Divergence of Desiccation-Related Traits in Sitobion avenae from Northwestern China


Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Aphid Sampling and Colony Establishment
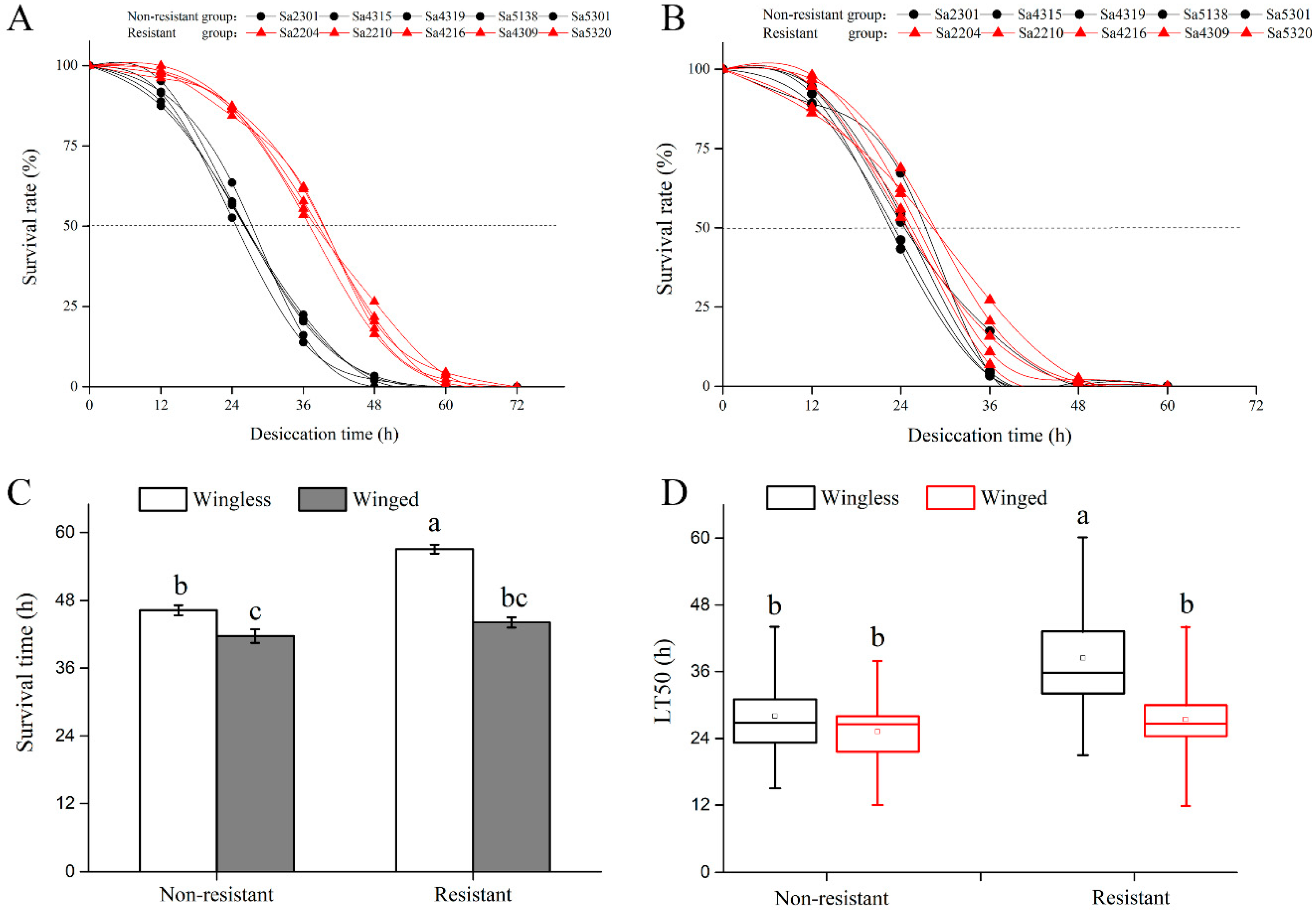
2.2. Screening for Desiccation-Resistant Clones
2.3. Bioassays for Survivorship and Water Balance Traits
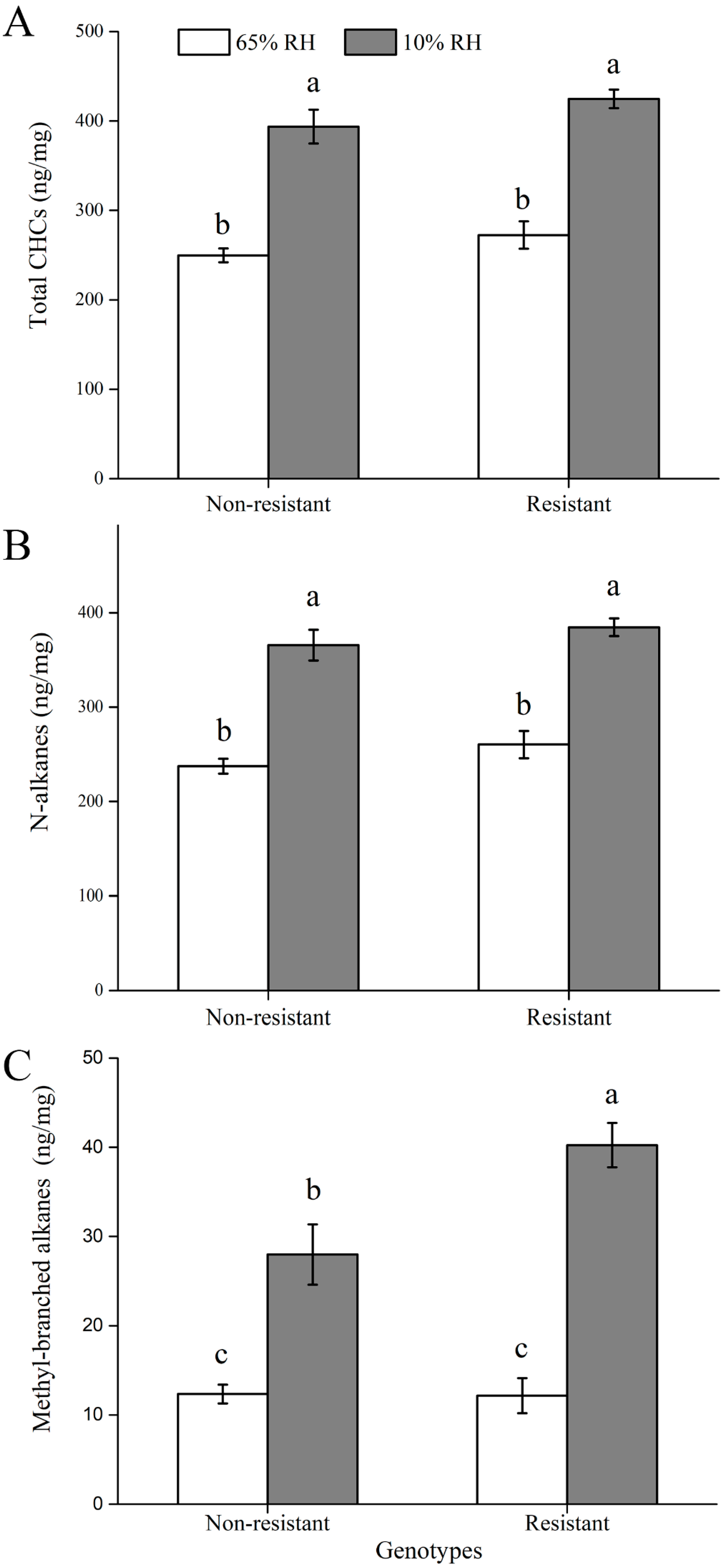
2.4. Cuticular Hydrocarbons
2.5. Statistical Analyses
3. Results
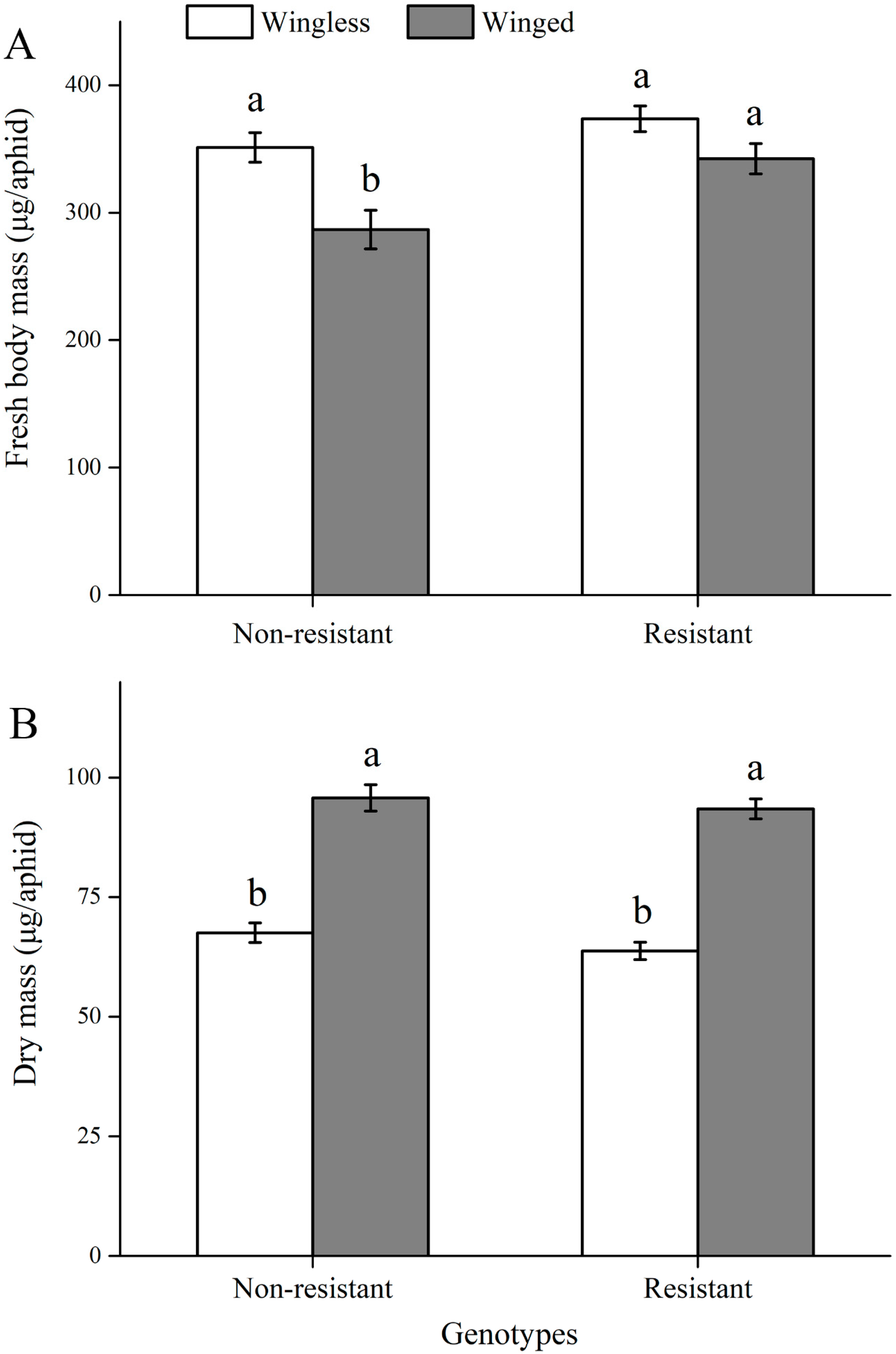
3.1. Comparisons of Fitness Traits
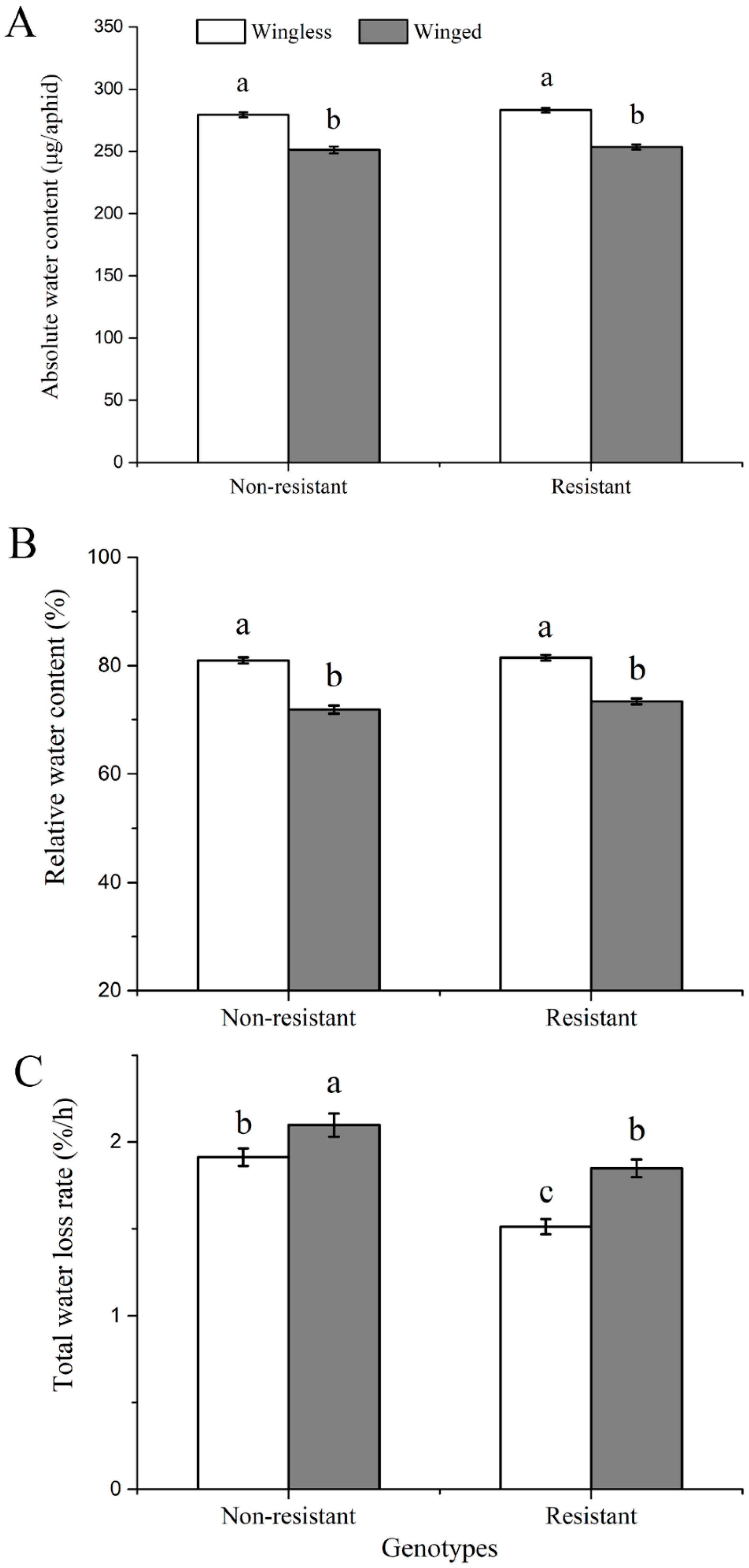
3.2. Comparisons of Water Balance Traits
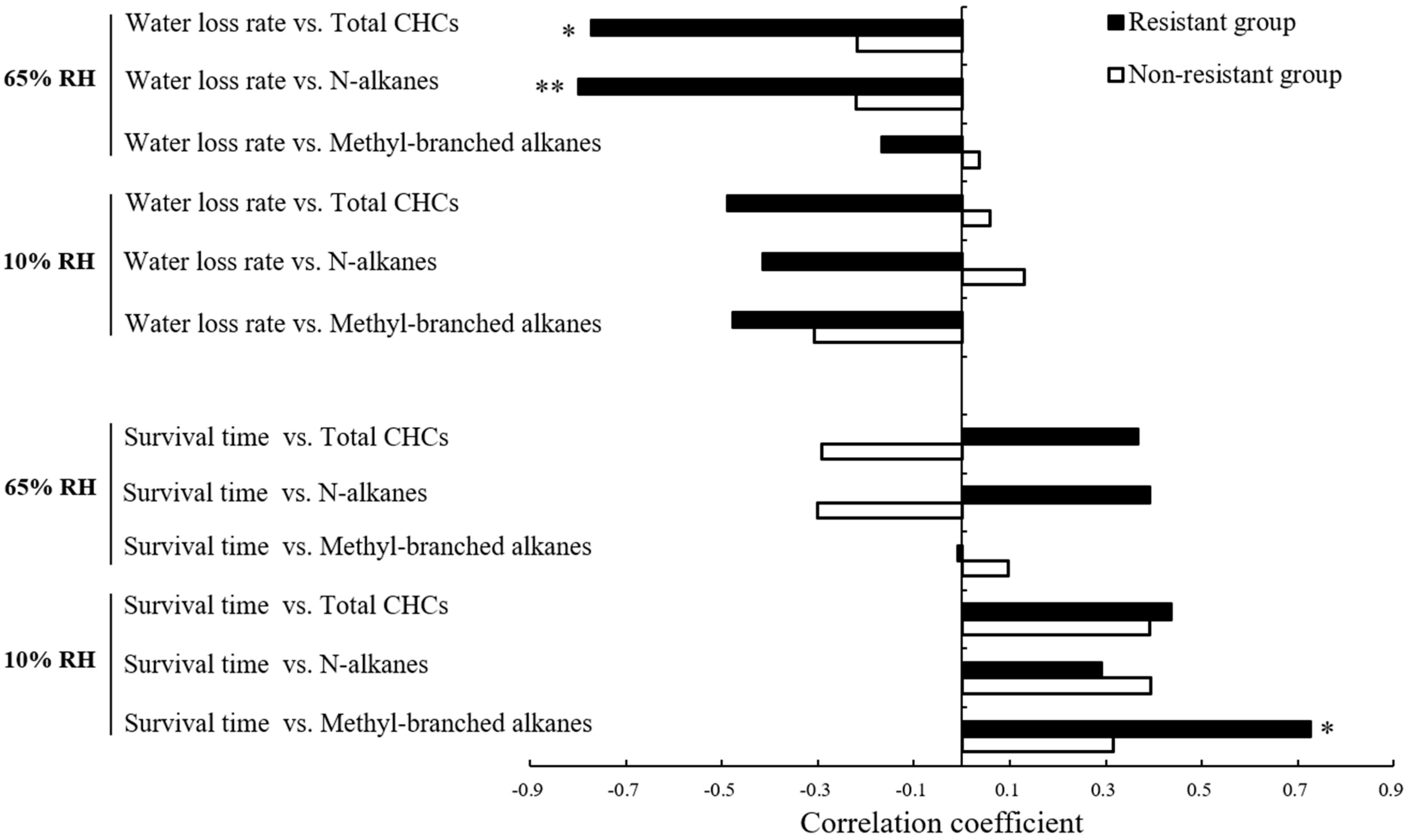
3.3. Correlations between Fitness and Water Balance Traits
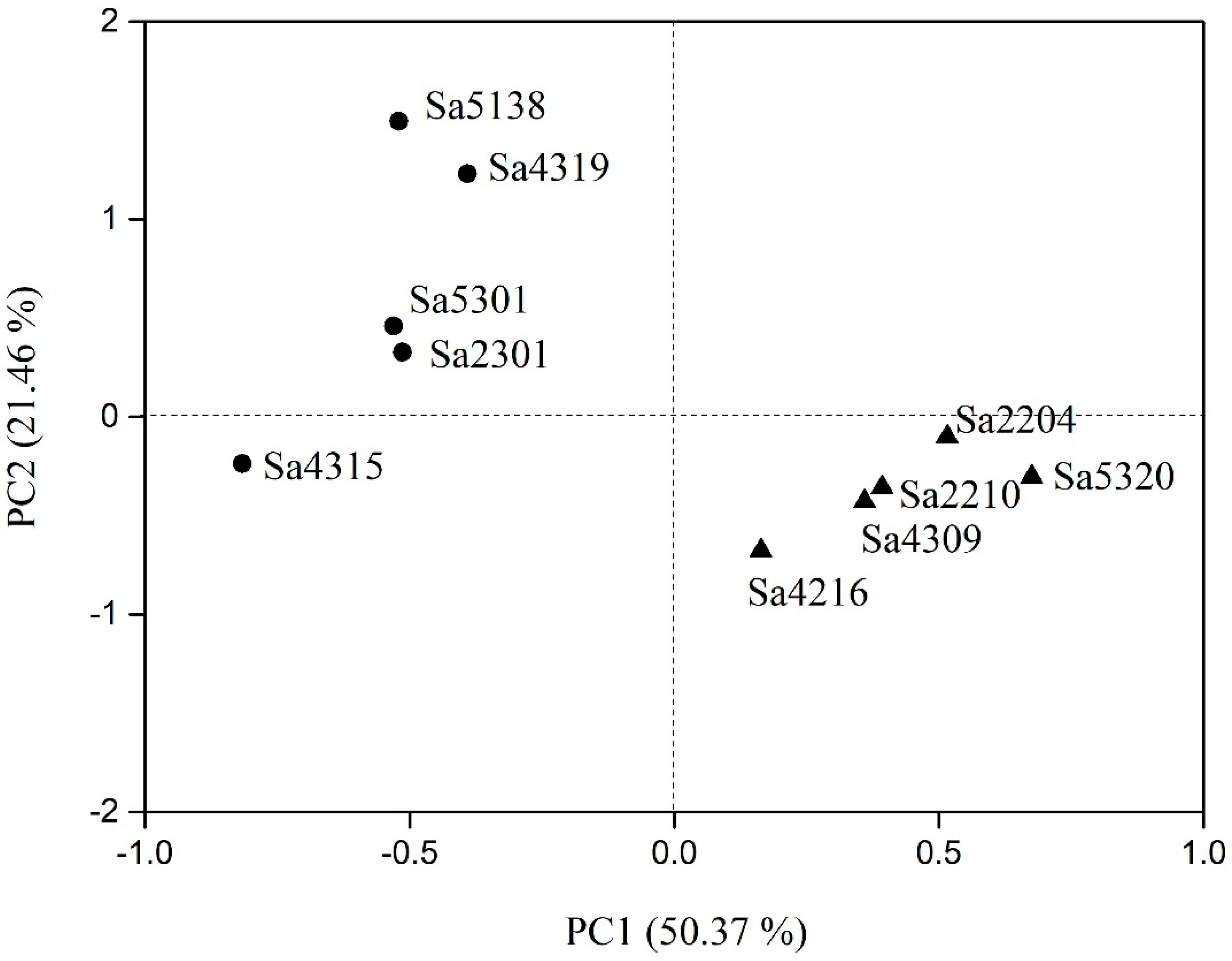
3.4. PCA Analyses of Desiccation-Related Traits
3.5. Cuticular Hydrocarbon (CHC) Contents
4. Discussion
4.1. Physiological Mechanisms of Desiccation Resistance in Aphids
4.2. Adaptive Evolution of Aphid Populations under Drought (or Desiccation Stress)
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- IPCC. Climate Change 2014: Synthesis Report; Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change (IPCC); Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; Volume 151.
- Holtzer, T.O.; Norman, J.M.; Perring, T.M.; Berry, J.S.; Heintz, J.C. Effects of microenvironment on the dynamics of spider-mite populations. Exp. Appl. Acarol. 1988, 4, 247–264. [Google Scholar] [CrossRef]
- Deng, Z.; Zhang, Q.; Pu, J.; Liu, D.; Guo, H.; Wang, Q.; Zhao, H.; Wang, H. Impact of climate warming on crop planting and production in Northwest China. Acta Ecol. Sin. 2008, 28, 3760–3768. [Google Scholar]
- Blum, A. Crop responses to drought and the interpretation of adaptation. Plant Growth Regul. 1996, 20, 135–148. [Google Scholar] [CrossRef]
- Basu, S.; Ramegowda, V.; Kumar, A.; Pereira, A. Plant adaptation to drought stress. F1000research 2016, 5, 1554–1564. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, A.G.; Chippindale, A.K.; Rose, M.R. Physiological mechanisms of evolved desiccation resistance in Drosophila melanogaster. J. Exp. Biol. 1997, 200, 1821–1832. [Google Scholar]
- Chown, S.L.; Klok, C.J. Water-balance characteristics respond to changes in body size in Subantarctic weevils. Physiol. Biochem. Zool. 2003, 76, 634–643. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.A. Acclimation for desiccation resistance in Drosophila: Species and population comparisons. J. Insect Physiol. 1991, 37, 757–762. [Google Scholar] [CrossRef]
- Bazinet, A.L.; Marshall, K.E.; Macmillan, H.A.; Williams, C.M.; Sinclair, B.J. Rapid changes in desiccation resistance in Drosophila melanogaster are facilitated by changes in cuticular permeability. J. Insect Physiol. 2010, 56, 2006–2012. [Google Scholar] [CrossRef]
- Parkash, R.; Ramniwas, S.; Kajla, B.; Aggarwal, D.D. Divergence of desiccation-related traits in two Drosophila species of the takahashii subgroup from the Western Himalayas. J. Exp. Biol. 2012, 215, 2181–2191. [Google Scholar] [CrossRef] [Green Version]
- Kalra, B.; Parkash, R. Sex-specific divergence for body size and desiccation-related traits in Drosophila hydei from the Western Himalayas. Comp. Biochem. Phys. A 2014, 177, 1–10. [Google Scholar] [CrossRef]
- Ferveur, J.F.; Jerme, C.; Rihani, K.; Cobb, M.; Everaerts, C. Desiccation resistance: Effect of cuticular hydrocarbons and water content in Drosophila melanogaster adults. PeerJ 2018, 6, e4318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Douglas, A.E. Phloem-sap feeding by animals: Problems and solutions. J. Exp. Bot. 2006, 57, 747–754. [Google Scholar] [CrossRef] [PubMed]
- Pons, X.; Tatchell, G.M. Drought stress and cereal aphid performance. Ann. Appl. Biol. 1995, 126, 19–31. [Google Scholar] [CrossRef]
- Mcvean, R.I.K.; Dixon, A.F.G. The effect of plant drought-stress on populations of the pea aphid Acyrthosiphon pisum. Ecol. Entomol. 2001, 26, 440–443. [Google Scholar] [CrossRef]
- Bestete, L.R.; Torres, J.B.; Silva, R.B.B.; Silva-Torres, C.S.A. Water stress and kaolin spray affect herbivorous insects’ success on cotton. Arthropod-Plant Int. 2016, 10, 445–453. [Google Scholar] [CrossRef]
- Archer, T.L.; Bynum, E.D.; Onken, A.B.; Wendt, C.W. Influence of water and nitrogen fertilizer on biology of the Russian wheat aphid (Homoptra: Aphdidae) on wheat. Crop Prot. 1995, 14, 165–169. [Google Scholar] [CrossRef]
- Burgess, A.J.; Warrington, S.; Allen-Williams, L. Cabbage aphid (Brevicoryne brassicae L.) ‘performance’ on oilseed rape (Brassica napus L.) experiencing water deficiency: Roles of temperature and food quality. Acta Hortic. 1996, 407, 499–506. [Google Scholar] [CrossRef]
- Dorschner, K.W. Insect-plant interactions: Greenbug (Homoptera: Aphididae) disrupt acclimation of winter wheat to drought stress. Environ. Entomol. 1986, 15, 118–121. [Google Scholar] [CrossRef]
- Li, H.; Payne, W.A.; Michels, G.J.; Rush, C.M. Reducing plant abiotic and biotic stress: Drought and attacks of greenbugs, corn leaf aphids and virus disease in dryland sorghum. Environ. Exp. Bot. 2008, 63, 305–316. [Google Scholar] [CrossRef]
- Tariq, M.; Wright, D.J.; Rossiter, J.T.; Staley, J.T. Aphids in a changing world: Testing the plant stress, plant vigour and pulsed stress hypotheses. Agric. Forest Entomol. 2012, 14, 177–185. [Google Scholar] [CrossRef]
- Dai, P.; Liu, D.G.; Shi, X.Q. Impacts of water deficiency on life history of Sitobion avenae clones from semiarid and moist areas. J. Econ. Entomol. 2015, 108, 2250–2258. [Google Scholar] [CrossRef] [PubMed]
- He, Y.; Liu, D.G.; Dai, P.; Wang, D.; Shi, X.Q. Genetic differentiation and structure of Sitobion avenae (Hemiptera: Aphididae) populations from moist, semiarid and arid areas in Northwestern China. J. Econ. Entomol. 2018, 111, 603–611. [Google Scholar] [CrossRef] [PubMed]
- Gibbs, A.G.; Matzkin, L.M. Evolution of water balance in the genus Drosophila. J. Exp. Biol. 2001, 204, 2331–2338. [Google Scholar] [PubMed]
- Addo-Bediako, A.; Chown, S.L.; Gaston, K.J. Revisiting water loss in insects: A large scale view. J. Insect Physiol. 2001, 47, 1377–1388. [Google Scholar] [CrossRef]
- Gibbs, A.G.; Rajpurohit, S. Insect Hydrocarbons: Cuticular Lipids and Water Balance; Cambridge University Press: Cambridge, UK, 2010; Volume 6, pp. 100–120. [Google Scholar]
- Chen, N.; Fan, Y.L.; Bai, Y.; Li, X.D.; Liu, T.X. Cytochrome p450 gene, CYP4G51, modulates hydrocarbon production in the pea aphid, Acyrthosiphon pisum. Insect Biochem. Mol. Biol. 2016, 76, 84–94. [Google Scholar] [CrossRef]
- Liu, S.H.; Stoltz, R.L.; Ni, X. Damage to wheat by Macrosiphum avenae (F.) (Homoptera: Aphididae) in Northwest China. J. Econ. Entomol. 1986, 79, 1688–1691. [Google Scholar]
- Huang, X.L.; Liu, D.G.; Gao, S.X.; Chen, H. Differential performance of Sitobion avenae populations from both sides of the Qinling Mountains under common garden conditions. Environ. Entomol. 2013, 42, 1174–1183. [Google Scholar] [CrossRef]
- Huang, X.L.; Liu, D.G.; Wang, D.; Shi, X.Q.; Simon, J.C. Molecular and quantitative genetic differentiation in Sitobion avenae populations from both sides of the Qinling Mountains. PLoS ONE 2015, 10, e0122343. [Google Scholar] [CrossRef] [Green Version]
- Dai, P.; Shi, X.Q.; Liu, D.G.; Ge, Z.H.; Wang, D.; Dai, X.J.; Yi, Z.H.; Meng, X.X. Life-history trait plasticity and its relationships with plant adaptation and insect fitness: A case study on the aphid Sitobion avenae. Sci. Rep. 2016, 6, 29974. [Google Scholar] [CrossRef]
- Liu, D.G.; Dai, P.; Li, S.R.; Ahmed, S.; Shang, Z.M.; Shi, X.Q. Life-history responses of insects to water deficit stress a case study with the aphid Sitobion avenae. BMC Ecol. 2018, 18, 17–31. [Google Scholar] [CrossRef]
- Bai, J.; Yan, J.P.; Su, K.H. Differential analysis of abrupt climate change between southern and northern Qinling mountains in the past 50 years. J. Shaanxi Norm. Univ. 2010, 38, 98–105. [Google Scholar]
- Zhai, L.; Feng, Q. Spatial and temporal pattern of precipitation and drought in Gansu Province, Northwest China. Nat. Hazards 2009, 49, 1–24. [Google Scholar] [CrossRef]
- Gao, S.; Liu, D.G. Differential performance of Sitobion avenae (Hemiptera: Aphididae) clones from wheat and barley with implications for its management through alternative cultural practices. J. Econ. Entomol. 2013, 106, 1294–1301. [Google Scholar] [CrossRef] [PubMed]
- Simon, J.C.; Baumann, S.; Sunnucks, P.; Hebert, P.D.N.; Pierre, J.S.; Gallic, J.F.L.; Dedryver, C.A. Reproductive mode and population genetic structure of the cereal aphid Sitobion avenae studied using phenotypic and microsatellite markers. Mol. Ecol. 1999, 8, 531–545. [Google Scholar] [CrossRef] [PubMed]
- Wilson, A.C.C.; Massonnet, B.; Simon, J.C.; Prunier-Leterme, N.; Dolatti, L.; Llewellyn, K.S.; Figueroa, C.C.; Ramirez, C.C.; Blackman, R.L.; Estoup, A.; et al. Cross-species amplification of microsatellite loci in aphids: Assessment and application. Mol. Ecol. 2004, 4, 104–109. [Google Scholar] [CrossRef]
- Dai, X.; Gao, S.; Liu, D. Genetic basis and selection for life-history trait plasticity on alternative host plants for the cereal aphid Sitobion avenae. PLoS ONE 2014, 9, e106179. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wharton, G.W. Water balance of insects. In Comprehensive Insect Physiology, Biochemistry, and Pharmacology; Pergamon Press: Oxford, UK, 1985; Volume 4, pp. 565–603. [Google Scholar]
- Young, H.P.; Schal, C. Cuticular hydrocarbon synthesis in relation to feeding and developmental stage in nymphs of Blattella germanica (Dictyoptera: Blattellidae). Ann. Entomol. Soc. Am. 1997, 90, 655–663. [Google Scholar] [CrossRef] [Green Version]
- Hadley, N.F. Ventilatory patterns and respiratory transpiration in adult terrestrial insects. Physiol. Zool. 1994, 67, 175–189. [Google Scholar] [CrossRef]
- Mackay, P.A.; Downer, R.G.H. Water content, weight change, and activity of apterous and alate virginoparous, Acyrthosiphon pisum (Harris) (Homoptera: Aphididae). Can. J. Zool. 1979, 57, 363–367. [Google Scholar] [CrossRef]
- Daniels, M.; Bale, J.S.; Newbury, H.J. A sublethal dose of thiamethoxam causes a reduction in xylem feeding by the bird cherry-oat aphid (Rhopalosiphum padi), which is associated with dehydration and reduced performance. J. Insect Physiol. 2009, 55, 758–765. [Google Scholar] [CrossRef]
- Nalam, V.J.; Keeretaweep, J.; Sarowar, S. Root-derived oxylipins promote green peach aphid performance on Arabidopsis foliage. Plant. Cell 2012, 24, 1643–1653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.; Neoh, K.B.; Appel, A.G.; Lee, C.Y. Subterranean termite open-air foraging and tolerance to desiccation: Comparative water relation of two sympatric Macrotermes spp. (Blattodea: Termitidae). Comp. Biochem. Phys. A 2012, 161, 201–207. [Google Scholar] [CrossRef] [PubMed]
- Gray, E.M.; Bradley, T.J. Physiology of desiccation resistance in Anopheles gambiae and Anopheles arabiensis. Am. J. Trop. Med. Hyg. 2005, 73, 553–559. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parkash, R.; Ranga, P. Sex-specific divergence for adaptations to dehydration stress in Drosophila kikkawai. J. Exp. Biol. 2013, 216, 3301–3313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zukowski, J.; Su, N.Y. Cuticular permeability, percent body water loss, and relative humidity equilibria comparisons of four termite species. J. Insect Sci. 2019, 19, 12–18. [Google Scholar] [CrossRef]
- Rolandi, C.; Mónica, S.I.; Schilman, P.E. Metabolism and water loss rate of the haematophagous insect Rhodnius prolixus: Effect of starvation and temperature. J. Exp. Biol. 2014, 217, 4414–4422. [Google Scholar] [CrossRef] [Green Version]
- How, Y.F.; Lee, C.Y. Effects of temperature and humidity on the survival and water loss of Cimex hemipterus (Hemiptera: Cimicidae). J. Med. Entomol. 2010, 47, 987–995. [Google Scholar] [CrossRef]
- Wang, S.Y.; Li, B.L.; Zhang, D.Y. NlCYP4G76 and NlCYP4G115 modulate susceptibility to desiccation and insecticide penetration through affecting cuticular hydrocarbon biosynthesis in Nilaparvata lugens (Hemiptera: Delphacidae). Front. Physiol. 2019, 10, 913–924. [Google Scholar] [CrossRef] [Green Version]
- Harrison, J.F. Insect acid-base physiology. Ann. Rev. Entomol. 2001, 46, 221–250. [Google Scholar] [CrossRef]
- Folk, D.G.; Bradley, T.J. Evolved patterns and rates of water loss and ion regulation in laboratory-selected populations of Drosophila melanogaster. J. Exp. Biol. 2003, 206, 2779–2786. [Google Scholar] [CrossRef] [Green Version]
- Bradley, T.J. The excretory system: Structure and physiology. In Comprehensive Insect Physiology, Biochemistry and Pharmacology; Pergamon Press: Elmsford, NY, USA, 1985; Volume 4, pp. 421–465. [Google Scholar]
- Marais, E.; Klok, C.J.; Terblanche, J.S.; Chown, S.L. Insect gas exchange patterns: A phylogenetic perspective. J. Exp. Biol. 2005, 208, 4495–4507. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chown, S.L.; Gibbs, A.G.; Hetz, S.K.; Klok, C.J.; Lighton, J.R.B.; Marais, E. Discontinuous gas exchange in insects: A clarification of hypotheses and approaches. Physiol. Biochem. Zool. 2006, 79, 333–343. [Google Scholar] [CrossRef] [Green Version]
- Terblanche, J.S.; Clusella-Trullas, S.; Chown, S.L. Phenotypic plasticity of gas exchange pattern and water loss in Scarabaeus spretus (Coleoptera: Scarabaeidae): Deconstructing the basis for metabolic rate variation. J. Exp. Biol. 2010, 213, 2940–2949. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Howard, R.W.; Jackson, L.L.; Banse, H.; Blows, M.W. Cuticular hydrocarbons of Drosophila birchii and D. serrata: Identification and role in mate choice in D. serrata. J. Chem. Ecol. 2003, 29, 961–976. [Google Scholar] [CrossRef] [PubMed]
- Johnson, R.A.; Gibbs, A.G. Effects of mating stage on water balance, metabolism and cuticular hydrocarbons of the desert harvester ant, Pogonomyrmex barbatus. J. Insect Physiol. 2004, 50, 943–953. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.Z.; Dai, Z.Y.; Shen, W.H.; Zhao, T.F. A comparasive study of some biological traits between alatae and apterous of bird cherry-oat aphid. J. Jiangsu Agric. Coll. 1995, 16, 31–34. [Google Scholar]
- Li, Y.L.; Zhao, Q.; Duan, X.L.; Song, C.M.; Chen, M.H. Transcription of four Rhopalosiphum padi (L.) heat shock protein genes and their responses to heat stress and insecticide exposure. Comp. Biochem. Phys. A 2017, 205, 48–57. [Google Scholar] [CrossRef]
- Brisson, J.A.; Davis, G.K.; Stern, D.L. Common genome-wide patterns of transcript accumulation underlying the wing polyphenism and polymorphism in the pea aphid (Acyrthosiphon pisum). Evol. Dev. 2007, 9, 338–346. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.W.; Liu, X.X.; Xu, X.L.; Li, Z.; Li, Y.S.; Song, D.Y.; Yu, T.; Zhu, F.; Zhang, Q.W.; Zhou, X.G. Gene expression profiling in winged and wingless cotton aphids, Aphis gossypii (Hemiptera: Aphididae). Int. J. Biol. Sci. 2014, 10, 257–267. [Google Scholar] [CrossRef] [Green Version]
- Kobayashi, M.; Ishikawa, H. Breakdown of indirect flight muscles of alate aphids (Acyrthosiphon pisum) in relation to their flight, feeding and reproductive behavior. J. Insect Physiol. 1993, 39, 549–554. [Google Scholar] [CrossRef]
- Shi, S.L.; Liu, X.X.; Zhang, Q.W.; Zhao, Z.W. Morph-specific differences in metabolism related to flight in the wing-dimorphic Aphis gossypii. Insect Sci. 2010, 6, 527–534. [Google Scholar] [CrossRef]
- Zachariassen, K.E.; Andersen, J.; Maloiy, G.M.O.; Kamau, J.M.Z. Transpiratory water loss and metabolism of beetles from arid areas in East Africa. Comp. Biochem. Phys. B 1987, 86, 403–408. [Google Scholar] [CrossRef]
- Gibbs, A.G.; Fukuzato, F.; Matzkin, L.M. Evolution of water conservation mechanisms in Drosophila. J. Exp. Biol. 2003, 206, 1183–1192. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- King, K.J.; Sinclair, B.J. Water loss in tree weta (Hemideina): Adaptation to the montane environment and a test of the melanisation-desiccation resistance hypothesis. J. Exp. Biol. 2015, 218, 1995–2004. [Google Scholar] [CrossRef] [Green Version]
- Parkash, R.; Lambhod, C.; Singh, D. Thermal developmental plasticity affects body size and water conservation of Drosophila nepalensis from the Western Himalayas. Bull. Entomol. Res. 2014, 104, 504–516. [Google Scholar] [CrossRef]
- Kellermann, V.; Hoffmann, A.A.; Overgaard, J.; Loeschcke, V.; Sgro, C.M. Plasticity for desiccation tolerance across Drosophila species is affected by phylogeny and climate in complex ways. Proc. Biol. Sci. 2018, 285, 48. [Google Scholar] [CrossRef] [Green Version]
- Davis, J.S.; Moyle, C. Constitutive and plastic gene expression variation associated with desiccation resistance differences in the Drosophila americana species group. Genes 2020, 11, 146. [Google Scholar] [CrossRef] [Green Version]
- Philip, B.N.; Yi, S.X.; Elnitsky, M.A.; Lee, R.E. Aquaporins play a role in desiccation and freeze tolerance in larvae of the goldenrod gall fly, Eurosta solidaginis. J. Exp. Biol. 2008, 211, 1114–1119. [Google Scholar] [CrossRef] [Green Version]
- Urbanski, J.M.; Benoit, J.B.; Michaud, M.R.; Denlinger, D.L.; Armbruster, P. The molecular physiology of increased egg desiccation resistance during diapause in the invasive mosquito, Aedes albopictus. Proc. Biol. Sci. 2010, 277, 2683–2692. [Google Scholar] [CrossRef] [Green Version]
- Benoit, J.B.; Lopez-Martinez, G.; Phillips, Z.P.; Patrick, K.R.; Denlinger, D.L. Heat shock proteins contribute to mosquito dehydration tolerance. J. Insect. Physiol. 2010, 56, 151–156. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Variance Source | df | F | p | % Total |
|---|---|---|---|---|---|
| Survival time | Genotype | 9 | 5.48 | <0.001 | 7.70 |
| Wing | 1 | 61.64 | <0.001 | 86.57 | |
| Genotype×wing | 9 | 3.08 | 0.002 | 4.33 | |
| Error | 242 | 1.40 | |||
| LT 50 | Genotype | 9 | 6.71 | <0.001 | 15.74 |
| Wing | 1 | 28.88 | <0.001 | 67.71 | |
| Genotype×wing | 9 | 6.06 | <0.001 | 14.21 | |
| Error | 242 | 2.34 | |||
| Absolute water content | Genotype | 9 | 6.44 | <0.001 | 5.66 |
| Wing | 1 | 105.21 | <0.001 | 92.46 | |
| Genotype×wing | 9 | 1.14 | 0.333 | 1.00 | |
| Error | 242 | 0.88 | |||
| Relative water content | Genotype | 9 | 6.37 | <0.001 | 4.82 |
| Wing | 1 | 123.85 | <0.001 | 93.58 | |
| Genotype×wing | 9 | 1.13 | 0.344 | 0.85 | |
| Error | 242 | 0.76 | |||
| Water loss rate | Genotype | 9 | 7.19 | <0.001 | 24.70 |
| Wing | 1 | 19.93 | <0.001 | 68.42 | |
| Genotype×wing | 9 | 1.00 | 0.437 | 3.45 | |
| Error | 242 | 3.43 |
| Wingless | Winged | |||||
|---|---|---|---|---|---|---|
| Absolute Water Content | Relative Water Content | Water Loss Rate | Absolute Water Content | Relative Water Content | Water Loss Rate | |
| Nonresistant genotypes | ||||||
| Survival time | 0.398 ** | −0.074 | −0.349 ** | 0.173 | −0.114 | −0.311 |
| LT 50 | 0.542 *** | −0.212 | −0.416 *** | 0.533 *** | −0.345 * | −0.342 * |
| Resistant genotypes | ||||||
| Survival time | 0.553 *** | −0.043 | −0.274 ** | 0.370 ** | −0.136 | −0.293 * |
| LT 50 | 0.425 *** | −0.068 | −0.334 ** | 0.481 *** | −0.232 | −0.415 * |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, Y.; Liu, D.; Liu, X.; Wang, B.; Shi, X. Divergence of Desiccation-Related Traits in Sitobion avenae from Northwestern China. Insects 2020, 11, 626. https://doi.org/10.3390/insects11090626
Yang Y, Liu D, Liu X, Wang B, Shi X. Divergence of Desiccation-Related Traits in Sitobion avenae from Northwestern China. Insects. 2020; 11(9):626. https://doi.org/10.3390/insects11090626
Chicago/Turabian StyleYang, Yujing, Deguang Liu, Xiaoming Liu, Biyao Wang, and Xiaoqin Shi. 2020. "Divergence of Desiccation-Related Traits in Sitobion avenae from Northwestern China" Insects 11, no. 9: 626. https://doi.org/10.3390/insects11090626