
Do Hydrothermal Shrimp Smell Vents?


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Collection
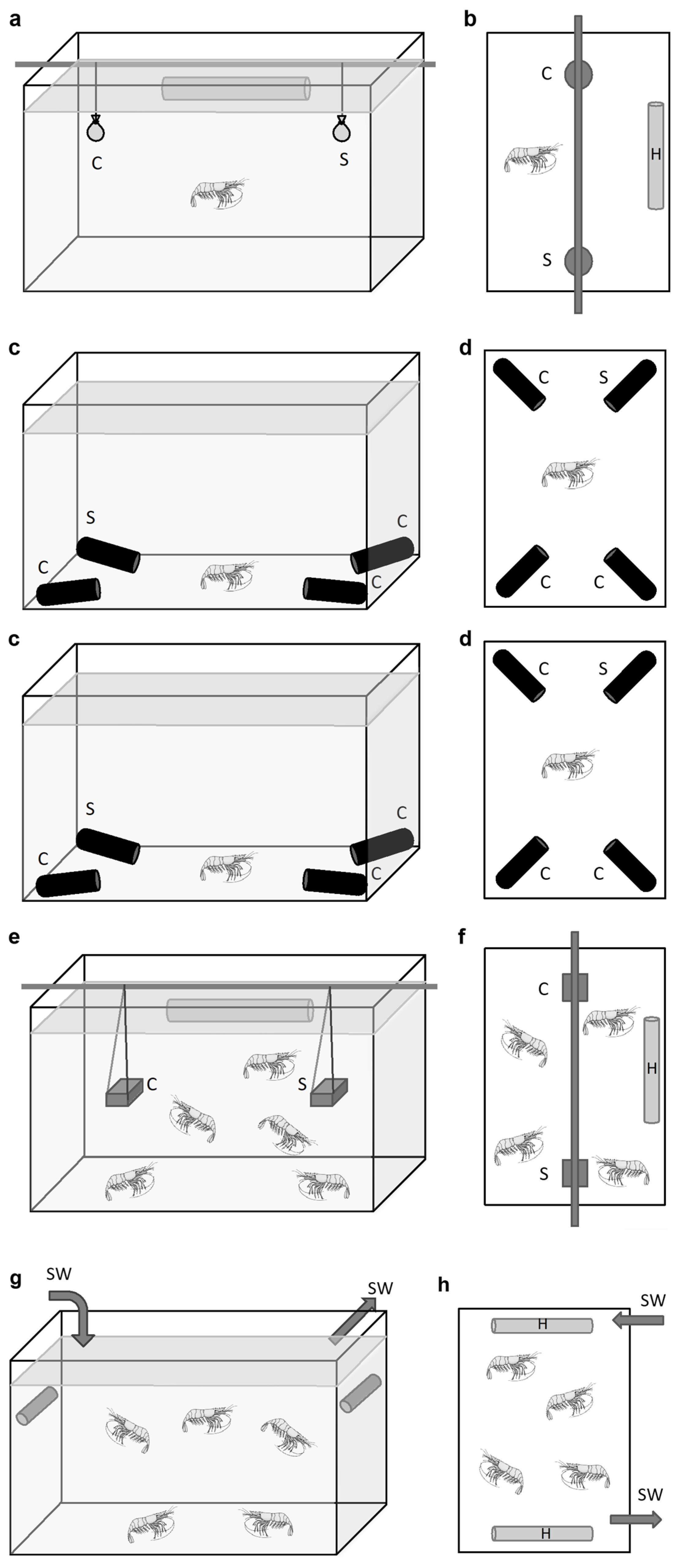
2.2. Behavior Experiments at In Situ Pressure on Hydrothermal Shrimp in Pressurized Aquaria
- -
- Four for grooming (3 batches of n = 3 adults and one batch of n = 8 juveniles of R. exoculata; total n = 17);
- -
- Two for sulfide pulses (one batch of n = 20 M. fortunata tested twice, and one batch of n = 20 R. exoculata tested once; total n = 40);
- -
- Eight for food and sulfide stimuli (3 batches of n = 10 R. exoculata for sulfide pH11 and 3 batches of n = 10 for sulfide pH4, and 2 batches of n = 5 or 6 M. fortunata for food; total n = 71);
- -
- One for temperature pulses (one batch of n = 20 adults and n = 10 juveniles of R. exoculata).
2.2.1. Grooming Behavior in Rimicaris exoculata
- -
- Batch 1: The specimens were collected at the Rainbow site (2300-m depth), and 3 individuals were transferred to IPOCAMP at 23 MPa and 10 °C upon their arrival on board;
- -
- Batch 2: The specimens were collected at the Broken Spur site (3100-m depth), and transferred to IPOCAMP at 30 MPa for maintenance at 20 °C. After 7 h, the aquarium was opened to select 3 individuals who were placed in a cage and further repressurized at 30 MPa and 20 °C in another IPOCAMP aquarium for the grooming observations;
- -
- Batch 3: The specimens were collected at the Broken Spur site, and transferred to BALIST aquarium at 30 MPa for maintenance at 10 °C. After 48 h, the aquarium was opened to select 3 adult individuals that were placed in a cage and further repressurized in IPOCAMP at 30 MPa and 10 °C for the grooming observations. After 96 h, the BALIST aquarium was opened again to select 8 juvenile individuals who were transferred to IPOCAMP for the same grooming observations as the adults.
2.2.2. Responses to Sulfide Pulse Stimuli on a Batch of Mirocaris fortunata and Rimicaris exoculata
2.2.3. Response to Food and Sulfide Stimuli during Experiments on a Batch of Mirocaris fortunata and Rimicaris exoculata
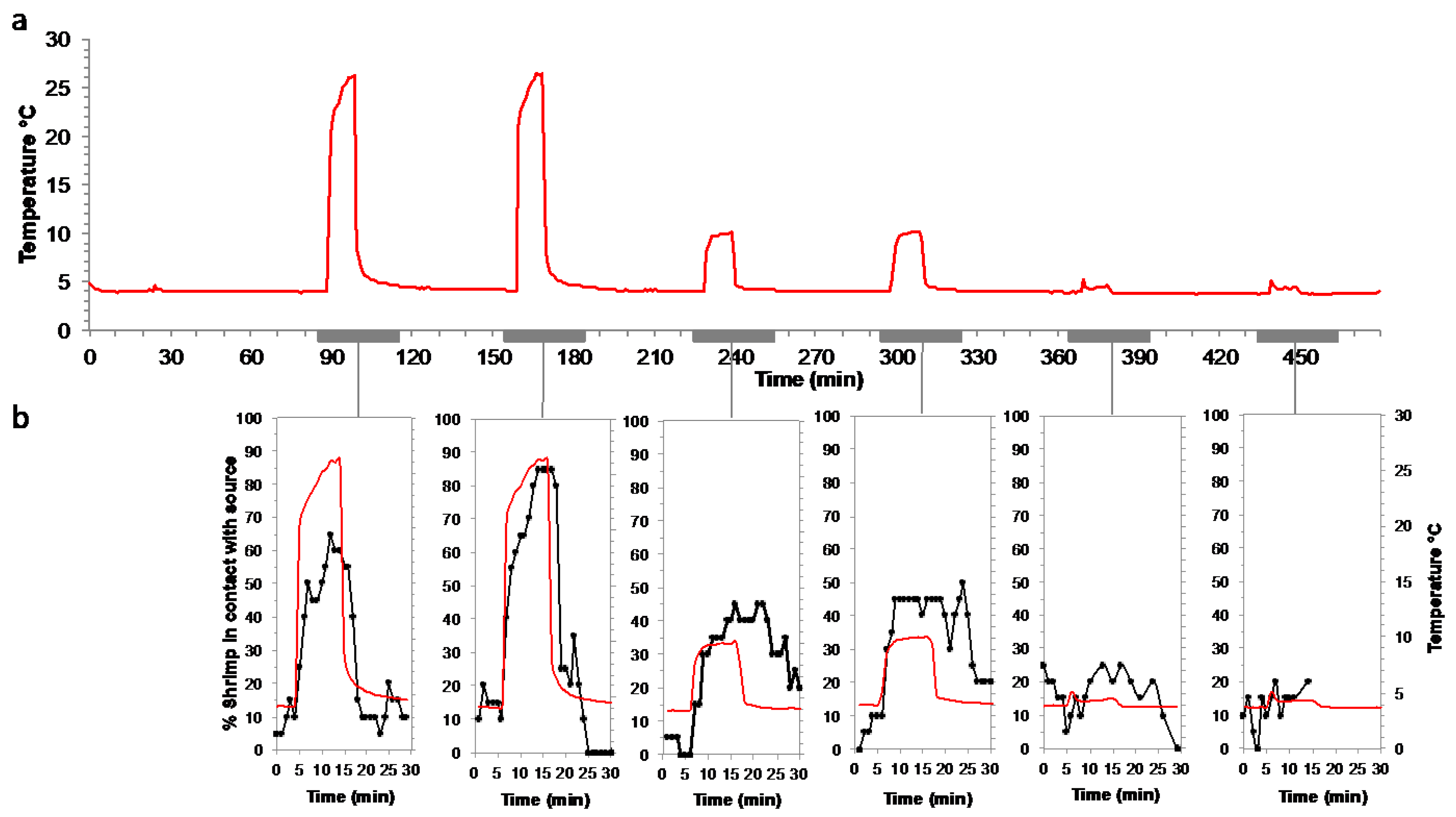
2.2.4. Responses to Warm Temperature Pulses at In Situ Pressure on a Batch of Rimicaris exoculata
2.3. Behavior Experiments at Atmospheric Pressure on Hydrothermal and Coastal Shrimp
2.3.1. Grooming Behavior in Palaemon serratus
2.3.2. Responses to Food Stimulus during Two-Choice Experiments on Single Individuals of Palaemon elegans and M. fortunata
2.3.3. Responses to Food and Sulfide Stimuli during Multiple-Choice Experiments on Single Individuals of Mirocaris fortunata and Palaemon elegans
2.3.4. Responses to Food and Sulfide Stimuli during Two-Choice Experiments on Multiple Individuals of M. fortunata and Palaemon elegans
2.3.5. Responses to Temperature Stimulus during Two-Choice Experiment on Multiple Individuals of Mirocaris fortunata and Palaemon elegans
2.4. Scanning Electron Microscopy (S.E.M.)
3. Results
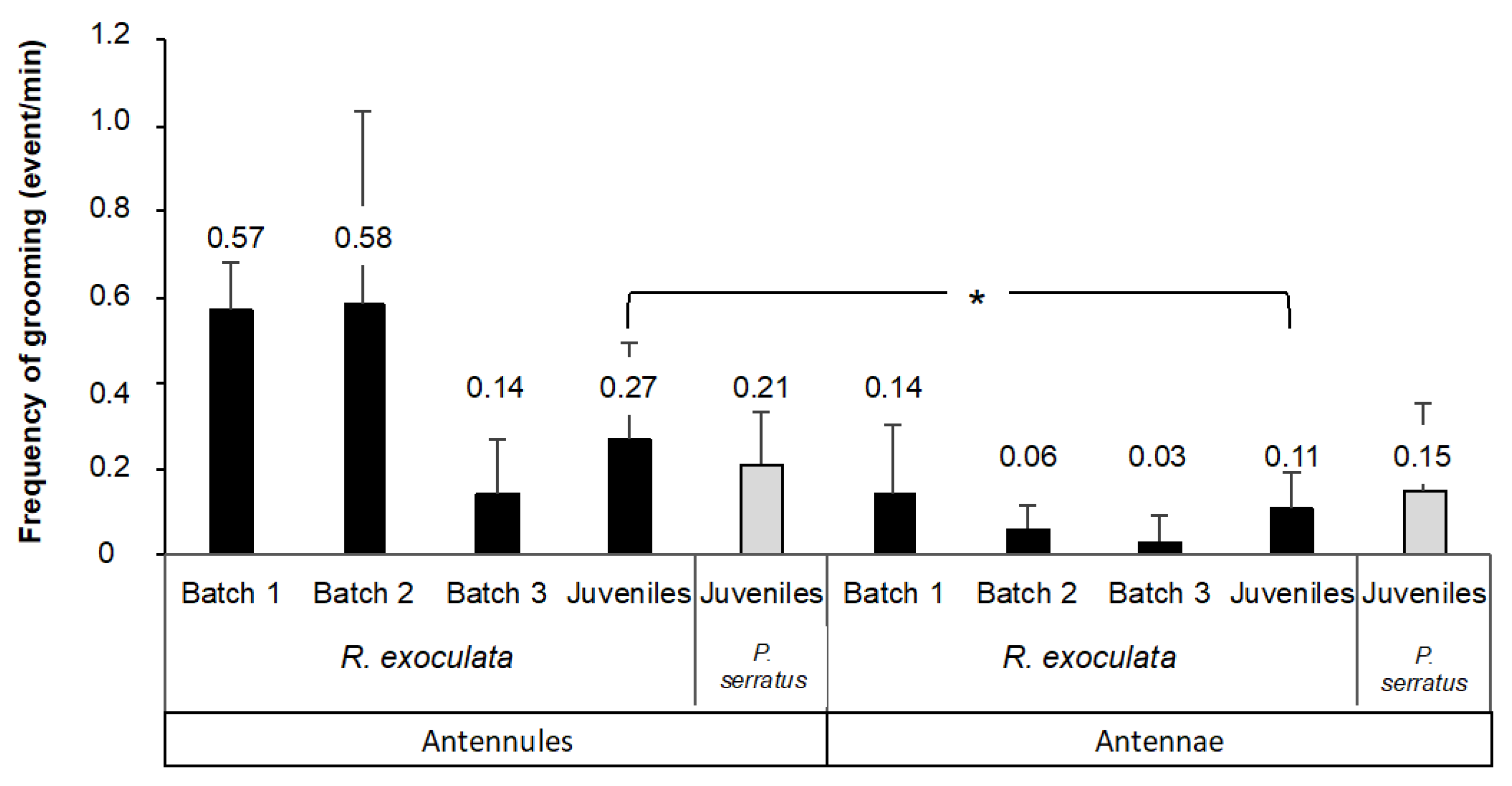
3.1. Grooming Behavior of Olfactory Appendages in Rimicaris exoculata and Palaemon serratus
3.2. Fouling of Olfactory Appendages in Rimicaris exoculata and Palaemon serratus
3.3. Experiments at In Situ Pressure on Mirocaris fortunata and Rimicaris exoculata—Attraction to Sulfide or Food Odor Stimuli
3.4. Experiments at Atmospheric Pressure on Mirocaris fortunata and Palaemon elegans—Attraction to Sulfide or Food Odor Stimuli
3.5. Attraction to Temperature on Rimicaris exoculata, Mirocaris fortunata and Palaemon elegans
4. Discussion
4.1. Grooming Behavior and Fouling of the Olfactory Appendages
4.2. Detection of Environmental Cues (Food Odors and Fluid Markers)
5. Conclusions
Author Contributions
Funding
International Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Steullet, P.; Dudar, O.; Flavus, T.; Zhou, M.; Derby, C.D. Selective ablation of antennular sensilla on the caribbean spiny lobster Panulirus argus suggests that dual antennular chemosensory pathways mediate odorant activation of searching and localization of food. J. Exp. Biol. 2001, 204, 4259–4269. [Google Scholar] [CrossRef]
- Derby, C.D.; Weissburg, M.J. The chemical senses and chemosensory ecology of crustaceans. In Crustacean Nervous Systems and Their Control of Behavior; Derby, C.D., Thiel, M., Eds.; Springer: New York, NY, USA, 2014; pp. 263–293. [Google Scholar]
- Schmidt, M.; Ache, B.W. Processing of antennular input in the brain of the spiny lobster, Panulirus argus. J. Comp. Phys. A 1996, 178, 605–628. [Google Scholar] [CrossRef]
- Ache, B.W. Chemoreception and thermoreception. In The Biology of Crustacea; Bliss, D., Ed.; Academic Press: New York, NY, USA, 1982; Volume 3, pp. 369–398. [Google Scholar]
- Derby, C.D.; Kozma, M.T.; Senatore, A.; Schmidt, M. Molecular mechanisms of reception and perireception in crustacean chemoreception: A comparative review. Chem. Senses 2016, 41, 381–398. [Google Scholar] [CrossRef] [Green Version]
- Schmitt, B.C.; Ache, B.W. Olfaction: Responses of a decapod crustacean are enhanced by flicking. Science 1979, 205, 204–206. [Google Scholar] [CrossRef] [PubMed]
- Devine, D.; Atema, J. Function of chemoreceptor organs in spatial orientation of the lobster, Homarus americanus: Differences and overlap. Biol. Bull. 1982, 163, 144–153. [Google Scholar] [CrossRef]
- Cowan, D.F. The role of olfaction in courtship behavior of the American lobster Homarus americanus. Biol. Bull. 1991, 181, 402–407. [Google Scholar] [CrossRef]
- Moore, P.A.; Scholz, N.; Atema, J. Chemical orientation of lobsters, Homarus americanus, in turbulent odor plumes. J. Chem. Ecol. 1991, 17, 1293–1307. [Google Scholar] [CrossRef] [PubMed]
- Derby, C.D.; Steullet, P. Why do animals have so many receptors? The role of multiple chemosensors in animal perception. Biol. Bull. 2001, 200, 211–215. [Google Scholar] [CrossRef] [PubMed]
- Shabani, S.; Kamio, M.; Derby, C.D. Spiny lobsters detect conspecific blood-borne alarm cues exclusively through olfactory sensilla. J. Exp. Biol. 2008, 211, 2600–2608. [Google Scholar] [CrossRef] [Green Version]
- Pond, D.W.; Segonzac, M.; Bell, M.V.; Dixon, D.R.; Fallick, A.E.; Sargent, J.R. Lipid and lipid carbonstable isotope composition of the hydrothermal vent shrimp Mirocaris fortunata: Evidence for nutritional dependence on photosynthetically fixed carbon. Mar. Ecol. Prog. Ser. 1997, 157, 221–231. [Google Scholar] [CrossRef] [Green Version]
- Herring, P.J.; Dixon, D.R. Extensive deep-sea dispersal of postlarval shrimp from a hydrothermal vent. Deep Sea Res. I 1998, 45, 2105–2118. [Google Scholar] [CrossRef]
- Sarradin, P.M.; Caprais, J.C.; Riso, R.; Kerouel, R.; Aminot, A. Chemical environment of the hydrothermal mussel communities in the Lucky Strike and Menez Gwen vent fields, Mid Atlantic Ridge. Cah. Biol. Mar. 1999, 40, 93–104. [Google Scholar]
- Sarrazin, J.; Juniper, S.K.; Massoth, G.; Legendre, P. Physical and chemical factors influencing species distributions on hydrothermal sulfide edifices of the Juan de Fuca Ridge, northeast Pacific. Mar. Ecol. Prog. Ser. 1999, 190, 89–112. [Google Scholar] [CrossRef]
- Le Bris, N.; Govenar, B.; Le Gall, C.; Fisher, C.R. Variability of physico-chemical conditions in 9°50′ N EPR diffuse flow vent habitats. Mar. Chem. 2006, 98, 167–182. [Google Scholar] [CrossRef]
- Pelli, D.G.; Chamberlain, S.C. The visibility of 350 °C black-body radiation by the shrimp Rimicaris exoculata and man. Nature 1989, 337, 460–461. [Google Scholar] [CrossRef] [PubMed]
- Charlou, J.L.; Donval, J.P.; Douville, E.; Jean-Baptiste, P.; Radford-Knoery, J.; Fouquet, Y.; Dapoigny, A.; Stievenard, M. Compared geochemical signatures and the evolution of Menez Gwen (37°50′ N) and Lucky Strike (37°17′ N) hydrothermal fluids, south of the Azores Triple Junction on the Mid-Atlantic Ridge. Chem. Geol. 2000, 171, 49–75. [Google Scholar] [CrossRef]
- Radford-Knoery, J.; Charlou, J.L.; Donval, J.P.; Aballea, M.; Fouquet, Y.; Ondreas, H. Distribution of dissolved sulfide, methane, and manganese near the seafloor at the Lucky Strike (37°17′ N) and Menez Gwen (37°50′ N) hydrothermal vent sites on the mid-Atlantic Ridge. Deep Sea Res. I 1998, 45, 367–386. [Google Scholar] [CrossRef]
- Tao, C.; Wu, T.; Liu, C.; Li, H.; Zhang, J. Fault inference and boundary recognition based on near-bottom magnetic data in the Longqi hydrothermal field. Mar. Geophys. Res. 2017, 38, 17–25. [Google Scholar] [CrossRef]
- Renninger, G.H.; Kass, L.; Gleeson, R.A.; van Dover, C.L.; Battelle, B.A.; Jinks, R.N.; Herzog, E.D.; Chamberlain, S.C. Sulfide as a chemical stimulus for deep-sea hydrothermal vent shrimp. Biol. Bull. 1995, 189, 69–76. [Google Scholar] [CrossRef] [PubMed]
- Jinks, R.N.; Battelle, B.A.; Herzog, E.D.; Kass, L.; Renninger, G.H.; Chamberlain, S.C. Sensory adaptations in hydrothermal vent shrimps from the Mid-Atlantic Ridge. Cah. Biol. Mar. 1998, 39, 309–312. [Google Scholar]
- Zbinden, M.; Berthod, C.; Montagné, N.; Machon, J.; Léger, N.; Chertemps, T.; Rabet, N.; Shillito, B.; Ravaux, J. Comparative study of chemosensory organs of shrimp from hydrothermal vent and coastal environments. Chem. Senses 2017, 42, 319–331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zbinden, M.; Gallet, A.; Szafranski, K.M.; Machon, J.; Ravaux, J.; Léger, N.; Duperron, S. Blow Your Nose, Shrimp! Unexpectedly Dense Bacterial Communities Occur on the Antennae and Antennules of Hydrothermal Vent Shrimp. Front. Mar. Sci. 2018, 5, 357. [Google Scholar] [CrossRef]
- Machon, J.; Lucas, P.; Ravaux, J.; Zbinden, M. Comparison of chemoreceptive abilities of the hydrothermal shrimp Mirocaris fortunata and the coastal shrimp Palaemon elegans. Chem. Senses 2018, 43, 489–501. [Google Scholar] [CrossRef] [PubMed]
- Machon, J.; Krieger, J.; Meth, R.; Zbinden, M.; Ravaux, J.; Montagné, N.; Chertemps, T.; Harzsch, S. Neuroanatomy of a hydrothermal vent shrimp provides insights into the evolution of crustacean integrative brain centers. eLife 2019, 8, e47550. [Google Scholar] [CrossRef]
- Machon, J. Sensory Adaptations in Shrimp from Deep Hydrothermal Vents: Comparison of Chemo- and Thermo-Sensory Abilities in the Vent Species Mirocaris fortunata and the Coastal Species Palaemon elegans. Ph.D. Thesis, Sorbonne Université, Paris, France, 2018. [Google Scholar]
- Bauer, R. Decapod crustaceans grooming: Functional morphology, adaptive value, and phylogenetic significance. In Functional Morphology of Feeding and Grooming in Crustacea; Felgenhauer, B.E., Watling, L., Thistle, A.A., Eds.; Brookfield: Rotterdam, The Netherlands, 1989; pp. 49–73. [Google Scholar]
- Barbato, J.; Daniel, P. Chemosensory activation of an antennular grooming behavior in the spiny lobster, Panulirus argus, is tunned narrowly to L-glutamate. Biol. Bull. 1997, 193, 107–115. [Google Scholar] [CrossRef] [PubMed]
- Bauer, R.T. Antifouling adaptations of marine shrimp (Crustacea: Decapoda: Caridea): Functional morphology and adaptive significance of antennular preening by the third maxillipeds. Mar. Biol. 1977, 40, 261–276. [Google Scholar] [CrossRef]
- Vismann, B. Sulfide species and total sulfide toxicity in the shrimp Crangon crangon. J. Exp. Mar. Biol. Ecol. 1996, 204, 141–154. [Google Scholar] [CrossRef]
- Stegner, M.E.; Stemme, T.; IIiffe, T.M.; Richter, S.; Wirkner, C.S. The brain in three crustaceans from cavernous darkness. BMC Neurosci. 2015, 16, 19. [Google Scholar] [CrossRef] [Green Version]
- Matabos, M.; Cuvelier, D.; Brouard, J.; Shillito, B.; Ravaux, J.; Zbinden, M.; Barthelemy, D.; Sarradin, P.M.; Sarrazin, J. Behavioural study of two hydrothermal crustacean decapods: Mirocaris fortunata and Segonzacia mesatlantica, from the Lucky Strike vent field (Mid-Atlantic Ridge). Deep Sea Res. II 2015, 121, 146–158. [Google Scholar] [CrossRef] [Green Version]
- Ravaux, J.; Cottin, D.; Chertemps, T.; Hamel, G.; Shillito, B. Hydrothermal vent shrimps display low expression of the heat-inducible hsp70 gene in nature. Mar. Ecol. Prog. Ser. 2009, 396, 153–156. [Google Scholar] [CrossRef] [Green Version]
- Shillito, B.; Hamel, G.; Duchi, C.; Cottin, D.; Sarrazin, J.; Sarradin, P.M.; Ravaux, J.; Gaill, F. Live capture of megafauna from 2300 m depth, using a newly designed pressurized recovery device. Deep Sea Res. I 2008, 55, 881–889. [Google Scholar] [CrossRef]
- Shillito, B.; Gaill, F.; Ravaux, J. The IPOCAMP pressure incubator for deep-sea fauna. J. Mar. Sci. Tech. 2014, 22, 97–102. [Google Scholar]
- Shillito, B.; Ravaux, J.; Sarrazin, J.; Zbinden, M.; Sarradin, P.M.; Barthelemy, D. Long-Term maintenance and public exhibition of deep-sea hydrothermal fauna: The AbyssBox project. Deep Sea Res. II 2015, 121, 137–145. [Google Scholar] [CrossRef] [Green Version]
- Shillito, B.; Desurmont, C.; Barthélémy, D.; Farabos, D.; Després, G.; Ravaux, J.; Zbinden, M.; Lamazière, A. Lipidome variations of dee-sea vent shrimps according to acclimation pressure: A homeoviscous response? Deep-sea Res. I 2020, 161, 103285. [Google Scholar] [CrossRef]
- Shillito, B.; Le Bris, N.; Hourdez, S.; Ravaux, J.; Cottin, D.; Caprais, J.C.; Jollivet, D.; Gaill, F. Temperature resistance studies on the deep-sea vent shrimp Mirocaris fortunata. J. Exp. Biol. 2006, 209, 945–955. [Google Scholar] [CrossRef] [Green Version]
- Sarrazin, J.; Legendre, P.; De Busserolles, F.; Fabri, M.C.; Guilini, K.; Ivanenko, V.N.; Morineaux, M.; Vanreusel, A.; Sarradin, P.M. Biodiversity patterns, environmental drivers and indicator species on a high-temperature hydrothermal edifice, Mid-Atlantic Ridge. Deep Sea Res. Part II Top. Stud. Oceanogr. 2015, 121, 177–192. [Google Scholar] [CrossRef] [Green Version]
- Ravaux, J.; Hamel, G.; Zbinden, M.; Tasiemski, A.A.; Boutet, I.; Léger, N.; Tanguy, A.; Jollivet, D.; Shillito, B. Thermal limit for metazoan life in question: In vivo heat tolerance of the Pompeii worm. PLoS ONE 2013, 8, e64074. [Google Scholar] [CrossRef] [Green Version]
- Van Maurik, L.N.; Wortham, J.L. Grooming as a secondary behavior in the shrimp Macrobrachium rosenbergii (Crustacea, Decapoda, Caridea). Zookeys 2014, 457, 55–77. [Google Scholar] [CrossRef] [Green Version]
- Corbari, L.; Zbinden, M.; Cambon-Bonavita, M.A.; Gaill, F.; Compère, P. Bacterial symbionts and mineral deposits in the branchial chamber of the hydrothermal vent shrimp Rimicaris exoculata: Relationship to moult cycle. Aqua Biol. 2008, 1, 225–238. [Google Scholar] [CrossRef] [Green Version]
- Kamio, M.; Derby, C. Finding food: How marine invertebrates use chemical cues to track and select food. Nat. Prod. Rep. 2017, 34, 514–528. [Google Scholar] [CrossRef]
- Wortham, J.L.; Pascual, S. Grooming behaviors and gill fouling un the commercially important bue crab (Callinectes sapidus) and stone crab (Menippe mercenaria). Nauplius 2017, 25, e2017028. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, M.; Chien, H.; Tadesse, T.; Johns, M.E.; Derby, C.D. Rosette-type tegumental glands associated with aesthetasc sensilla in the olfactory organ of the Caribbean spiny lobster, Panulirus argus. Cell Tissue Res. 2006, 325, 369–395. [Google Scholar] [CrossRef]
- Trapido-Rosenthal, H.; Gleeson, R.; Carr, W.E.S. The efflux of amino acids from the olfactory organ of the spiny lobster: Biochemical measurements and physiological effects. Biol. Bull. 1990, 179, 374–382. [Google Scholar] [CrossRef] [PubMed]
- Sarrazin, J.; Robigou, V.; Juniper, S.K.; Delaney, J.R. Biological and geological dynamics over four years on a high-temperature sulfide structure at the Juan de Fuca Ridge hydrothermal observatory. Mar. Ecol. Prog. Ser. 1997, 153, 5–24. [Google Scholar] [CrossRef] [Green Version]
- Sarrazin, J.; Levesque, C.; Juniper, S.; Tivey, M. Mosaic community dynamics on Juan de Fuca Ridge sulphide edifices: Substratum, temperature and implications for trophic structure. Cah. Biol. Mar. 2002, 43, 275–279. [Google Scholar]
- Gebruk, A.V.; Southward, E.C.; Kennedy, H.; Southward, A.J. Food sources, behaviour, and distribution of hydrothermal vent shrimps at the Mid-Atlantic Ridge. J. Mar. Biol. Assoc. UK 2000, 80, 485–499. [Google Scholar] [CrossRef]
- Desbruyères, D.; Biscoito, M.; Caprais, J.C.; Colaço, A.; Comtet, T.; Crassous, P.; Fouquet, Y.; Khripounoff, A.; Le Bris, N.; Olu, K.; et al. Variations in deep-sea hydrothermal vent communities on the Mid-Atlantic Ridge near the Azores plateau. Deep Sea Res II 2001, 48, 1325–1346. [Google Scholar] [CrossRef]
- Cuvelier, D.; Sarrazin, J.; Colaço, A.; Copley, J.; Desbruyères, D.; Glover, A.G.; Tyler, P.; Santos, R.S. Distribution and spatial variation of hydrothermal faunal assemblages at Lucky Strike (Mid-Atlantic Ridge) revealed by high-resolution video image analysis. Deep Sea Res. Part I Oceanogr. Res. Pap. 2009, 56, 2026–2040. [Google Scholar] [CrossRef] [Green Version]
- Zbinden, M.; Cambon-Bonavita, M.A. Rimicaris exoculata: Biology and ecology of a shrimp from deep-sea hydrothermal vents associated with ectosymbiotic bacteria. Mar. Ecol. Prog. Ser. 2020, 652, 187–222. [Google Scholar] [CrossRef]
- Kenning, M.; Lehmann, P.; Lindström, M.; Harzsch, S. Heading which way? Y-maze chemical assays: Not all crustaceans are alike. Helgoland Mar. Res. 2015, 69, 305–311. [Google Scholar] [CrossRef] [Green Version]
- Moore, P.A.; Grills, J.L. Chemical orientation to food by the crayfish Orconectes rusticus: Influence of hydrodynamics. Anim. Behav. 1999, 58, 953–963. [Google Scholar] [CrossRef] [PubMed]
- Segonzac, M.; De Saint Laurent, M.; Casanova, B. L’énigme du comportement trophique des crevettes Alvinocarididae des sites hydrothermaux de la dorsale médio-atlantique. Cah. Biol. Mar. 1993, 34, 535–571. [Google Scholar]
- Charlou, J.L.; Donval, J.P.; Fouquet, Y.; Jean-Baptiste, P.; Holm, N. Geochemistry of high H2 and CH4 vent fluids issuing from ultramafic rocks at the Rainbow hydrothermal field (36°14′N, MAR). Chem. Geol. 2002, 191, 345–359. [Google Scholar] [CrossRef]
- Portail, M.; Brandily, C.; Cathalot, C.; Colaço, A.; Gélinas, Y.; Husson, B.; Sarradin, P.M.; Sarrazin, J. Food-web complexity across hydrothermal vents on the Azores triple junction. Deep Sea Res. Part I Oceanogr. Res. Pap. 2018, 131, 101–120. [Google Scholar] [CrossRef]
- Zbinden, M.; Le Bris, N.; Gaill, F.; Compère, P. Distribution of bacteria and associated minerals in the gill chamber of the vent shrimp Rimicaris exoculata and related biogeochemical processes. Mar. Ecol. Prog. Ser. 2004, 284, 237–251. [Google Scholar] [CrossRef] [Green Version]
- Somero, G.N. Linking biogeography to physiology: Evolutionary and acclimatory adjustments of thermal limits. Front. Zool. 2005, 2, 1. [Google Scholar] [CrossRef] [Green Version]
- Lee, R.W. Thermal tolerances of deep-sea hydrothermal vent animals from the Northeast Pacific. Biol. Bull. 2003, 205, 98–101. [Google Scholar] [CrossRef]
- Bates, A.E.; Lee, R.W.; Tunnicliffe, V.; Lamare, M.D. Deep-sea hydrothermal vent animals seek cool fluids in a highly variable thermal environment. Nat. Commun. 2010, 1, 14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, F.; Brown, A.; Mestre, N.C.; Reed, A.J.; Thatje, S. Thermal adaptations in deep-sea hydrothermal vent and shallow-water shrimp. Deep Sea Res. Part II Top. Stud. Oceanogr. 2013, 92, 234–239. [Google Scholar] [CrossRef]
- Lagerspetz, K.Y.H.; Vainio, L.A. Thermal behaviour of crustaceans. Biol. Rev. 2006, 81, 237–258. [Google Scholar] [CrossRef]
- Jury, S.H.; Watson, W.H. 3rd. Thermosensitivity of the lobster, Homarus americanus, as determined by cardiac assay. Biol. Bull. 2000, 199, 257–264. [Google Scholar] [CrossRef]
- Forward, R.B. Behavioral responses of crustacean larvae to rates of temperature change. Biol. Bull. 1990, 178, 195–204. [Google Scholar] [CrossRef] [PubMed]
- Schmidt, C.; Le Bris, N.; Gaill, F. Interactions of deep-sea vent invertebrates with their environment: The case of Rimicaris exoculata. J. Shell. Res. 2008, 27, 9–90. [Google Scholar] [CrossRef] [Green Version]
- Baker, E.T.; Resing, J.A.; Haymon, R.M.; Tunnicliffe, V.; Lavelle, J.W.; Martinez, F.; Ferrini, V.; Walker, S.L.; Nakamura, K. How many vent fields? New estimates of vent field populations on ocean ridges from precise mapping of hydrothermal discharge locations. Earth Planet. Sci. Lett. 2016, 449, 186–196. [Google Scholar] [CrossRef]














{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stimulus | Hydrothermal Species | Coastal Species | |
|---|---|---|---|
| Mirocaris fortunata | Rimicaris exoculata | Palaemon elegans | |
| Food | - | nt | + |
| Food + Sulfide | - | nt | + |
| Sulfide | - | - | - |
| Temperature | + | + | - |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ravaux, J.; Machon, J.; Shillito, B.; Barthélémy, D.; Amand, L.; Cabral, M.; Delcour, E.; Zbinden, M. Do Hydrothermal Shrimp Smell Vents? Insects 2021, 12, 1043. https://doi.org/10.3390/insects12111043
Ravaux J, Machon J, Shillito B, Barthélémy D, Amand L, Cabral M, Delcour E, Zbinden M. Do Hydrothermal Shrimp Smell Vents? Insects. 2021; 12(11):1043. https://doi.org/10.3390/insects12111043
Chicago/Turabian StyleRavaux, Juliette, Julia Machon, Bruce Shillito, Dominique Barthélémy, Louis Amand, Mélanie Cabral, Elise Delcour, and Magali Zbinden. 2021. "Do Hydrothermal Shrimp Smell Vents?" Insects 12, no. 11: 1043. https://doi.org/10.3390/insects12111043