1. Introduction
Maize is a significant crop in China, especially since the country has become a net importer of grain [
1]. China is the world’s second-largest corn producer after the U.S. However, maize production has been threatened by fall armyworm (FAW),
Spodoptera frugiperda, J.E. Smith, 1797, (Lepidoptera: Noctuidae), which is highly ranked as one of the most destructive pests of the maize crop. This pest is native to the American continent, including tropical and subtropical regions [
2], and has recently invaded Africa, Asia, and Australia with a highly detrimental impact on global food security in all invaded areas. FAW was first reported in West Africa (Nigeria and Ghana) [
3] and later spread to other countries in sub-Saharan Africa [
4]. In May 2018, FAW was observed in Karnataka in the southwestern part of India [
5], and its occurrence was also reported in southeastern Asian countries, such as Thailand, Bangladesh, and Myanmar, by late 2018 [
6].
FAW was first reported in China on 11 December 2018 [
7], when the corn-specific strain of
S. frugiperda invaded [
8].
Spodoptera frugiperda invaded 26 provinces after migrating to China from Myanmar, and Guangxi and Yunnan were the major areas invaded by the pest [
9]. The larvae of
S. frugiperda have the potential to damage over 350 species of plants, including major staple and economic crops such as sorghum, corn, barley, rice, soybean, tobacco, tomato, and peanut [
10], but show a high preference for corn, sugarcane, and sorghum in China. During the early attack of
S. frugiperda larvae in 2019, extremely significant damage was caused to cornfields in Yunnan Province, China [
11].
Spodoptera frugiperda was reported to cause 67 and 22% maize yield losses in Zambia and Ghana, respectively [
12], while in Kenya and Ethiopia, this invasive pest caused maize yield losses of 47 and 32%, respectively [
13], resulting in economic losses of 1.08 and 4.66 billion USD in Africa [
14].
Spodoptera frugiperda larvae were also reported to feed on growing points and young organs in the peanut field and cause 78 and 65% damage at seedling and flowering stages, respectively [
15].
Spodoptera frugiperda causes serious damage (30 to 90%) in wheat fields, while 10 to 80% damage was recorded in the barley field [
16].
This invasive pest can potentially cause damage to tobacco fields if its population reaches its peak [
17]. Therefore, many studies have focused on developing biopesticides as a component of integrated pest management (IPM) against insect pests, to minimize damage and increase the productivity of crops [
18,
19,
20,
21,
22,
23]. However, continuous applications of synthetic insecticides by farmers to control
S. frugiperda have become a common practice, which is not only uneconomical but also harmful to the environment and natural enemies [
24,
25] and consequently leads to the development of pest resistance. Some pesticides, e.g., methomyl, endosulfan, methyl parathion, and lindane, have been used for
S. frugiperda management and are known to be highly hazardous [
26,
27], and endosulfan and lindane are banned in many countries [
28]. Moreover, pesticides are not safe for farmers due to their toxicity, and the majority of farmers have little information or knowledge on the precautionary measures to take during the application of these insecticides in the field. Hence, there is a strong need to develop alternative management methods to synthetic insecticides that are safe, environmentally friendly, and economical, to sustainably control
S. frugiperda.
Entomopathogenic fungi (EPFs) have been recognized as comprehensive biopesticides in the management of many destructive pests, and could also be integrated as a key component of IPM strategies for FAW control [
29,
30] as they were found to be an efficient control practice against a variety of insect pests [
31,
32]. One of the perceived disadvantages of EPFs is that they do not cause quick mortality (killing speed) to insects, similar to their counterpart synthetic insecticides, due to their different modes of action, therefore insects continue to feed and cause damage to crops [
33,
34]. However, it is important to note that these EPFs help to minimize damage to crops (below the economic threshold) by inducing infection of host pests, which ultimately leads to a reduction in feeding, oviposition, development, mating, and other physiological traits of insects [
35]. A recent study revealed that some
Metarhizium anisopliae isolates showed significant mortality to
S. frugiperda early immature stages [
36].
However, there is little evidence on the efficiency of locally available EPFs in China that are associated with eggs and can affect the feeding performance of S. frugiperda larvae. Hence, the objective of this work was to find effective fungal isolates that can disrupt the eggs and feeding of S. frugiperda, to develop microbial-based biopesticides for use against this notorious invasive lepidopteran pest in China and abroad.
4. Discussion
Numerous pathogens, including viruses, fungi, protozoa, nematodes, and bacteria, have been associated with
S. frugiperda [
44], but only a few cause disease to the pest [
45,
46,
47,
48,
49,
50,
51].
Spodoptera frugiperda nuclear polyhedrosis virus (NPV) is the most important among these pathogens and induces significant mortality to the pest and the entomopathogenic fungal species
Erynia radicans,
Nomuraea rileyi, and
Entomophaga aulicae. Disease or natural epizooty mostly appear too late in the pest population to alleviate high levels of defoliation, regardless of causing microbial agents. Thus, any entomopathogens that might cause high infection to immature stages (eggs and neonate larvae) of
S. frugiperda, before it reaches its voracious feeding stage, could significantly suppress the pest populations. Therefore, the present study focused on screening selected entomopathogenic fungal isolates that could effectively induce high infections to immature stages of
S. frugiperda, as a prerequisite for developing biopesticides to use against the pest.
The results indicated that first to sixth instar
S. frugiperda larvae were not susceptible to the tested entomopathogenic fungal isolates at different concentrations. Previous studies reported that even though some fungal isolates, e.g.,
Aspergillus sp., were found to be least virulent,
B. bassiana was the most virulent to the fourth instar striped rice stem borer larvae,
Chilo suppressalis (Walker) (Lepidoptera: Crambidae) [
52], while another study reported that
B. bassiana caused 12.99% larval mortality to
S. frugiperda [
53]. Similarly, although
P. citrinum did not cause significant larval mortality to
S. frugiperda, it was identified as the most toxic isolate, causing 98.67% mortality to the second instar cotton leafworm larvae,
Spodoptera litura (Fabricius) (Lepidoptera: Noctuidae) [
54].
Penicillium citrinum strains are also known to be associated with mosquito larvae [
55], causing 100% mortality to the third instar southern house mosquito larvae,
Culex quinquefasciatus (Say) (Diptera: Culicidae), within 2 h, using a conidial suspension at a concentration of 1
× 10
6 conidia mL
−1 in a laboratory experiment [
56].
Beauveria bassiana isolates caused 3.3–88.5% mortality to second instar larvae of the spotted stem borer,
Chilo partellus (Swinhoe) (Lepidoptera: Pyralidae), while 30–84.4% mortality was obtained against fifth and sixth instar larvae of the maize stalk borer,
Busseola fusca (Fuller) (Lepidoptera: Noctuidae) [
57]. In previous studies, some fungal isolates showed high virulence to first instar larvae, while low virulence to later instar larvae was observed; for instance,
Cladosporium sp. caused 54% mortality in first instar larvae but was least pathogenic to later instar of the corn earworm larvae,
Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae) [
58]. Few species in the genus
Cladosporium have been reported to be pathogenic against homopteran insects such as aphids [
59] and whiteflies [
60]. Second instar
S. frugiperda larvae were also reported to be less susceptible to the
B. bassiana ICIPE 676 isolate, which caused moderate larval mortality of 30% [
36]. However,
B. bassiana isolates from soil caused 98.3% mortality to third instar
S. frugiperda larvae, while fungi of the same strain, isolated from endophytically colonized maize plants, caused 75% mortality to third instar
S. frugiperda larvae [
61]. Variations in the effectiveness of
B. bassiana against different instars of
S. frugiperda larvae have also been reported [
62]. In contrast,
S. frugiperda was found to be the least susceptible of the most extensively tested insects to
B. bassiana isolates when compared with other lepidopteran insects [
63]. Hence, further studies are warranted to elucidate the mechanisms behind this low susceptibility of
S. frugiperda larvae to the tested fungal isolates in this study. Interestingly, in contrast to the low infection rates observed, the
B. bassiana strain caused 96.6% mortality to second instar
S. frugiperda larvae at 1 × 10
9 conidia ml-
1, with a lethal time of 3.6 days [
64]. Ramzi et al. [
65] demonstrated that
C. suppressalis larvae were highly susceptible to the two commercial
B. bassiana isolates, BB1 and BB2. Resistance to fungal infection as the larvae mature may also be attributed to the composition of the larval integument that allowed for effective penetration of the fungus, resulting in increased mortality in early larval instars [
66]. Meekes [
67] reported that molting may be the reason for loss of inoculum and hence lower chances of fungal infection, although molting does not always result in the prevention of fungal infection. Furthermore, the low pathogenicity of fungal isolates could also be attributed to host–pathogen interactions between these isolates and
S. frugiperda larvae, such as efficient attachment of conidia to the integument, negative impacts of integument composition with penetration tube of the fungi, and immune responses of the larvae toward conidia [
52]. All these phenomena require detailed experiments to precisely elucidate the mechanism.
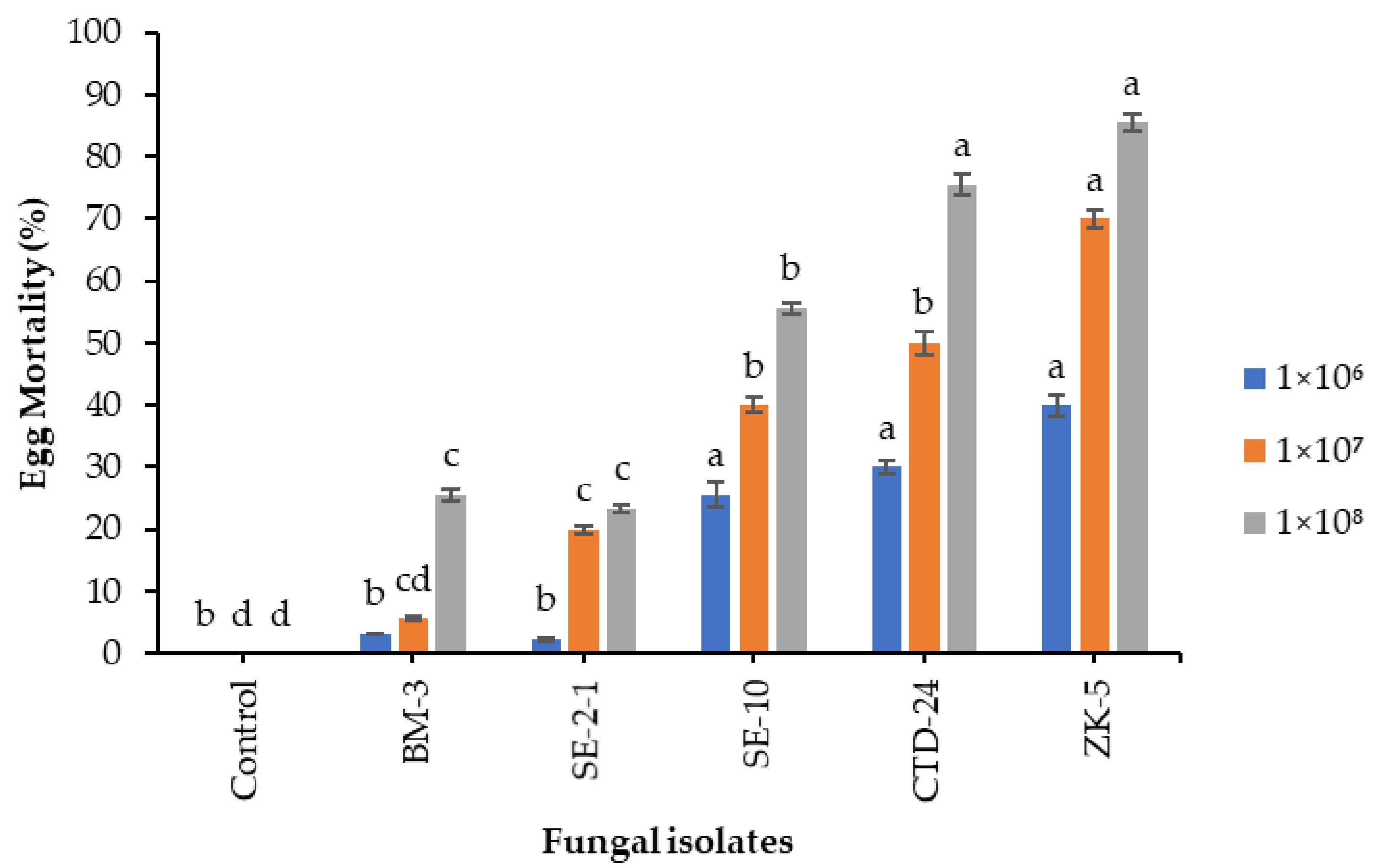
Insects are especially vulnerable to microbial infection during the egg development stage due to their immobility [
68]. Insect eggs require great amounts of nutrients for their development until hatching; therefore, eggs are the most targetable immature stage by pathogenic microorganisms [
69,
70]. The results of the present study showed that the eggs of
S. frugiperda were highly susceptible to the tested entomopathogenic fungal isolates. In line with the present study,
B. bassiana isolates Bb39, Bb23, Bb9, Bb40, Bb19, and Bb21 showed 92, 89.2, 87.6, 82.8, 58, and 38% FAW egg mortality, respectively, at 1 × 10
8 conidia mL
−1 [
71].
Beauveria bassiana isolates BbSA-1, BbSA-2, and BbSA-3 caused 100% egg mortality to the red palm weevil,
Rhynchophorus ferrugineus (Olivier) (Coleoptera: Curculionidae) [
72]. The eggs of
S. fruigiperda were moderately susceptible to
B. bassiana isolates ICIPE 621 and ICIPE 35, causing 51 and 29.5% egg mortalities, respectively [
36]. Al-Kherb [
73] also reported ovicidal activity of
B. bassiana to the beet armyworm,
Spodoptera exigua (Hübner) (Lepidoptera: Noctuidae), causing 50% egg mortality. Bahar et al. [
58] reported that
Cladosporium isolate RM16 caused 64% egg mortality in
Helicoverpa armigera (Hübner) (Lepidoptera: Noctuidae). Anand and Tiwary [
74] reported 100% mortality of unscaled eggs of
S. litura due to infection with
Aspergillus sp., while
P. citrinum caused 2.5% egg mortality to congregating fireflies,
Pteroptyx bearni (Coleoptera: Lampyridae) [
75]. Virulence associated with
B. bassiana to the egg stage has been previously reported for other lepidopteran insects, such as 89–100% egg mortality of the bean pod borer,
Maruca vitrata (Fabricius) (Lepidoptera: Crambidae) [
76], 27.4–96.9% egg mortality of the coffee leafminer,
Perileucoptera coffeella (Guérin-Meneville) (Lepidoptera: Lyonetiidae) [
77], and 63% egg mortality of the potato tuber moth,
Phthorimaea operculella (Zeller) (Lepidoptera: Gelechiidae) [
78]. Histopathological studies on the infection of insect eggs by EPFs revealed successful adhesion, germination, and penetration into eggs within 24 h postinoculation in the red spider mite,
Tetranychus urticae (Koch) [
79] and within 6 h postinoculation in the tomato leafminer,
Tuta absoluta (povolny) [
80], and intense extrusion of mycelium covering the eggs was observed in
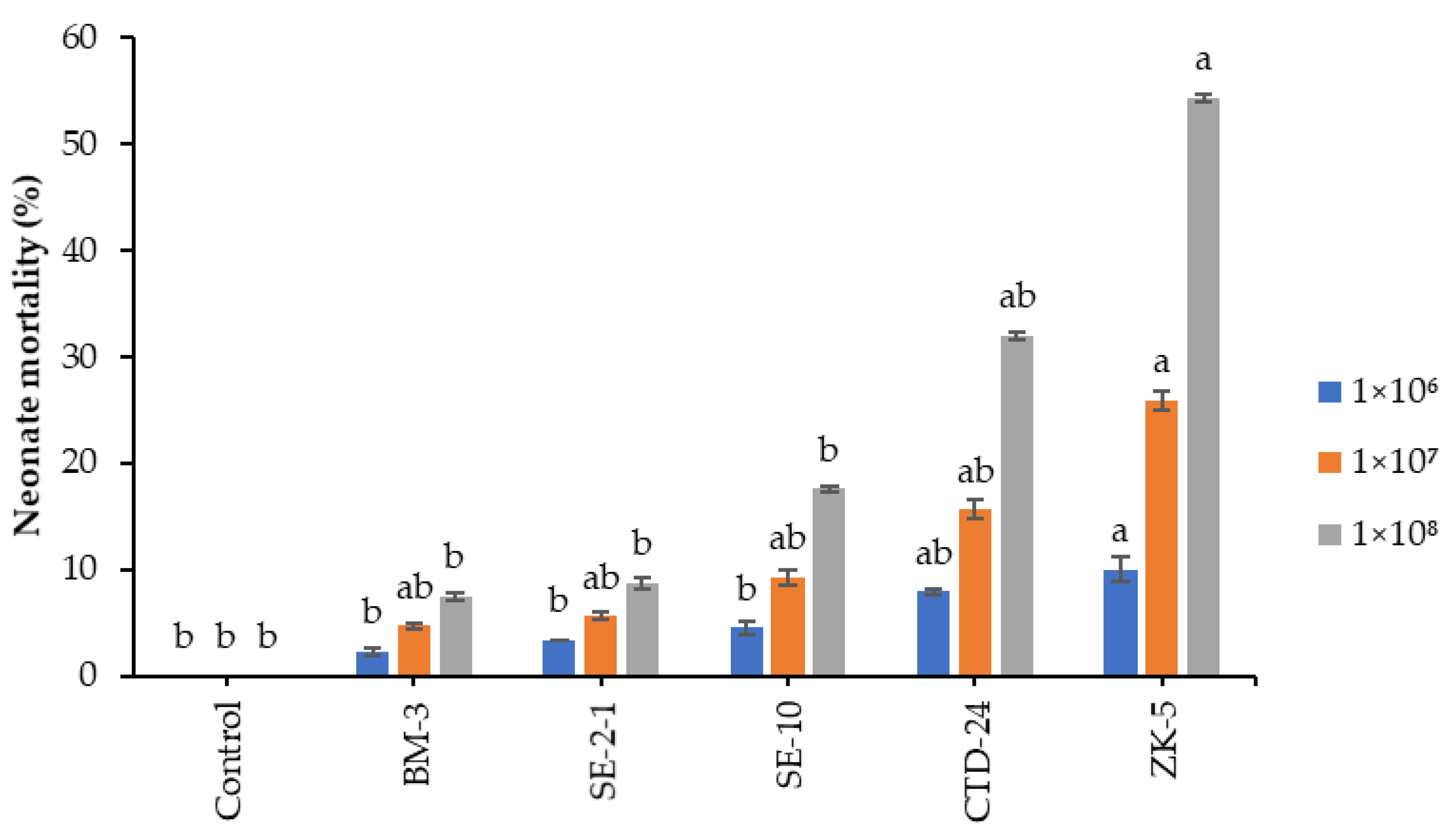
T. absolute 72 h postinoculation. In a previous report, some fungal isolates were highly virulent to neonate larvae; for example,
B. bassiana isolates caused 53.9 to 83.9% neonate mortality to
S. fruigiperda [
36]. Entomopathogenic fungi can infect insects at any stage of their life, although not all host stages are equally susceptible to pathogen infection [
38].
Beauveria bassiana ZK-5 and
P. citrinum CTD-24 caused the highest cumulative mortality to eggs and neonates of
S. frugiperda in the present study. Our findings are consistent with those of Akutse et al. [
36], who found that
B. bassiana ICIPE 621 caused a cumulative mortality of 87.5% to eggs and neonates of
S. frugiperda. Gui et al. [
59] found that
Cladosporium aphidis fungal isolates were highly virulent to various aphid species, causing a cumulative mortality of 80% within 8 days at 1 × 10
8 spores mL
−1.
The considerable reduction in food consumption by insects has been attributed to the production of toxic substances by EPFs inside the host that lead to mechanical disruption in insect structural integrity [
81].
Beauveria bassiana reduced the food consumption of the Mediterranean corn stalk borer,
Sesamia nonagrioides (Lepidoptera: Noctuidae), by 50% at the highest conidial concentrations [
82]. In previous studies, a reduction in feeding due to infection of
B. bassiana fungus from other insect species has also been reported, such as in the Colorado potato beetle,
Leptinotarsa decemlineata (Say) (Coleoptera: Chrysomelidae), the bean flower thrip,
Megalurothrips sjostedti (Trybom) (Thysanoptera: Thripidae), the spotted stem borer,
C. partellus (Swinhoe) (Lepidoptera: Crambidae), the greenish silk-moth,
Ocinara varians (Walker) (Lepidoptera: Bombycidae), and the pea leaf miner,
Liriomyza huidobrensis (LIRIHU) (Diptera: Agromyzidae) [
34,
83,
84,
85]. Tefera and Pringle [
81] observed that
B. bassiana significantly reduced the food consumption of
C. partellus larvae.
In contrast, Cheung and Grula [
86] reported no significant reduction in feeding for the corn earworm larvae,
Helicoverpa zea, infected with
B. bassiana before death. The reduction in food consumption by infected insects due to fungi is one of the important factors in host mortality, which indicates fungal pathogen virulence and needs to be further investigated to determine the level of pathogenicity or antifeedant effects [
87].
Beauveria bassiana caused a 60% reduction in food consumption to the long-horned grasshoppers,
Uvarovistia zebra (Uvarov) (Orthoptera: Tettigoniidae), at a concentration of 5 × 10
6 spores mL
−1 [
88].


 ,
, 




{kind=link}
{kind=link}