The Population Growth of Spodoptera frugiperda on Six Cash Crop Species and Implications for Its Occurrence and Damage Potential in China

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Insect Culture
2.2. Plant Species
2.3. Life Table Study of S. frugiperda
2.4. Life Table Data Analysis
2.5. Population Projection of S. frugiperda
2.6. Statistical Analysis
3. Results
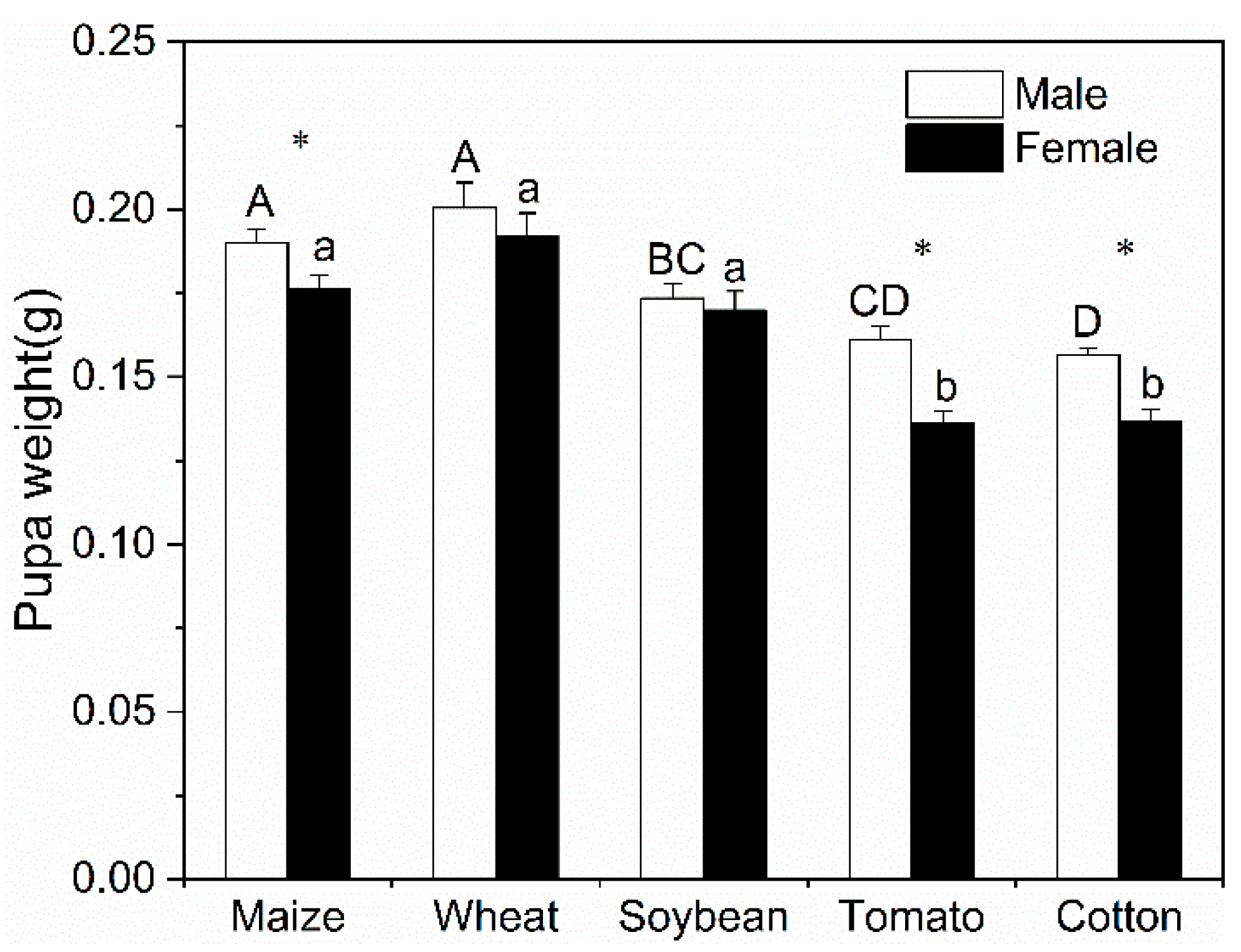
3.1. Life History of S. frugiperda
3.2. Population Parameters of S. frugiperda
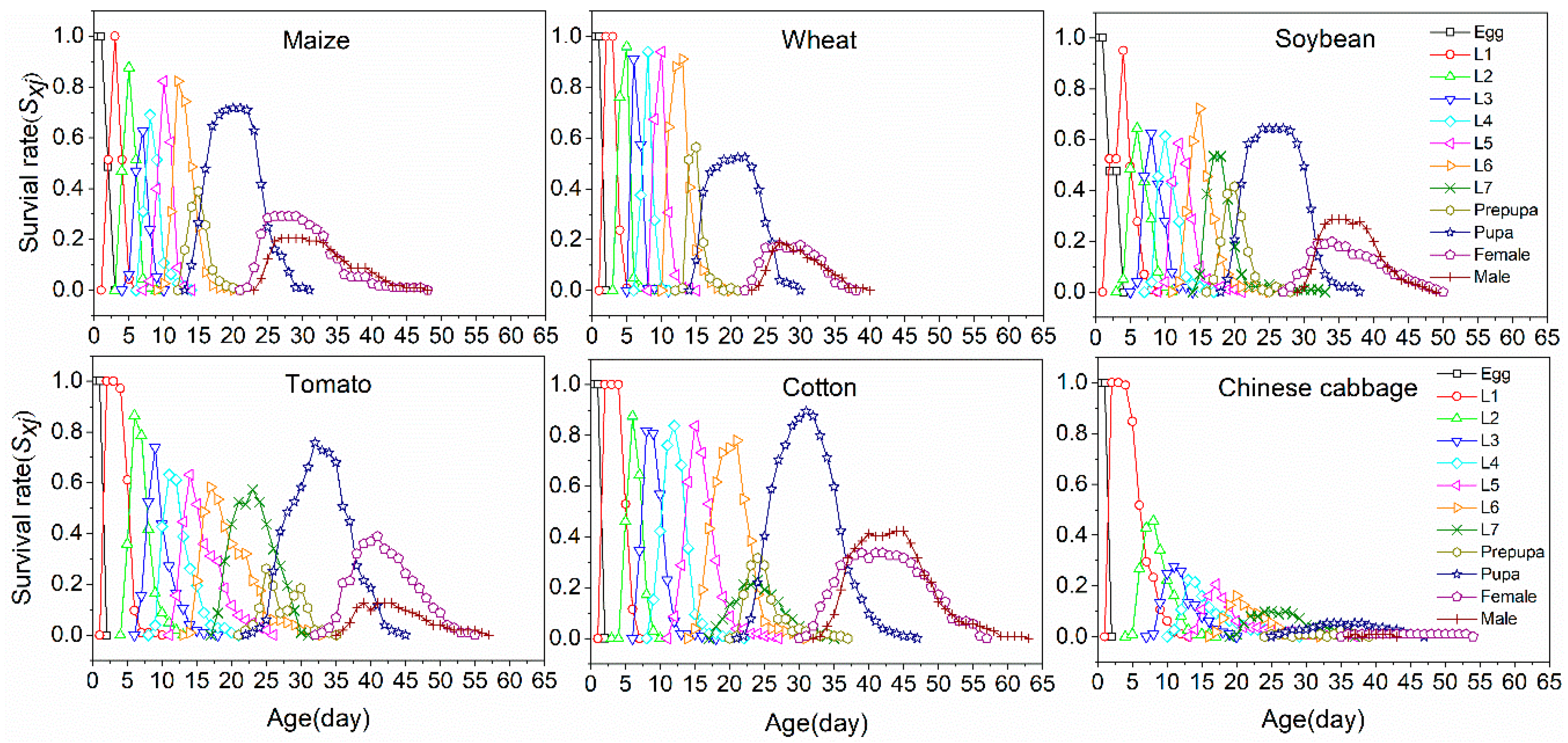
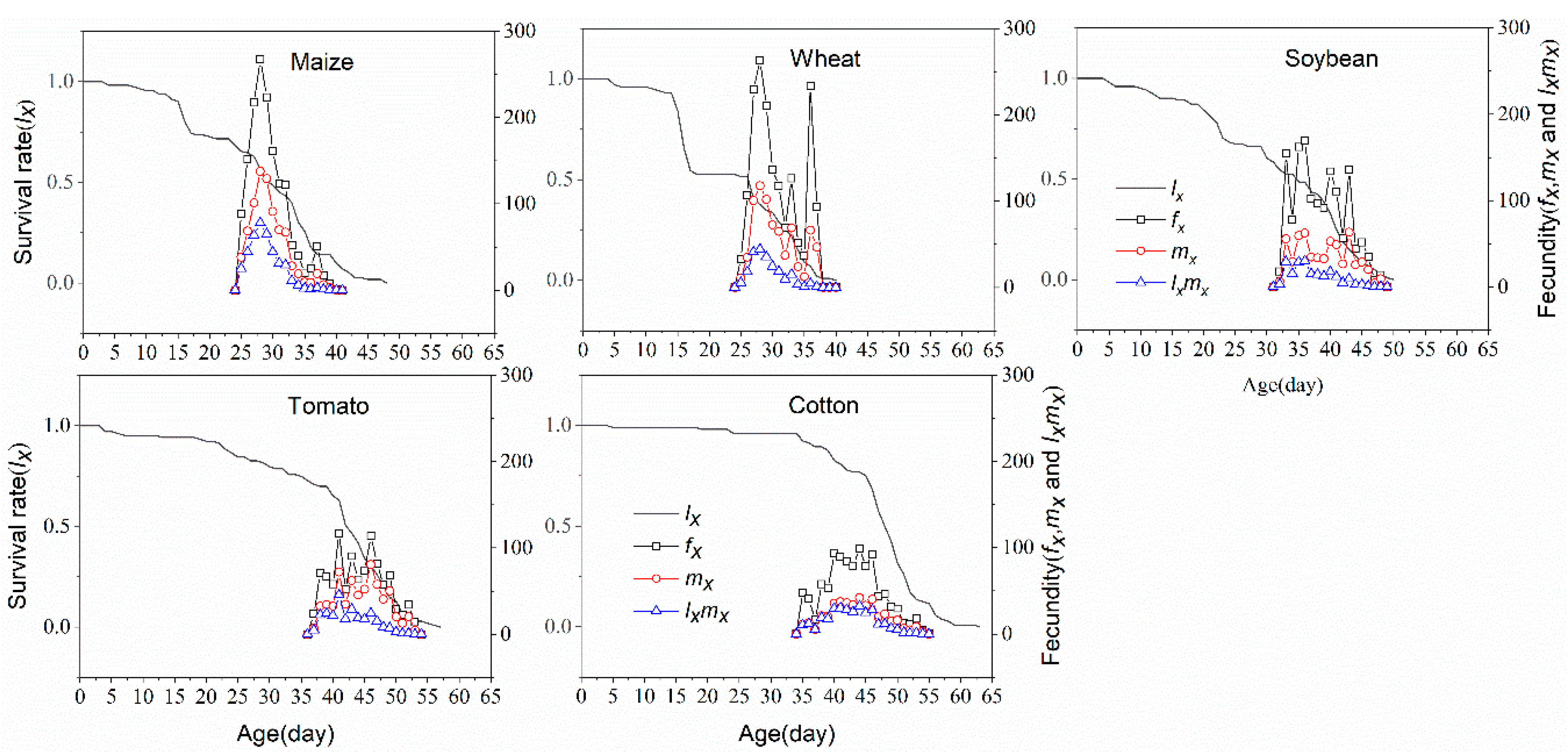
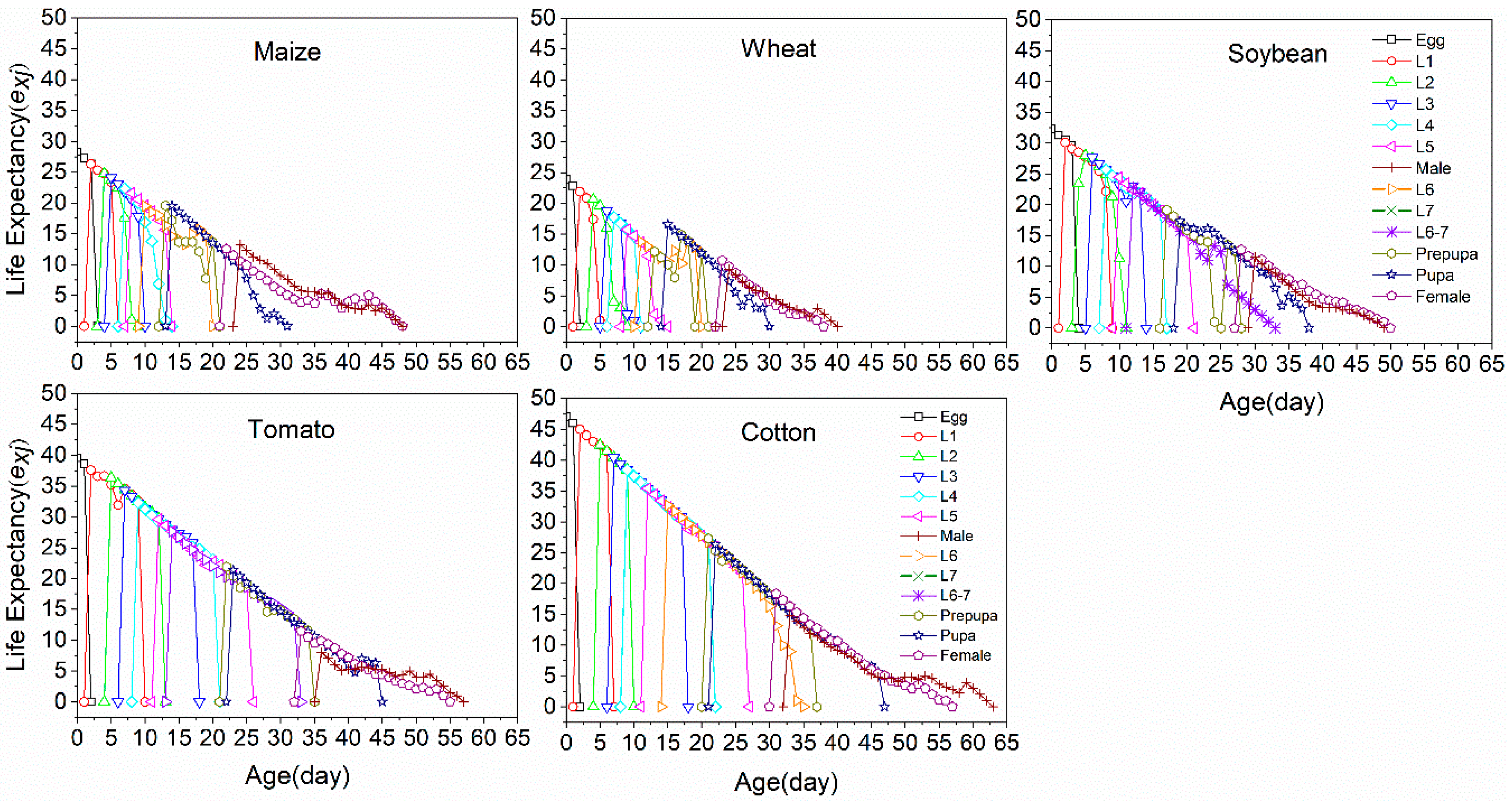
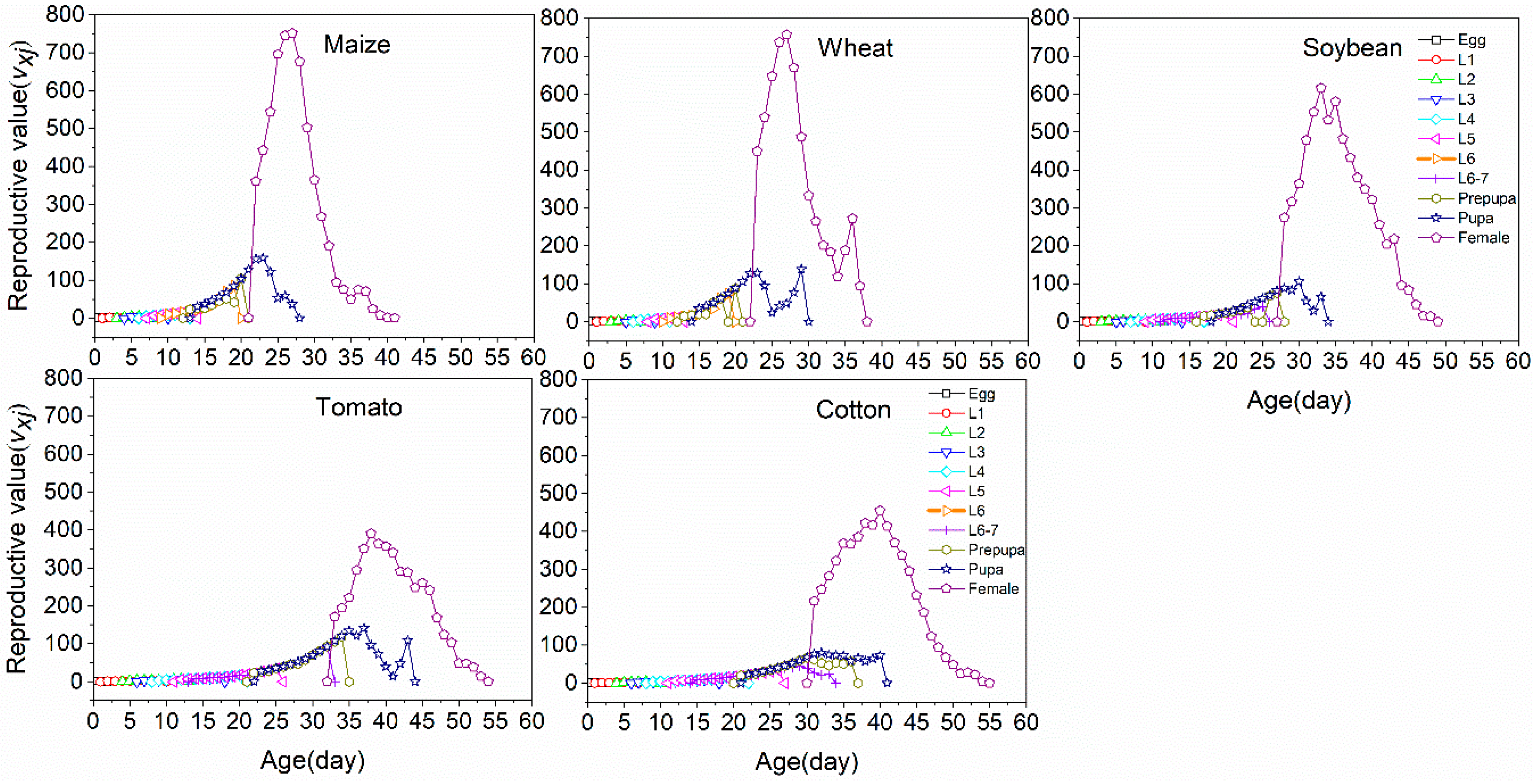
3.3. Life Table Analysis
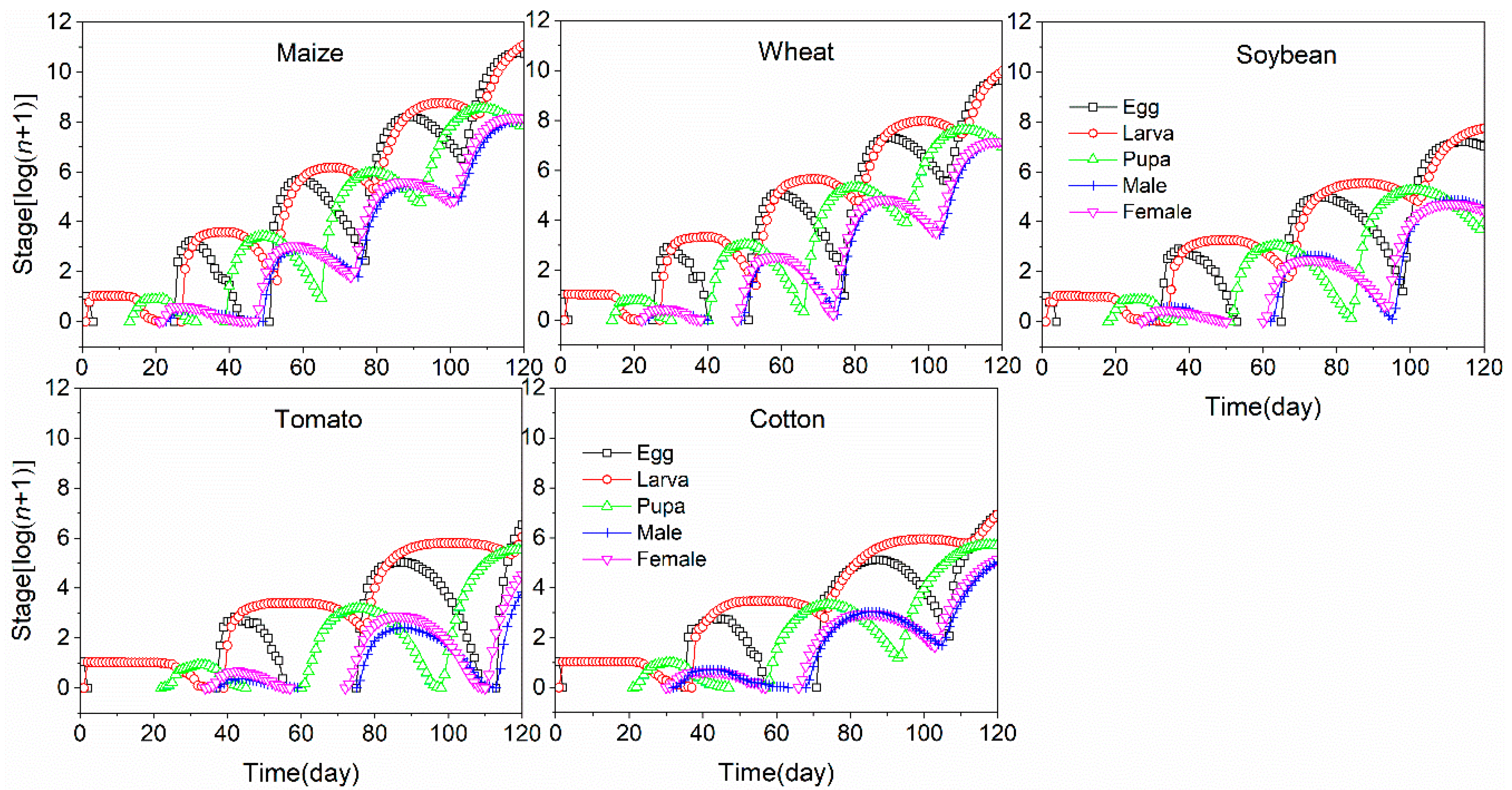
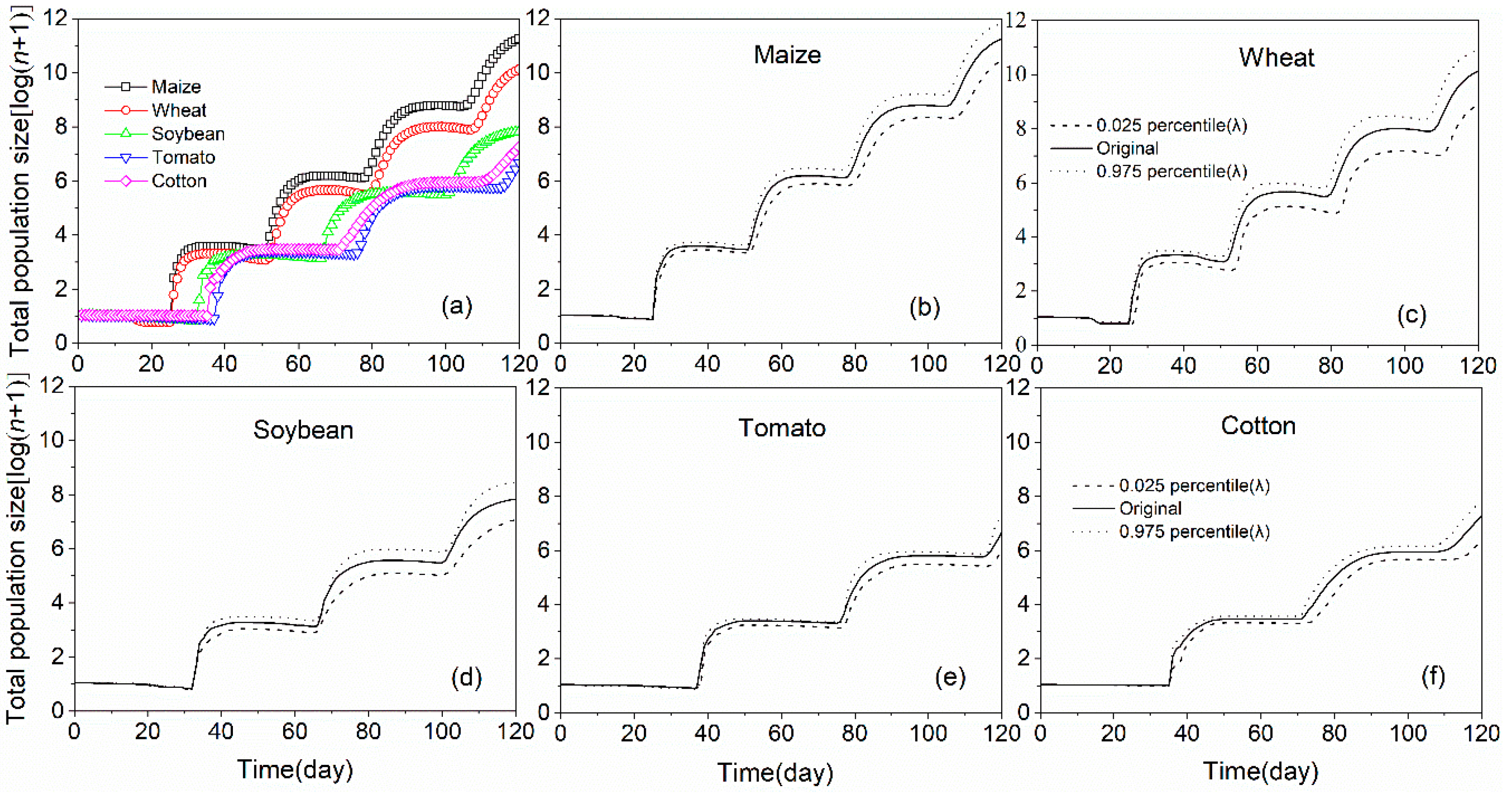
3.4. Population Projection of S. frugiperda
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Johnson, S.J. Migration and the life history strategy of the fall armyworm, Spodoptera frugiperda in the western hemisphere. Int. J. Trop. Insect Sci. 1987, 8, 543–549. [Google Scholar] [CrossRef]
- Montezano, D.G.; Specht, A.; Sosa-Gómez, D.R.; Roque-Specht, V.F.; Sousa-Silva, J.C.; Paula-Moraes, S.V.; Peterson, J.A.; Hunt, T.E. Host Plants of Spodoptera frugiperda (Lepidoptera: Noctuidae) in the Americas. Afr. Entomol. 2018, 26, 286–300. [Google Scholar] [CrossRef] [Green Version]
- Westbrook, J.; Nagoshi, R.; Meagher, R.; Fleischer, S.; Jairam, S. Modeling seasonal migration of fall armyworm moths. Int. J. Biometeorol. 2016, 60, 255–267. [Google Scholar] [CrossRef] [PubMed]
- Dumas, P.; Legeai, F.; Lemaitre, C.; Scaon, E.; Orsucci, M.; Labadie, K.; Gimenez, S.; Clamens, A.L.; Henri, H.; Vavre, F.; et al. Spodoptera frugiperda (Lepidoptera: Noctuidae) host-plant variants: Two host strains or two distinct species? Genetica 2015, 143, 305–316. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deshmukh, S.; Kalleshwaraswamy, C.M.; Asokan, R.; Swamy, H.M.M.; Maruthi, M.S.; Pavithra, B.P.; Hegde, K.; Navi, S.; Prabhu, S.T.; Goergen, G. First report of the fall armyworm, Spodoptera frugiperda (J. E. Smith) (Lepidoptera: Noctuidae), an alien invasive pest on maize in India. Pest. Manag. Hortic. Ecosyst. 2018, 24, 23–29. [Google Scholar]
- FAO. Global Action for Fall Armyworm Control. Available online: http://www.fao.org/fall-armyworm/faw-management/zh/ (accessed on 17 January 2020).
- Yang, X.L.; Liu, Y.C.; Luo, M.Z.; Li, Y.; Wang, W.H.; Wan, F.; Jiang, H. Spodoptera frugiperda moved into southwestern China for the first time in Jiangcheng County, Yunnan Province. Yunnan Agric. 2019, 1, 72. [Google Scholar]
- Jiang, Y.Y.; Liu, J.; Xie, M.C.; Li, Y.H.; Yang, J.J.; Zhang, M.L.; Qiu, K. Observation on law of diffusion damage of Spodoptera frugiperdain in China in 2019. Plant. Prot. 2019, 45, 10–19. [Google Scholar]
- Wu, K. Management strategies of fall armyworm (Spodoptera frugiperda) in China. Plant. Prot. 2020, 46, 1–5. [Google Scholar]
- Guillebeau, L.P.; All, J.N. Use of pyrethroids, methomyl, and chlorpyrifos to control fall armyworm (Lepidoptera: Noctuidae) in whorl stage field corn, sweet corn and sorghum. Fla. Entomol. 1991, 74, 261–270. [Google Scholar] [CrossRef]
- Burtet, L.M.; Bernardi, O.; Melo, A.A.; Pes, M.P.; Strahl, T.T.; Guedes, J.V.C. Managing fall armyworm, Spodoptera frugiperda (Lepidoptera: Noctuidae), with Bt maize and insecticides in southern Brazil. Pest Manag. Sci. 2017, 73, 2569–2577. [Google Scholar] [CrossRef]
- Farias, J.R.; Andow, D.A.; Horikoshi, R.J.; Sorgatto, R.J.; Fresia, P.; Santos, A.C.; Omoto, C. Field-evolved resistance to Cry1F maize by Spodoptera frugiperda (Lepidoptera: Noctuidae) in Brazil. Crop. Prot. 2014, 64, 150–158. [Google Scholar] [CrossRef]
- Gutiérrez-Moreno, R.; Mota-Sanchez, D.; Blanco, C.A.; Whalon, M.E.; Terán-Santofimio, H.; Rodriguez-Macie, J.C.; DiFonzo, C. Field-evolved resistance of the fall armyworm (Lepidoptera: Noctuidae) to synthetic insecticides in Puerto Rico and Mexico. J. Econ. Entomol. 2019, 112, 792–802. [Google Scholar] [CrossRef] [PubMed]
- He, Q.; Zhou, G. Climatic suitability of potential summer maize planting zones in China. Acta Geogr. Sin. 2011, 66, 1443–1450. [Google Scholar]
- He, Q.; Zhou, G. Climatic suitability of potential spring maize cultivation distribution in China. Acta Ecol. Sin. 2012, 32, 3931–3939. [Google Scholar]
- Awmack, C.S.; Leather, S.R. Host plant quality and fecundity in herbivorous insects. Annu. Rev. Entomol. 2002, 47, 817–844. [Google Scholar] [CrossRef]
- Chi, H.; Su, H.Y. Age-stage, two-sex life tables of Aphidius gifuensis (Ashmead) (Hymenoptera: Braconidae) and its host Myzus persicae (Sulzer) (Homoptera: Aphididae) with mathematical proof of the relationship between female fecundity and the net reproductive rate. Environ. Entomol. 2006, 35, 10–21. [Google Scholar] [CrossRef]
- Tuan, S.J.; Lee, C.C.; Chi, H. Population and damage projection of Spodoptera litura (F.) on peanuts (Arachis hypogaea L.) under different conditions using the age-stage, two-sex life table. Pest Manag. Sci. 2014, 70, 805–813. [Google Scholar] [CrossRef]
- Qayyum, A.; Aziz, M.A.; Iftikhar, A.; Hafeez, F.; Atlihan, R. Demographic parameters of Lipaphis erysimi (Hemiptera: Aphididae) on different cultivars of Brassica vegetables. J. Econ. Entomol. 2018, 111, 1885–1894. [Google Scholar] [CrossRef]
- Chi, H. TWOSEX-MSChart: A Computer Program for the Age-Stage, Two-Sex Life Table Analysis. 2019. Available online: http://140.120.197.173/Ecology/Download/Twosex-MSChart.zip (accessed on 9 December 2019).
- Chi, H.; Liu, H. Two new methods for the study of insect population ecology. Bull. Inst. Zool. Acad. Sin. 1985, 24, 225–240. [Google Scholar]
- Chi, H. Life-table analysis incorporating both sexes and variable development rates among individuals. Environ. Entomol. 1988, 17, 26–34. [Google Scholar] [CrossRef]
- Chi, H. TIMING-MSChart: A Computer Program for the Age Stage, Two-Sex Life Table Analysis. 2019. Available online: http://140.120.197.173/Ecology/Download/TIMING.zip (accessed on 20 December 2019).
- Tibshirani, R.J.; Efron, B. An Introduction to the Bootstrap; Chapman & Hall: London, UK, 1993; Volume 57, pp. 45–82. [Google Scholar]
- Saeed, R.; Sayyed, A.H.; Shad, S.A.; Zaka, S.M. Effect of different host plants on the fitness of diamond-back moth, Plutella xylostella (Lepidoptera: Plutellidae). Crop. Protect. 2010, 29, 178–182. [Google Scholar] [CrossRef]
- Ba, T.X.; Zhang, Y.H.; Zhang, Z.; Guan, D.D.; Li, C.C.; Ji, Z.Y.; Yin, X.T.; Zhang, A.H.; Tang, Q.B.; Liu, Y.H.; et al. The host preference and population life tables of Spodoptera frugiperda (Lepidoptera: Noctuidae) fed on maize and wheat. Plant Prot. 2020, 46, 17–23. [Google Scholar]
- Barros, E.M.; Torres, J.B.; Bueno, A.F. Oviposition, development, and reproduction of Spodoptera frugiperda (J.E. Smith) (Lepidoptera: Noctuidae) fed on different hosts of economic importance. Neotrop. Entomol. 2010, 39, 996–1001. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Y.Q.; Wang, X.Q.; Zhong, Y.W. Fall armyworm Spodoptera frugiperda feeding on cabbage in Zhejiang. Plant Prot. 2019, 45, 90–91. [Google Scholar]
- Pencoe, N.L.; Martin, P.B. Development and reproduction of fall armyworms on several wild grasses. Environ. Entomol. 1981, 10, 999–1002. [Google Scholar] [CrossRef]
- Pencoe, N.L.; Martin, P.B. Fall armyworm (Lepidoptera: Noctuidae) larval development and adult fecundity on five grass hosts. Environ. Entomol. 1982, 11, 720–723. [Google Scholar] [CrossRef]
- Esperk, T.; Tammaru, T.; Nylin, S. Intraspecific variability in number of larval instars in insects. J. Econ. Entomol. 2007, 100, 627–645. [Google Scholar] [CrossRef]
- Wu, Z.; Shi, P.; Zeng, Y.; Huang, W.; Huang, Q.; Ma, X.; Guo, L. Population life tables of Spodoptera frugiperda (Lepidoptera: Noctuidae) fed on three host plants. Plant Prot. 2019, 45, 59–64. [Google Scholar]
- Taylor, P.; Shields, E. Development of the armyworm (Lepidoptera: Noctuidae) under fluctuating daily temperature regimes. Environ. Entomol. 1990, 19, 1422–1431. [Google Scholar] [CrossRef]
- Chen, C.; Zhou, H.; Xia, Q.; Chen, Y.; Xue, F. Temperature-dependent development and protogyny in Helicoverpa armigera. Chin. J. Appl. Entomol. 2012, 49, 867–873. [Google Scholar]
- Guo, J.W.; Yang, F.; Li, P.; Liu, X.D.; Wu, Q.L.; Hu, G.; Zhai, B.P. Female bias in an immigratory population of Cnaphalocrocis medinalis moths based on field surveys and laboratory tests. Sci. Rep. 2019, 9, 18388. [Google Scholar] [CrossRef] [Green Version]
- Alami, S.; Naseri, B.; Golizadeh, A.; Razmjou, J. Age-stage, two-sex life table of the tomato looper, Chrysodeixis chalcites (Lepidoptera: Noctuidae), on different bean cultivars. Arthropod-Plant Interact. 2014, 8, 475–484. [Google Scholar] [CrossRef]
- Atlihan, R.; Kasap, İ.; Özgökçe, M.S.; Polat-Akköprü, E.; Chi, H. Population growth of Dysaphis pyri (Hemiptera: Aphididae) on different pear cultivars with discussion on curve fitting in life table studies. J. Econ. Entomol. 2017, 110, 1890–1898. [Google Scholar] [CrossRef]
- Özgökçe, M.S.; Chi, H.; Atlıhan, R.; Kara, H. Demography and population projection of Myzus persicae (Sulz.) (Hemiptera: Aphididae) on five pepper (Capsicum annuum L.) cultivars. Phytoparasitica 2018, 46, 153–167. [Google Scholar] [CrossRef]
- Sousa, F.; Mendes, S.; Santos Amaya, O.; Araujo, O.; Oliveira, E.; Pereira, E. Life-history traits of Spodoptera frugiperda populations exposed to low-dose Bt maize. PLoS ONE 2016, 11, e0156608. [Google Scholar] [CrossRef] [Green Version]
- Xie, D.J.; Zhang, L.; Cheng, Y.X.; Jiang, X.F. Age-stage two-sex life table for laboratory populations of fall armyworm, Spodoptera frugiperda at different temperatures. Plant Prot. 2019, 45, 20–27. [Google Scholar]
- Zuo, T.Q.; Zhang, B.; Zhang, S.T.; Zheng, C.Y.; Wan, F.H. Combined effects of high temperature and acetamiprid on life table parameters of the F1 offspring of the treated Frankliniella occidentalis (Thysanoptera: Thripidae). Acta Entomol. Sin. 2015, 58, 767–775. [Google Scholar]
- Tsai, J.H.; Wang, J.J. Effects of host plants on biology and life table parameters of Aphis spiraecola (Homoptera: Aphididae). Environ. Entomol. 2001, 30, 44–50. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Stage (Days) | N | Maize | N | Wheat | N | Soybean | N | Tomato | N | Cotton | N | Chinese Cabbage |
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Egg | 113 | 2.49 ± 0.05 b | 101 | 2.00 ± 0 c | 101 | 2.95 ± 0.10 a | 103 | 2.00 ± 0 c | 104 | 2.00 ± 0 c | 112 | 2.0 ± 0.04 b |
| First instar | 111 | 2.07 ± 0.02 e | 97 | 2.21 ± 0.04 d | 99 | 2.84 ± 0.06 c | 98 | 3.74 ± 0.08 b | 103 | 3.65 ± 0.07 b | 57 | 4.77 ± 0.15 a |
| Second instar | 110 | 1.93 ± 0.03 e | 97 | 1.87 ± 0.04 e | 96 | 2.05 ± 0.03 d | 98 | 2.89 ± 0.09 b | 103 | 2.20 ± 0.04 c | 48 | 3.79 ± 0.15 a |
| Third instar | 109 | 1.49 ± 0.05 e | 96 | 1.57 ± 0.05 e | 95 | 2.03 ± 0.02 d | 98 | 2.60 ± 0.09 c | 103 | 2.94 ± 0.10 b | 39 | 3.46 ± 0.19 a |
| Fourth instar | 106 | 1.74 ± 0.05 d | 96 | 1.69 ± 0.05 d | 92 | 2.07 ± 0.04 c | 97 | 2.89 ± 0.08 b | 103 | 3.47 ± 0.06 a | 34 | 3.32 ± 0.17 a |
| Fifth instar | 105 | 2.08 ± 0.04 d | 95 | 2.08 ± 0.03 d | 91 | 2.29 ± 0.05 c | 96 | 3.42 ± 0.17 b | 102 | 3.81 ± 0.10 a | 24 | 3.17 ± 0.16 b |
| Sixth instar | 100 | 2.95 ± 0.10 d | 89 | 3.33 ± 0.08 b | 87 | 2.97 ± 0.08 c | 94 | 4.29 ± 0.15 b | 102 | 4.94 ± 0.13 a | 17 | 3.88 ± 0.33 b |
| Seventh instar | - | - | - | - | 37 | 3.59 ± 0.12 b | 60 | 5.77 ± 0.17 a | 34 | 5.44 ± 0.13 a | 10 | 7.00 ± 0.78 a |
| Larvae | 100 | 12.21 ± 0.11 f | 89 | 12.66 ± 0.11 e | 83 | 16.65 ± 0.16 d | 87 | 24.74 ± 0.26 b | 101 | 22.81 ± 0.27 c | 10 | 29.10 ± 0.32 a |
| Prepupa | 82 | 1.39 ± 0.06 cd | 54 | 1.30 ± 0.07 d | 76 | 1.83 ± 0.05 a | 83 | 1.54 ± 0.05b c | 99 | 1.59 ± 0.06 b | 9 | 1.56 ± 0.22 abcd |
| Pupa | 64 | 8.90 ± 0.10 b | 42 | 8.97 ± 0.13 b | 55 | 9.90 ± 0.13 a | 73 | 9.96 ± 0.10 a | 97 | 10.13 ± 0.09 a | 3 | 10.33 ± 0.49 |
| Preadult | 64 | 24.67 ± 0.16 d | 42 | 25.18 ± 0.23 d | 55 | 31.50 ± 0.24 c | 73 | 38.06 ± 0.30 a | 97 | 36.64 ± 0.31 b | 3 | 37.33 ± 0.62 |
| Adult longevity | ||||||||||||
| Female | 36 | 16.21 ± 0.11 a | 19 | 13.01 ± 1.41 a | 21 | 9.33 ± 1.84 b | 40 | 13.04 ± 1.52 a | 39 | 16.22 ± 1.89 a | 1 | 18.0 ± 0 |
| Male | 25 | 9.09 ± 1.64 a | 20 | 7.11 ± 1.53 b | 31 | 9.21 ± 0.70 a | 24 | 7.04 ± 0.64 b | 55 | 9.13 ± 0.61 a | 2 | 2.5 ± 0 |
| Stage | Maize | Wheat | Soybean | Tomato | Cotton | Chinese Cabbage |
|---|---|---|---|---|---|---|
| APOP (d) | 2.89 ± 0.21 c | 3.39 ± 0.34 bc | 4.10 ± 0 a | 4.08 ± 0.24 b | 4.72 ± 0.44 a | 4.0 |
| TPOP (d) | 26.85 ± 0.27 e | 27.94 ± 0.48 d | 34.65 ± 0.39 c | 41.33 ± 0.39 a | 39.83 ± 0.51 b | 40.0 |
| Female proportion (Nf/N) | 0.318 ± 0.044 ab | 0.188 ± 0.039 c | 0.208 ± 0.040 bc | 0.447 ± 0.049 a | 0.375 ± 0.047 a | - |
| Male proportion (Nm/N) | 0.221 ± 0.039 b | 0.198 ± 0.039 b | 0.307 ± 0.046 b | 0.233 ± 0.042 b | 0.528 ± 0.049 a | - |
| Oviposition days (d) | 7.22 ± 0.55 ab | 4.89 ± 0.64 c | 5.65 ± 0.69 bc | 4.70 ± 0.39 c | 8.11 ± 0.39 a | 12 |
| Fecundity(F) (eggs) | 1275.56 ± 155.29 a | 1180.84 ± 179.15 ab | 963.86 ± 144.89 ab | 586.17 ± 69.78 c | 803.51 ± 75.33 b | 112.90 |
| Hatching rate | 0.9496 ± 0.1438 a | 0.8938 ± 0.0408 ab | 0.9594 ± 0.0091 a | 0.9248 ± 0.0238 ab | 0.8353 ± 0.0280 b | 0 |
| Plant Species | R0 | r (Day−1) | λ (Day−1) | T (Day) |
|---|---|---|---|---|
| Maize | 406.37 ± 74.43 a | 0.2056 ± 0.0072 a | 1.2283 ± 0.0088 a | 29.21 ± 0.32 c |
| Wheat | 222.14 ± 56.53 b | 0.1827 ± 0.0101 a | 1.2004 ± 0.0120 a | 29.58 ± 0.40 c |
| Soybean | 200.41 ± 48.73 b | 0.1418 ± 0.0075 b | 1.1524 ± 0.0086 b | 37.37 ± 0.63 b |
| Tomato | 261.79 ± 42.18 ab | 0.1296 ± 0.0044 b | 1.1384 ± 0.0050 b | 42.96 ± 0.52 a |
| Cotton | 301.32 ± 47.40 ab | 0.1342 ± 0.0046 b | 1.1436 ± 0.0053 b | 42.55 ± 0.65 a |
| Chinese cabbage | 8.0 | 0.0444 | 1.0454 | 46.81 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wang, W.; He, P.; Zhang, Y.; Liu, T.; Jing, X.; Zhang, S. The Population Growth of Spodoptera frugiperda on Six Cash Crop Species and Implications for Its Occurrence and Damage Potential in China. Insects 2020, 11, 639. https://doi.org/10.3390/insects11090639
Wang W, He P, Zhang Y, Liu T, Jing X, Zhang S. The Population Growth of Spodoptera frugiperda on Six Cash Crop Species and Implications for Its Occurrence and Damage Potential in China. Insects. 2020; 11(9):639. https://doi.org/10.3390/insects11090639
Chicago/Turabian StyleWang, Wenwen, Pengyang He, Yiyang Zhang, Tongxian Liu, Xiangfeng Jing, and Shize Zhang. 2020. "The Population Growth of Spodoptera frugiperda on Six Cash Crop Species and Implications for Its Occurrence and Damage Potential in China" Insects 11, no. 9: 639. https://doi.org/10.3390/insects11090639