
Effect of Hypoxia on the Lethal Mortality Time of Adult Sitophilus oryzae L.




Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Rearing
2.2. Hypoxia Treatment
2.3. Experimental Setup and Design
2.4. Data Collection
2.4.1. Adult Mortality
2.4.2. Adult Emergence
2.4.3. Temperature and RH
2.5. Statistical Analysis
3. Results
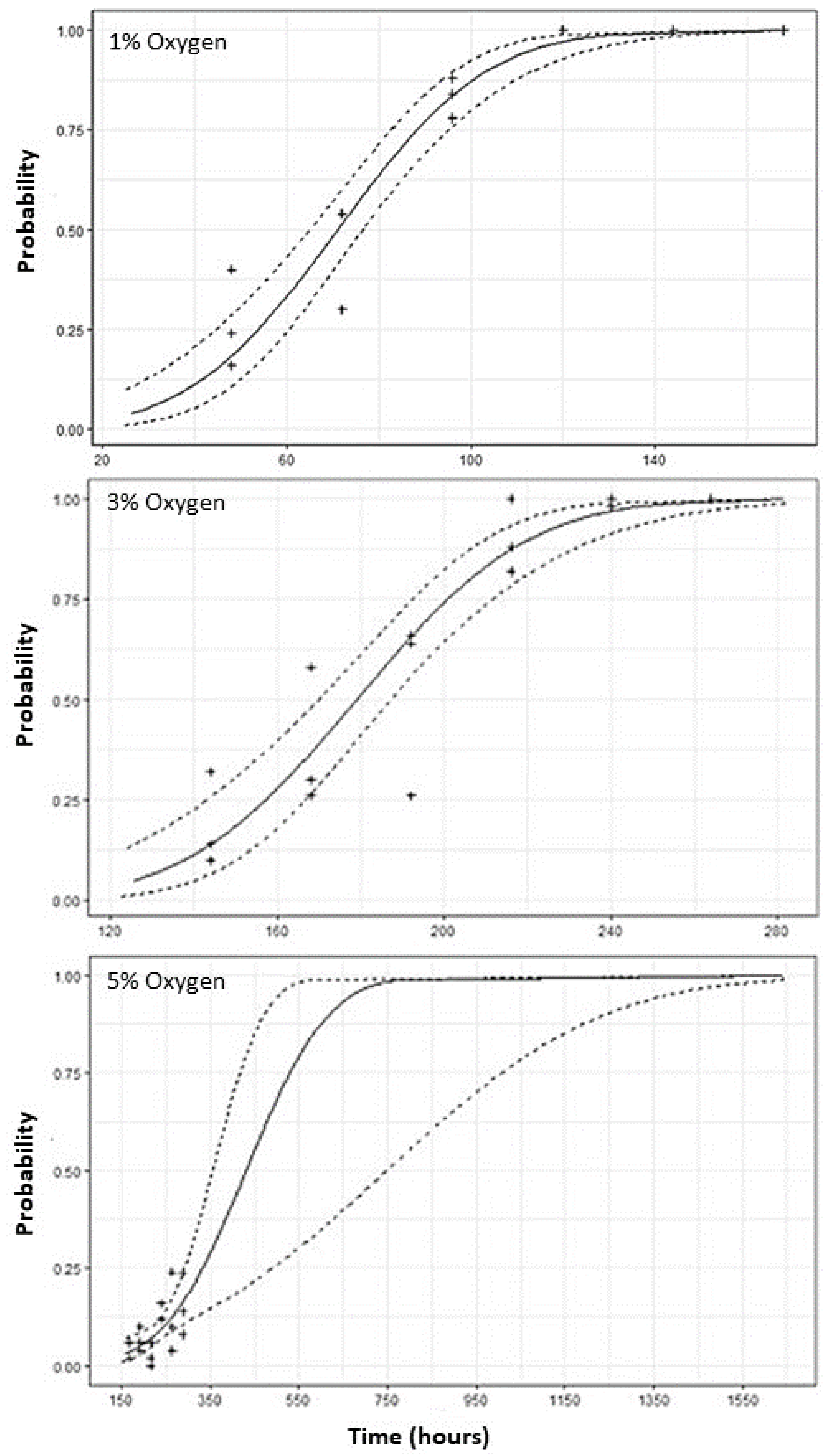
3.1. Lethal Time
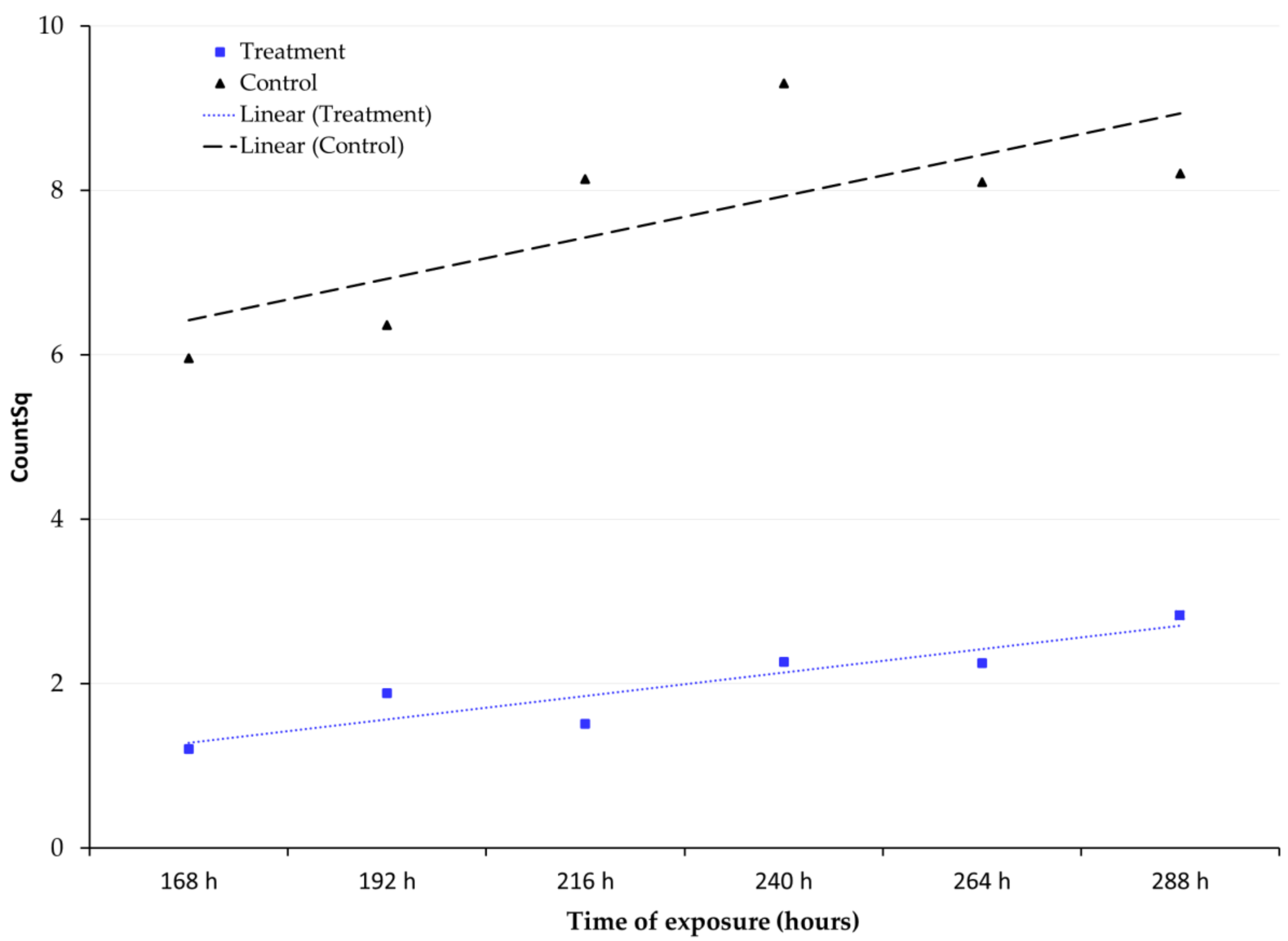
3.2. Adult Emergence
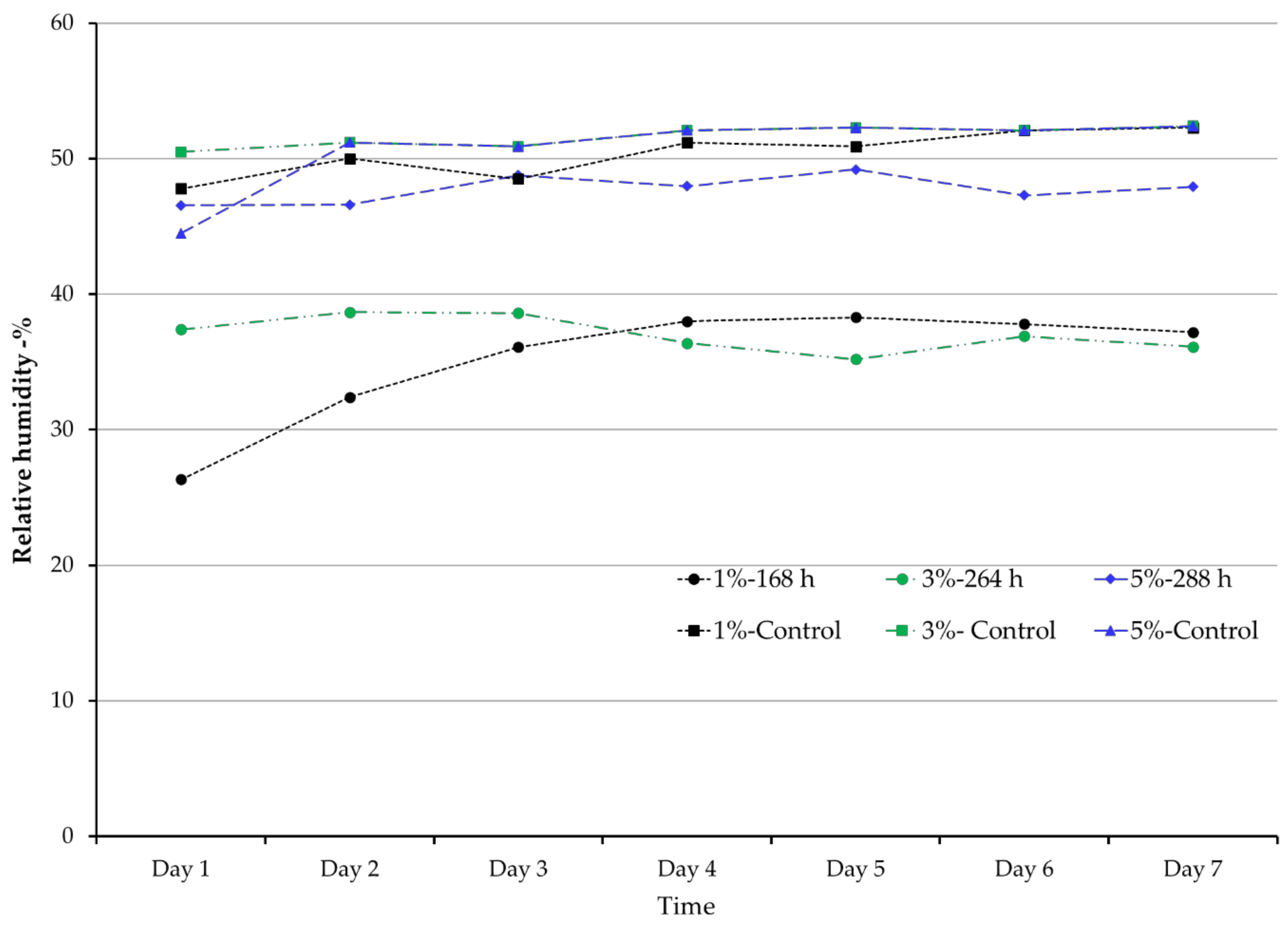
3.3. Temperature and RH
4. Discussion
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Martin, D.T.; Baributsa, D.; Huesing, J.E.; Williams, S.B.; Murdock, L.L. PICS bags protect wheat grain, Triticum aestivum (L.), against rice weevil, Sitophilus oryzae (L.) (Coleoptera: Curculionidae). J. Stored Prod. Res. 2015, 63, 22–30. [Google Scholar] [CrossRef]
- Mason, L.J.; McDonough, M. Biology, Behavior, and Ecology of Stored Grain and Legume Insects. In Stored Product Protection; Hagstrum, D.W., Phillips, T.W., Cuperus, G., Eds.; Kansas State University: Manhattan, KS, USA, 2012; pp. 7–20. ISBN 978-0-98550-030-6. [Google Scholar]
- Longstaff, B.C. Biology of the grain pest species of the genus Sitophilus (Coleoptera: Curculionidae): A critical review. Prot. Ecol. 1981, 3, 83–130. [Google Scholar]
- Baoua, I.B.; Amadou, L.; Bakoye, O.; Baributsa, D.; Murdock, L.L. Triple bagging hermetic technology for post-harvest preservation of paddy rice Oryza sativa L. in the Sahel of West Africa. J. Stored Prod. Res. 2016, 68, 73–79. [Google Scholar] [CrossRef]
- Kandel, P.; Kharel, K.; Njoroge, A.; Smith, B.W.; Díaz-Valderrama, J.R.; Timilsina, R.H.; Paudel, G.P.; Baributsa, D. On-Farm Grain Storage and Challenges in Bagmati Province, Nepal. Sustainability 2021, 13, 7959. [Google Scholar] [CrossRef]
- Kumar, D.; Kalita, P. Reducing Postharvest Losses during Storage of Grain Crops to Strengthen Food Security in Developing Countries. Foods 2017, 6, 8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Afful, E.; Elliott, B.; Nayak, M.K.; Phillips, T.W. Phosphine Resistance in North American Field Populations of the Lesser Grain Borer, Rhyzopertha Dominica (Coleoptera: Bostrichidae). J. Econ. Entomol. 2018, 111, 463–469. [Google Scholar] [CrossRef] [PubMed]
- Chen, Z.; Schlipalius, D.; Opit, G.; Subramanyam, B.; Phillips, T.W. Diagnostic molecular markers for phosphine resistance in U.S. populations of Tribolium castaneum and Rhyzopertha dominica. PLoS ONE 2015, 10, e0121343. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Porter, I.; Fraser, P. Progress and remaining challenges with the phase-out of methyl bromide under the Montreal Protocol. Acta Hortic. 2020, 1270. [Google Scholar] [CrossRef]
- Fleurat-Lessard, F. Effect of modified atmospheres on insect and mites infesting stored products. In Food Preservation by Modified Atmospheres; Calderon, M., Barkai-Golan, R., Eds.; CRC Press: Boca Raton, FL, USA, 1990; pp. 21–38. [Google Scholar]
- Navarro, S. The effects of low oxygen tensions on three stored-product insect pests. Phytoparasitica 1978, 6, 51–58. [Google Scholar] [CrossRef]
- Adler, C.; Corinth, H.; Reichmuth, C. Modified atmospheres. In Alternatives to Pesticides in Stored Product IPM; Subramanyam, B., Hagstrum, D., Eds.; Kluwer Academic Publishers: Boston, MA, USA, 2000; pp. 105–146. ISBN 978-8-57811-079-6. [Google Scholar]
- Navarro, S. Modified Atmospheres for the Control of Stored-Product Insects and Mites. In Insect Management for Food Storage and Processing; Elsevier: Amsterdam, The Netherlands, 2006; pp. 105–145. ISBN 978-0-12810-429-3. [Google Scholar]
- Bailey, S. Air-tight storage of grain; its effects on insect pests. I. Calandra granaria L. (Coleoptera, Curculionidae). Aust. J. Agric. Res. 1955, 6, 33. [Google Scholar] [CrossRef]
- Banks, H.; PC, A. Comparative advantages of high CO2 and low O2 types of controlled atmospheres for grain storage. In Food Preservation by Modified Atmospheres; Calderon, M., Barkai-Golan, R., Eds.; CRC Press: Boca Raton, FL, USA, 1990; pp. 93–122. [Google Scholar]
- Brandl, D.G.; Soderstrom, E.L.; Schreiber, F.E. Effects of Low-Oxygen Atmospheres Containing Different Concentrations of Carbon Dioxide on Mortality of the Navel Orangeworm, Amyelois transitella Walker (Lepidoptera: Pyralidae). J. Econ. Entomol. 1983, 76, 828–830. [Google Scholar] [CrossRef]
- Harein, P.K.; Press, A.F. Mortality of stored-peanut insects exposed to mixtures of atmospheric gases at various temperatures. J. Stored Prod. Res. 1968, 4, 77–82. [Google Scholar] [CrossRef]
- Njoroge, A.W.; Mankin, R.W.; Smith, B.; Baributsa, D. Effects of Hypoxia on Acoustic Activity of Two Stored-Product Pests, Adult Emergence, and Grain Quality. J. Econ. Entomol. 2019, 112, 1989–1996. [Google Scholar] [CrossRef] [PubMed]
- Njoroge, A.W.; Mankin, R.W.; Smith, B.W.; Baributsa, D. Effects of Hermetic Storage on Adult Sitophilus oryzae L. (Coleoptera: Curculionidae) Acoustic Activity Patterns and Mortality. J. Econ. Entomol. 2017, 110, 2707–2715. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kharel, K.; Mason, L.J.J.; Murdock, L.L.L.; Baributsa, D. Efficacy of Hypoxia against Tribolium castaneum (Coleoptera: Tenebrionidae) throughout Ontogeny. J. Econ. Entomol. 2019, 112, 1463–1468. [Google Scholar] [CrossRef] [PubMed]
- Singano, C.D.; Mvumi, B.M.; Stathers, T.E. Effectiveness of grain storage facilities and protectants in controlling stored-maize insect pests in a climate-risk prone area of Shire Valley, Southern Malawi. J. Stored Prod. Res. 2019, 83, 130–147. [Google Scholar] [CrossRef]
- Mankin, R.W.; Mizrach, A.; Hetzroni, A.; Levsky, S.; Nakache, Y.; Soroker, V. Temporal and spectral features of sounds of wood-boring beetle larvae: Identifiable patterns of activity enable improved discrimination from background noise. BioOne 2008, 91, 241–248. [Google Scholar] [CrossRef]
- Johnson, P. Residuals and Analysis of Fit. 2016. Available online: http://pj.freefaculty.org/stat/GLM/GLM2_v2.pdf (accessed on 15 April 2021).
- Hlina, B.L. Analysis of Ecotoxicology 2020; Package ‘Ecotox’. Available online: https://cran.r-project.org/web/packages/ecotox/ecotox.pdf (accessed on 15 April 2021).
- Navarro, S. The use of modified and controlled atmospheres for the disinfestation of stored products. J. Pest. Sci. 2012, 85, 301–322. [Google Scholar] [CrossRef]
- Baoua, I.B.; Amadou, L.; Lowenberg-DeBoer, J.D.; Murdock, L.L. Side by side comparison of GrainPro and PICS bags for postharvest preservation of cowpea grain in Niger. J. Stored Prod. Res. 2013, 54, 13–16. [Google Scholar] [CrossRef]
- Yan, Y.; Williams, S.; Baributsa, D.; Murdock, L. Hypoxia Treatment of Callosobruchus maculatus Females and Its Effects on Reproductive Output and Development of Progeny Following Exposure. Insects 2016, 7, 26. [Google Scholar] [CrossRef] [PubMed] [Green Version]




{kind=link}
{kind=link}
{kind=link}
| 1% | Time | 48 h | 72 h | 96 h | 120 h | 144 h | 168 h | Control |
| Mortality (%) | 20.33a | 46.67b | 89.33c | 100c | 100c | 100c | 2.83a | |
| 3% | Time | 144 h | 168 h | 192 h | 216 h | 240 h | 264 h | Control |
| Mortality (%) | 15.67ab | 30.33bc | 42.67c | 75.33d | 97.67e | 100e | 1.17a | |
| 5% | Time | 168 h | 192 h | 216 h | 240 h | 264 h | 288 h | Control |
| Mortality (%) | 4.33a | 7.67a | 8.67ab | 14.67bc | 16.33bc | 21.00c | 1.00c |
| % Oxygen | LT_Value a | N | Mortality (h) | LCL b | UCL c | χ2 d | DF e | Slope |
|---|---|---|---|---|---|---|---|---|
| 1% | 25 | 1600 | 54.9 | 49.4 | 59.3 | 102.8 | 34 | 0.046 |
| 50 | 1600 | 69.7 | 65.9 | 73.3 | ||||
| 99 | 1600 | 120.6 | 112.9 | 131.3 | ||||
| 3% | 25 | 1600 | 164.2 | 155.3 | 171.3 | 169.5 | 34 | 0.029 |
| 50 | 1600 | 187.8 | 181.6 | 193.7 | ||||
| 99 | 1600 | 269.0 | 256.1 | 287.2 | ||||
| 5% | 25 | 1600 | 298.3 | 279.3 | 333.2 | 49.7 | 34 | 0.0076 |
| 50 | 1600 | 386.6 | 352.2 | 474.8 | ||||
| 99 | 1600 | 691.4 | 575.8 | 925.6 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kandel, P.; Scharf, M.E.; Mason, L.J.; Baributsa, D. Effect of Hypoxia on the Lethal Mortality Time of Adult Sitophilus oryzae L. Insects 2021, 12, 952. https://doi.org/10.3390/insects12100952
Kandel P, Scharf ME, Mason LJ, Baributsa D. Effect of Hypoxia on the Lethal Mortality Time of Adult Sitophilus oryzae L. Insects. 2021; 12(10):952. https://doi.org/10.3390/insects12100952
Chicago/Turabian StyleKandel, Pragya, Michael E. Scharf, Linda J. Mason, and Dieudonne Baributsa. 2021. "Effect of Hypoxia on the Lethal Mortality Time of Adult Sitophilus oryzae L." Insects 12, no. 10: 952. https://doi.org/10.3390/insects12100952