117, 32545–32556. [Google Scholar] [CrossRef]
- Colella, S.; Parisot, N.; Simonet, P.; Gaget, K.; Duport, G.; Baa-Puyoulet, P.; Rahbé, Y.; Charles, H.; Febvay, G.; Callaerts, P.; et al. Bacteriocyte reprogramming to cope with nutritional stress in a phloem sap feeding hemipteran, the pea aphid Acyrthosiphon pisum. Front. Physiol. 2018, 9, 1498. [Google Scholar] [CrossRef]
- Richards, S.; Gibbs, R.A.; Weinstock, G.M.; Brown, S.J.; Denell, R.; Beeman, R.W.; Gibbs, R.; Beeman, R.W.; Brown, S.J.; Bucher, G.; et al. The genome of the model beetle and pest Tribolium castaneum. Nature 2008, 452, 949–955. [Google Scholar] [PubMed] [Green Version]
- Herndon, N.; Shelton, J.; Gerischer, L.; Ioannidis, P.; Ninova, M.; Dönitz, J.; Waterhouse, R.M.; Liang, C.; Damm, C.; Siemanowski, J.; et al. Enhanced genome assembly and a new official gene set for Tribolium castaneum. BMC Genom. 2020, 21, 47. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhou, L.; Jiang, G.; Chan, G.; Santos, C.P.; Severson, D.W.; Xiao, L. Michelob_x is the missing inhibitor of apoptosis protein antagonist in mosquito genomes. EMBO Rep. 2005, 6, 769–774. [Google Scholar] [CrossRef] [PubMed]
- Zdobnov, E.M.; von Mering, C.; Letunic, I.; Torrents, D.; Suyama, M.; Copley, R.R.; Christophides, G.K.; Thomasova, D.; Holt, R.A.; Subramanian, G.M.; et al. Comparative genome and proteome analysis of Anopheles gambiae and Drosophila melanogaster. Science 2002, 298, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Christophides, G.K.; Zdobnov, E.; Barillas-Mury, C.; Birney, E.; Blandin, S.; Blass, C.; Brey, P.T.; Collins, F.H.; Danielli, A.; Dimopoulos, G.; et al. Immunity-related genes and gene families in Anopheles gambiae. Science 2002, 298, 159–165. [Google Scholar] [CrossRef] [Green Version]
- Meng, X.; Zhu, F.; Chen, K. Silkworm: A promising model organism in life science. J. Insect Sci. 2017, 17, 97. [Google Scholar] [CrossRef]
- Brown, S.J.; Shippy, T.D.; Miller, S.; Bolognesi, R.; Beeman, R.W.; Lorenzen, M.D.; Bucher, G.; Wimmer, E.A.; Klingler, M. The Red flour beetle, Tribolium castaneum (Coleoptera): A model for studies of development and pest biology. Cold Spring Harb. Protoc. 2009, 2009, db.emo126. [Google Scholar] [CrossRef]
- Werren, J.H.; Loehlin, D.W. The parasitoid wasp Nasonia: An emerging model system with haploid male genetics. Cold Spring Harb. Protoc. 2009, 2009, db–emo134. [Google Scholar] [CrossRef] [Green Version]
- Chipman, A.D. Oncopeltus fasciatus as an evo-devo research organism. Genesis 2017, 55, e23020. [Google Scholar] [CrossRef]
- Zhan, S.; Merlin, C.; Boore, J.L.; Reppert, S.M. The monarch butterfly genome yields insights into long-distance migration. Cell 2011, 147, 1171–1185. [Google Scholar] [CrossRef] [Green Version]
- Kawahara, A.Y.; Plotkin, D.; Espeland, M.; Meusemann, K.; Toussaint, E.F.A.; Donath, A.; Gimnich, F.; Frandsen, P.B.; Zwick, A.; Dos Reis, M.; et al. Phylogenomics reveals the evolutionary timing and pattern of butterflies and moths. Proc. Natl. Acad. Sci. USA 2019, 116, 22657–22663. [Google Scholar] [CrossRef] [Green Version]
- International Aphid Genomics Consortium Genome sequence of the pea aphid Acyrthosiphon pisum. PLoS Biol. 2010, 8, e1000313.
- Julca, I.; Marcet-Houben, M.; Cruz, F.; Vargas-Chavez, C.; Johnston, J.S.; Gómez-Garrido, J.; Frias, L.; Corvelo, A.; Loska, D.; Cámara, F.; et al. Phylogenomics identifies an ancestral burst of gene duplications predating the diversification of Aphidomorpha. Mol. Biol. Evol. 2020, 37, 730–756. [Google Scholar] [CrossRef] [PubMed]
- Schlötterer, C.; Tautz, D. Slippage Synthesis of Simple Sequence DNA. Nucleic Acids Res. 1992, 20, 211–215. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Noé, L.; Kucherov, G. YASS: Enhancing the sensitivity of DNA similarity search. Nucleic Acids Res. 2005, 33, W540-3. [Google Scholar] [CrossRef] [Green Version]
- Haining, W.N.; Carboy-Newcomb, C.; Wei, C.L.; Steller, H. The proapoptotic function of Drosophila hid is conserved in mammalian cells. Proc. Natl. Acad. Sci. USA 1999, 96, 4936–4941. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sandu, C.; Ryoo, H.D.; Steller, H. Drosophila IAP antagonists form multimeric complexes to promote cell death. J. Cell Biol. 2010, 190, 1039–1052. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Clem, R.J. The Role of IAP Antagonist proteins in the core apoptosis pathway of the mosquito disease vector Aedes aegypti. Apoptosis 2011, 16, 235–248. [Google Scholar] [CrossRef] [Green Version]
- Oakley, T.H.; Ostman, B.; Wilson, A.C.V. Repression and loss of gene expression outpaces activation and gain in recently duplicated fly genes. Proc. Natl. Acad. Sci. USA 2006, 103, 11637–11641. [Google Scholar] [CrossRef] [Green Version]
- Force, A.; Lynch, M.; Pickett, F.B.; Amores, A.; Yan, Y.L.; Postlethwait, J. Preservation of duplicate genes by complementary, degenerative mutations. Genetics 1999, 151, 1531–1545. [Google Scholar] [CrossRef]
- Chen, Z.-X.; Sturgill, D.; Qu, J.; Jiang, H.; Park, S.; Boley, N.; Suzuki, A.M.; Fletcher, A.R.; Plachetzki, D.C.; FitzGerald, P.C.; et al. Comparative validation of the D. melanogaster modENCODE transcriptome annotation. Genome Res. 2014, 24, 1209–1223. [Google Scholar] [CrossRef] [Green Version]
- Pruitt, K.D.; Tatusova, T.; Maglott, D.R. NCBI Reference Sequence (RefSeq): A Curated Non-Redundant Sequence Database of Genomes, Transcripts and Proteins. Nucleic Acids Res. 2005, 33, D501-4. [Google Scholar] [CrossRef] [Green Version]
- Altschul, S.F.; Koonin, E.V. Iterated Profile Searches with PSI-BLAST—a Tool for Discovery in Protein Databases. Trends Biochem. Sci. 1998, 23, 444–447. [Google Scholar] [CrossRef]
- McGinnis, S.; Madden, T.L. BLAST: At the Core of a Powerful and Diverse Set of Sequence Analysis Tools. Nucleic Acids Res. 2004, 32, W20-5. [Google Scholar] [CrossRef] [PubMed]
- Löytynoja, A.; Goldman, N. webPRANK: A Phylogeny-Aware Multiple Sequence Aligner with Interactive Alignment Browser. BMC Bioinform. 2010, 11, 579. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sievers, F.; Wilm, A.; Dineen, D.; Gibson, T.J.; Karplus, K.; Li, W.; Lopez, R.; McWilliam, H.; Remmert, M.; Söding, J.; et al. Fast, Scalable Generation of High-quality Protein Multiple Sequence Alignments Using Clustal Omega. Mol. Syst. Biol. 2011, 7, 539. [Google Scholar] [CrossRef]
- Notredame, C.; Higgins, D.G.; Heringa, J. T-Coffee: A Novel Method for Fast and Accurate Multiple Sequence Alignment. J. Mol. Biol. 2000, 302, 205–217. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Average Lengths | % Conserved Sites | Divergence Times | % Divergence/Million Years | |
|---|---|---|---|---|
| Diptera (n = 13) | 302.0 | 17.2% | 200 | 0.41% |
| Lepidoptera (n = 13) | 210.1 | 54.7% | 120 | 0.38% |
| Coleoptera: Polyphaga (n = 24) | 189.3 | 4.2% | 250 | 0.38% |
| Hymenoptera (n = 19) | 244.9 | 54.3% | 250 | 0.18% |
| Hemiptera wo Aphidoidea (n = 8) | 225.5 | 24.4% | 260 | 0.29% |
| Aphidoidea (n = 10) | 252.8 | 25.3% | 25 | 2.99% |
| Dictyoptera (n = 3) | 253.3 | 56.1% | 175 | 0.25% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the author. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Friedrich, M. Deep Conservation of Hid-Like RHG Gene Family Homologs in Winged Insects Revealed by “Taxon Hopping” BLAST. Insects 2021, 12, 957. https://doi.org/10.3390/insects12110957
Friedrich M. Deep Conservation of Hid-Like RHG Gene Family Homologs in Winged Insects Revealed by “Taxon Hopping” BLAST. Insects. 2021; 12(11):957. https://doi.org/10.3390/insects12110957
Chicago/Turabian StyleFriedrich, Markus. 2021. "Deep Conservation of Hid-Like RHG Gene Family Homologs in Winged Insects Revealed by “Taxon Hopping” BLAST" Insects 12, no. 11: 957. https://doi.org/10.3390/insects12110957